


Vector Competence of Culex quinquefasciatus from Brazil for West Nile Virus

 , , , , , and
, , , , , and 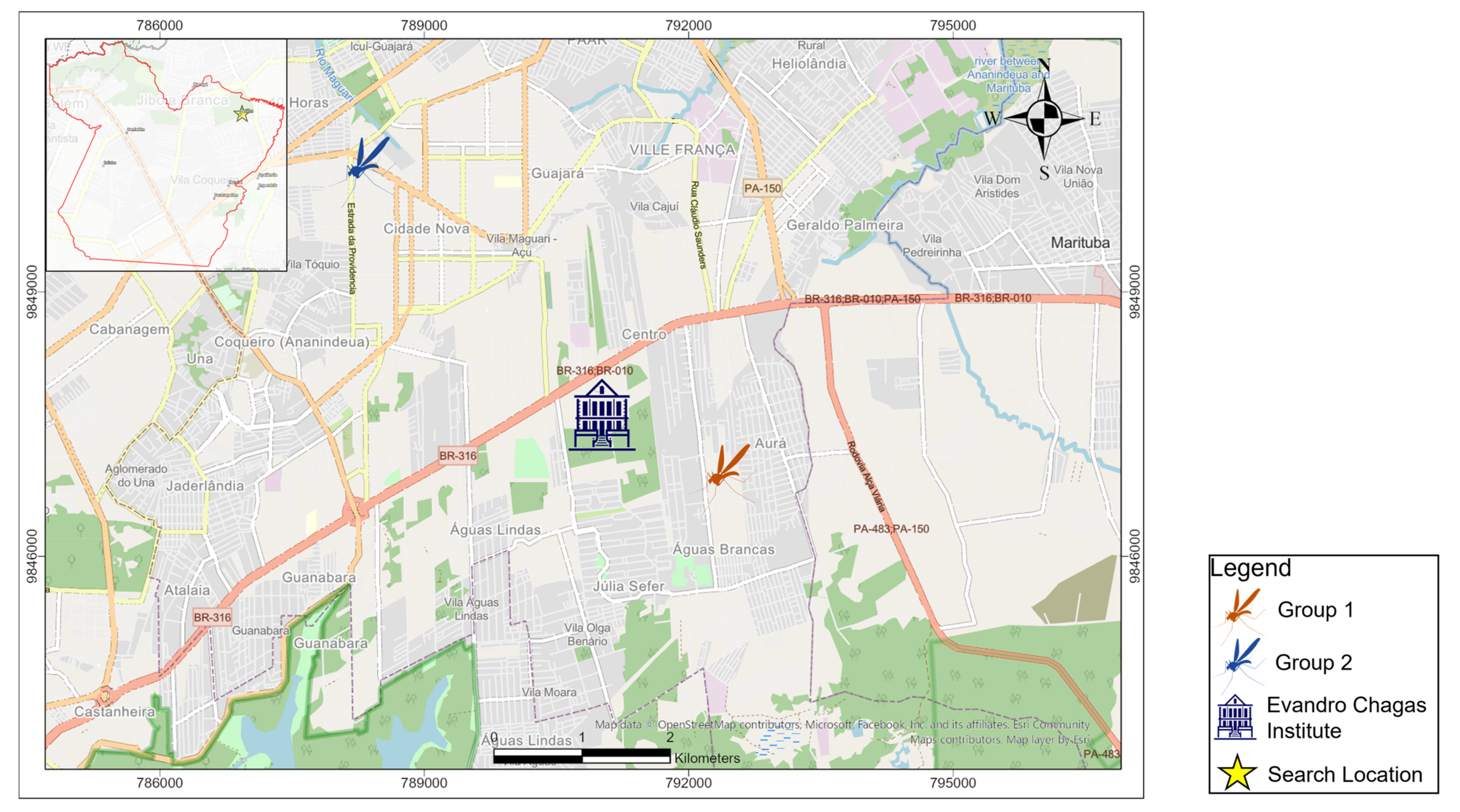{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Infection
2.2. Viral Strain
2.3. Viral Stock Preparation
2.4. Mosquitoes Infection
2.5. Mosquito Segmentation
2.6. Virus Isolation
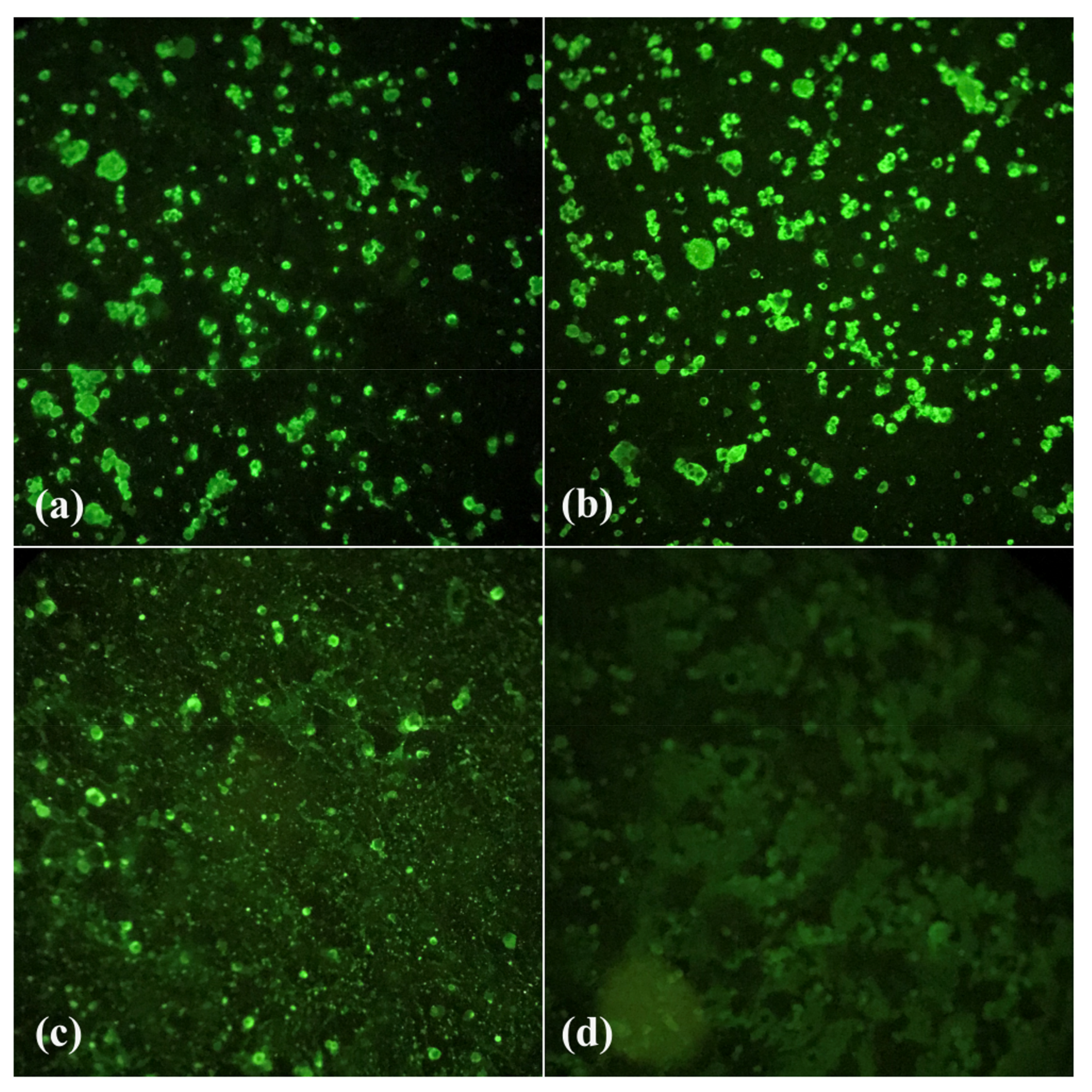
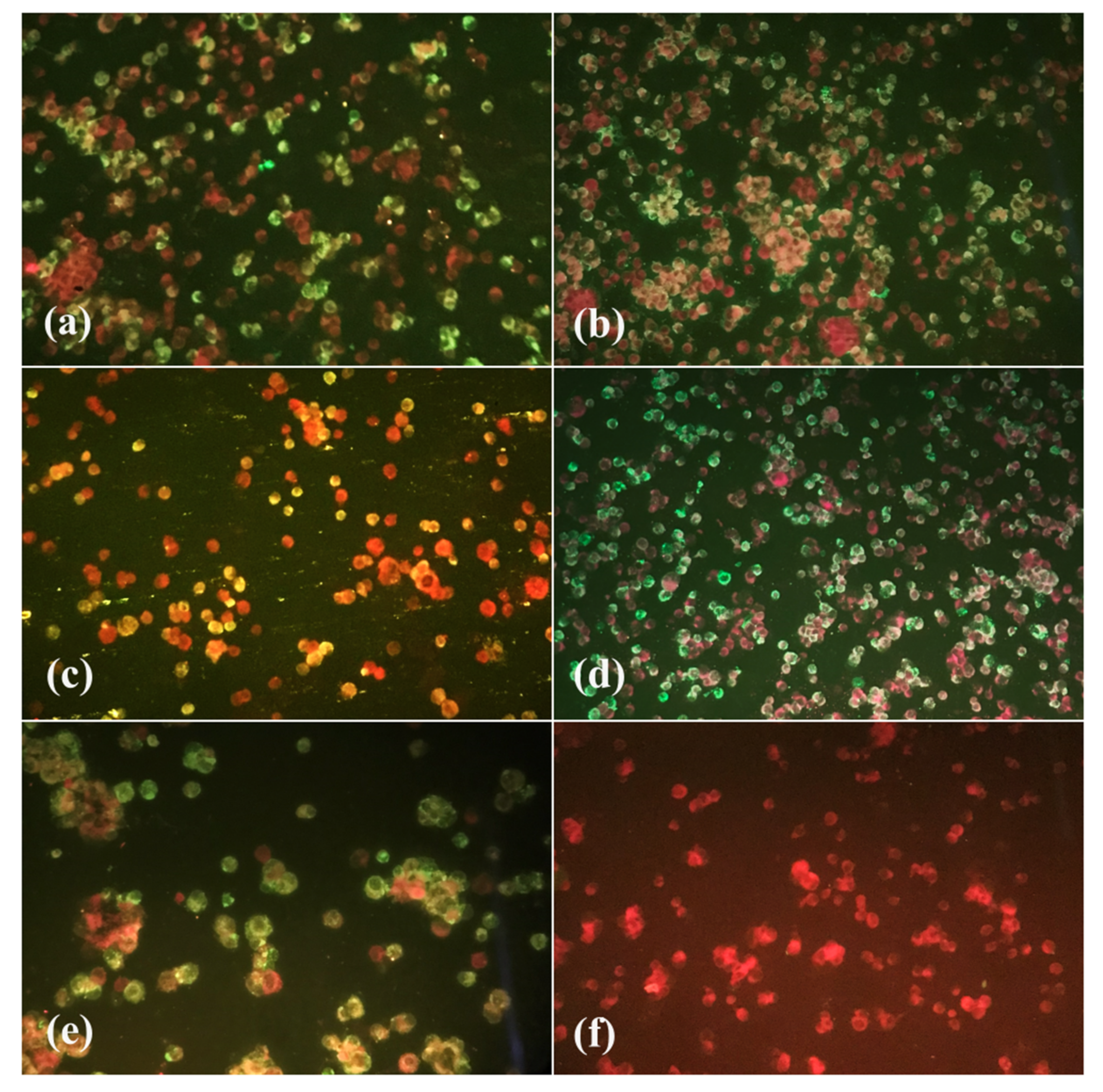
2.7. Indirect Immunofluorescence Test (IF)
2.8. Viral Titration
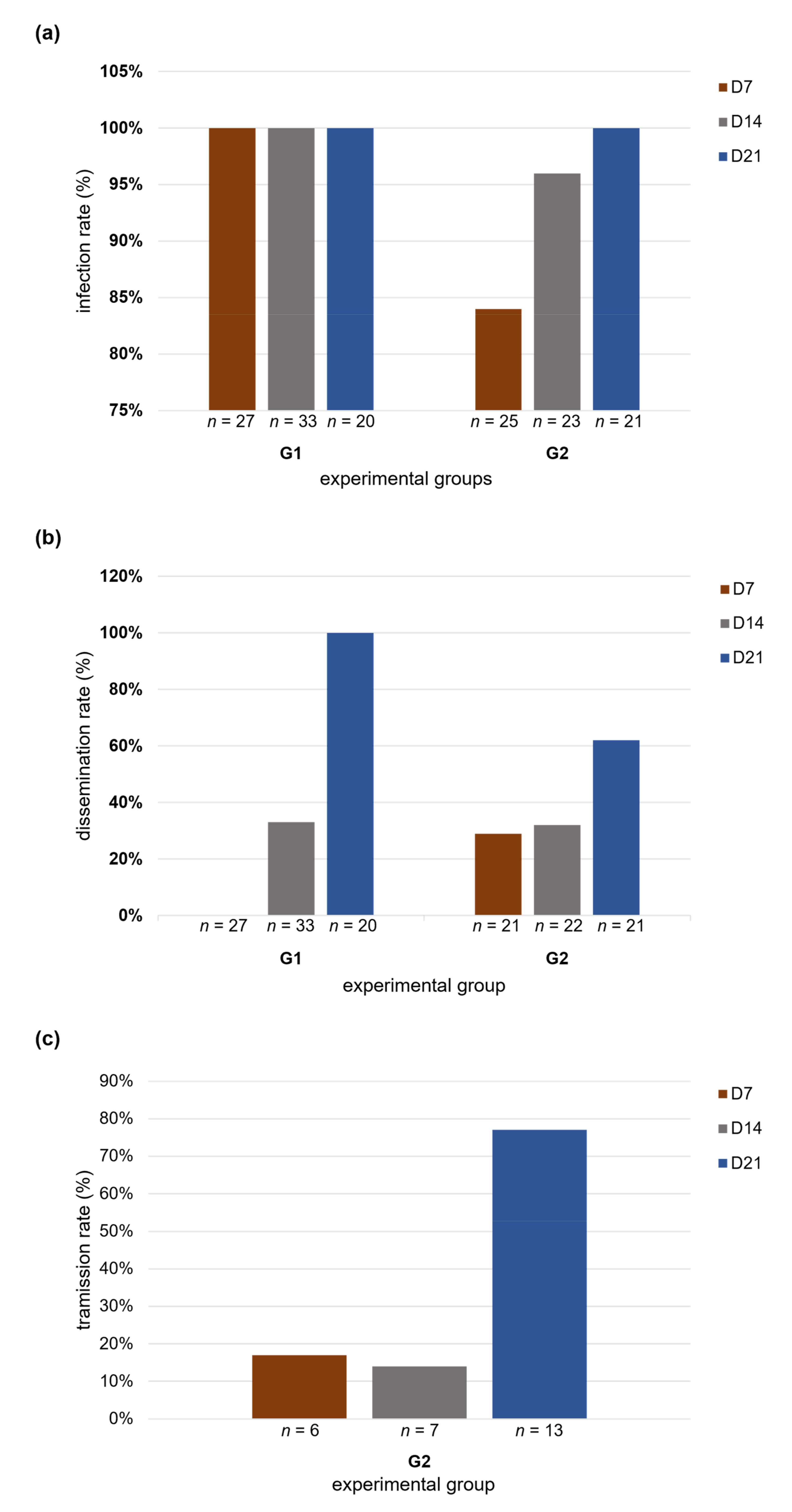
2.9. Infection, Dissemination and Transmission Rates
2.10. Statistical Analysis
3. Results
3.1. Infection, Dissemination and Transmission Rates
3.2. Viral Titration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopes, N.; Nozawa, C.; Elisa, R.; Linhares, C. Características Gerais e Epidemiologia Dos Arbovírus Emergentes No Brasil General Features and Epidemiology of Emerging Arboviruses in Brazil Características Generales y Epidemiología de Los Arbovirus Emergentes En Brasil. Rev. Pan-Amaz. Saude 2014, 5, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Neves, D.P.; De Filippis, T.; Dias-Lima, A.; Oda, W.Y. Artrópodes. In Parasitologia Básica; Atheneu: Rio de Janeiro, Brazil, 2019; p. 2144. ISBN 978-85-388-0934-0. [Google Scholar]
- Donalisio, M.R.; Freitas, A.R.R.; Von Zuben, A.P.B. Arboviruses Emerging in Brazil: Challenges for Clinic and Implications for Public Health. Rev. Saude Publica 2017, 51, 30. [Google Scholar] [CrossRef] [PubMed]
- Duguma, D.; Rueda, L.M.; Debboun, M. Mosquito-Borne Diseases. In Mosquitoes, Communities, and Public Health in Texas; Debboun, M., Nava, M.R., Rueda, L.M., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 319–337. ISBN 978-0-12-814545-6. [Google Scholar]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A Neurotropic Virus Isolated from the Blood of a Native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar] [CrossRef]
- Martins, L.C.; Da Silva, E.V.P.; Casseb, L.M.N.; Da Silva, S.P.; Cruz, A.C.R.; Pantoja, J.A.d.S.; Medeiros, D.B.d.A.; Martins, A.J.; Da Cruz, E.D.R.M.; De Araújo, M.T.F.; et al. First Isolation of West Nile Virus in Brazil. Mem. Inst. Oswaldo Cruz 2019, 114, 180332. [Google Scholar] [CrossRef]
- Ciota, A.T.; Kramer, L.D. Vector-Virus Interactions and Transmission Dynamics of West Nile Virus. Viruses 2013, 5, 3021–3047. [Google Scholar] [CrossRef]
- Ministry of Health; Secretariat of Health Surveillance. Chapter 6—West Nile Fever. In Guia de Vigilância em Saúde: Volume Único; Oliveira, W.K., Rohlfs, D.B., Macário, E.M., Pereira, G.F.M., Croda, J.H., Brito, S.M.F., Eds.; Ministério da Saúde: Brasília, Brazil, 2019; 1, pp. 389–400. ISBN 978-85-334-2706-8. [Google Scholar]
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile Virus Responsible for an Outbreak of Encephalitis in the Northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [Green Version]
- Colpitts, T.M.; Conway, M.J.; Montgomery, R.R.; Fikrig, E. West Nile Virus: Biology, Transmission, and Human Infection. Clin. Microbiol. Rev. 2012, 25, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Laboratory-Acquired West Nile Virus Infections—United States. 2002. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm5150a2.htm (accessed on 18 August 2021).
- Iwamoto, M.; Jernigan, D.B.; Guasch, A.; Trepka, M.J.; Blackmore, C.G.; Hellinger, W.C.; Pham, S.M.; Zaki, S.; Lanciotti, R.S.; Lance-Parker, S.E.; et al. Transmission of West Nile Virus from an Organ Donor to Four Transplant Recipients. N. Engl. J. Med. 2009, 348, 2196–2203. [Google Scholar] [CrossRef]
- Winston, D.J.; Vikram, H.R.; Rabe, I.B.; Dhillon, G.; Mulligan, D.; Hong, J.C.; Busuttil, R.W.; Nowicki, M.J.; Mone, T.; Civen, R.; et al. Donor-Derived West Nile Virus Infection in Solid Organ Transplant Recipients: Report of Four Additional Cases and Review of Clinical, Diagnostic, and Therapeutic Features HHS Public Access. Transplantation 2014, 97, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Pealer, L.N.; Marfin, A.A.; Petersen, L.R.; Lanciotti, R.S.; Page, P.L.; Stramer, S.L.; Stobierski, M.G.; Signs, K.; Newman, B.; Kapoor, H.; et al. Transmission of West Nile Virus through Blood Transfusion in the United States in 2002. N. Engl. J. Med. 2009, 349, 1236–1245. [Google Scholar] [CrossRef]
- Harrington, T.; Kuehnert, M.J.; Kamel, H.; Lanciotti, R.S.; Hand, S.; Currier, M.; Chamberland, M.E.; Petersen, L.R.; Marfin, A.A. West Nile Virus Infection Transmitted by Blood Transfusion. Transfusion 2003, 43, 1018–1022. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Intrauterine West Nile Virus Infection—New York. 2002. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm5150a3.htm (accessed on 18 August 2021).
- Martins-Acebes, M.A.; Saiz, J.-C. West Nile Virus: A Re-Emerging Pathogen Revisited. World J. Virol. 2012, 1, 51–70. [Google Scholar] [CrossRef]
- Ain-Najwa, M.Y.; Yasmin, A.R.; Omar, A.R.; Arshad, S.S.; Abu, J.; Mohammed, H.O.; Kumar, K.; Loong, S.K.; Rovie-Ryan, J.J.; Mohd-Kharip-Shah, A.K. Evidence of West Nile Virus Infection in Migratory and Resident Wild Birds in West Coast of Peninsular Malaysia. One Health 2020, 10, 100134. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Virus Del Nilo Occidental. Available online: https://www.who.int/es/news-room/fact-sheets/detail/west-nile-virus (accessed on 18 August 2021).
- Morel, A.P.; Webster, A.; Zitelli, L.C.; Umeno, K.; Souza, U.A.; Prusch, F.; Anicet, M.; Marsicano, G.; Bandarra, P.; Trainini, G.; et al. Serosurvey of West Nile Virus (WNV) in Free-Ranging Raptors from Brazil. Braz. J. Microbiol. 2021, 52, 411–418. [Google Scholar] [CrossRef]
- Ciota, A.T. West Nile Virus and Its Vectors. Curr. Opin. Insect Sci. 2017, 22, 28–36. [Google Scholar] [CrossRef]
- Ribeiro, P.B.; Costa, P.R.P.; Loeck, A.E.; Vianna, E.E.; Silveira Júnior, P. Exigências Térmicas de Culex Quinquefasciatus (Diptera, Culicidae) Em Pelotas, Rio Grande Do Sul, Brasil. Iheringia Série Zool. 2004, 94, 177–180. [Google Scholar] [CrossRef]
- Harbach, R.E. Culex Pipiens: Species Versus Species Complex—Taxonomic History and Perspective. J. Am. Mosq. Control. Assoc. 2012, 28, 10–23. [Google Scholar] [CrossRef]
- Laporta, G.Z.; Urbinatti, P.R.; Natal, D. Aspectos Ecológicos Da População de Culex Quinquefasciatus Say (Diptera, Culicidae) Em Abrigos Situados No Parque Ecológico Do Tietê, São Paulo, SP. Rev. Bras. Entomol. 2006, 50, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, S.J.; Belkin, J.N. Collection Records of the Project “Mosquitoes of Middle America.” 13. South America: Brazil (BRA, BRAP, BRB), Ecuador (ECU), Peru (PER), Chile (CH). Mosq. Syst. 1979, 11, 61–118. [Google Scholar]
- Hutchings, R.S.G.; Sallum, M.A.M.; Hutchings, R.W. Mosquito (Diptera: Culicidae) Diversity of a Forest-Fragment Mosaic in the Amazon Rain Forest. J. Med. Entomol. 2011, 48, 173–187. [Google Scholar] [CrossRef]
- Hutchings, R.S.G.; Hutchings, R.W.; Sallum, M.A.M. Culicidae (Diptera, Culicomorpha) from the Western Brazilian Amazon: Juami-Japurá Ecological Station. Rev. Bras. Entomol. 2010, 54, 687–691. [Google Scholar] [CrossRef]
- Da Cardoso, J.C.; De Paula, M.B.; Fernandes, A.; Dos Santos, E.; De Almeida, M.A.B.; Da Fonseca, D.F.; Sallum, M.A.M. Novos Registros e Potencial Epidemiológico de Algumas Espécies de Mosquitos (Diptera, Culicidae), No Estado Do Rio Grande Do Sul. Rev. Soc. Bras. Med. Trop. 2010, 43, 552–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, R.S.G.; Hutchings, R.W.; Menezes, I.S.; Motta, M.D.A.; Sallum, M.A.M. Mosquitoes (Diptera: Culicidae) From the Northwestern Brazilian Amazon: Padauari River. J. Med. Entomol. 2016, 53, 1330–1347. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, R.S.G.; Hutchings, R.W.; Menezes, I.S.; Motta, M.D.A.; Sallum, M.A.M. Mosquitoes (Diptera: Culicidae) From the Northwestern Brazilian Amazon: Araçá River. J. Med. Entomol. 2018, 55, 1188–1209. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, R.S.G.; Hutchings, R.W.; Menezes, I.S.; Sallum, M.A.M. Mosquitoes (Diptera: Culicidae) From the Southwestern Brazilian Amazon: Liberdade and Gregório Rivers. J. Med. Entomol. 2020, 57, 1793–1811. [Google Scholar] [CrossRef]
- Ministério da Saúde; Secretaria de Vigilância em Saúde. Guia de Vigilância Do Culex Quinquefasciatus; Ministério da Saúde: Brasilia, Brazil, 2011; Volume 2.
- Centers of Disease Control and Prevention. Mosquito Control: West Nile Virus. Available online: https://www.cdc.gov/westnile/vectorcontrol/index.html (accessed on 13 February 2023).
- Vogels, C.B.; Goertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector Competence of European Mosquitoes for West Nile Virus. Emerg. Microbes Infect. 2017, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Vogels, C.B.F.; Fros, J.J.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J.M. Vector Competence of Northern European Culex Pipiens Biotypes and Hybrids for West Nile Virus Is Differentially Affected by Temperature. Parasites Vectors 2016, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, B.F.; Serra, O.P.; Heinen, L.B.d.S.; Zuchi, N.; De Souza, V.C.; Naveca, F.G.; Dos Santos, M.A.M.; Slhessarenko, R.D. Detection of Oropouche Virus Segment S in Patients and in Culex Quinquefasciatus in the State of Mato Grosso, Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 745–754. [Google Scholar] [CrossRef]
- Richards, S.L.; Anderson, S.L.; Lord, C.C. Vector Competence of Culex Pipiens Quinquefasciatus (Diptera: Culicidae) for West Nile Virus Isolates from Florida. Trop. Med. Int. Health 2014, 19, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.P.; José, F.; Lemos, A.; Roberto Da Silva, J. Digestão Em Insetos. In Tópicos Avançados em Entomologia Molecular; Instituto Nacional de Ciência e Tecnologia em Entomologia Molecular (INCT): Rio de Janeiro, Brazil, 2012; pp. 1–32. ISBN 978-85-916127-1-0. [Google Scholar]
- Vionette Do Amaral, R.J.; Dansa-Petretski, M. Interação Patógeno-Vetor: Dengue. In Tópicos Avançados em Entomologia Molecular; Instituto Nacional de Ciência e Tecnologia em Entomologia Molecular: Rio de Janeiro, Brazil, 2012; Volume 1, pp. 1–35. [Google Scholar]
- Sojka, D.; Franta, Z.; Horn, M.; Caffrey, C.R.; Mareš, M.; Kopáček, P. New Insights into the Machinery of Blood Digestion by Ticks. Trends Parasitol. 2013, 29, 276–285. [Google Scholar] [CrossRef]
- Silva, H.H.G.; Silva, L.G.; Lira, K.S. Metodologia de Criação, Manutenção de Adultos e Estocagem de Ovos de Aedes Aegypti (Linnaeus, 1762) Em Laboratório. Rev. Patol. Trop. J. Trop. Pathol. 1998, 27, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Kerst, A.J.; Nasci, R.S.; Godsey, M.S.; Mitchell, C.J.; Savage, H.M.; Komar, N.; Panella, N.A.; Allen, B.C.; Volpe, K.E.; et al. Rapid Detection of West Nile Virus from Human Clinical Specimens, Field-Collected Mosquitoes, and Avian Samples by a TaqMan Reverse Transcriptase-PCR Assay. J. Clin. Microbiol. 2000, 38, 4066–4071. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Kerst, A.J. Nucleic Acid Sequence-Based Amplification Assays for Rapid Detection of West Nile and St. Louis Encephalitis Viruses. J. Clin. Microbiol. 2001, 39, 4506–4513. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.B. A Method for the Isolation and Identification of Dengue Viruses, Using Mosquito Cell Cultures. Am. J. Trop. Med. Hyg. 1979, 28, 1053–1059. [Google Scholar] [CrossRef]
- Shope, R.E.; Sather, G.E. Arboviruses. In Diagnostic Procedures for Viral, Rickettsial and Chlamydial Infections; Schmidt, N.J., Lennette, D.A., Lennette, E.T., Lennette, E.H., Emmons, R.W., Eds.; American Public Health Association: Washington, DC, USA, 1979; pp. 767–814. ISBN 978-0875532202. [Google Scholar]
- Pesko, K.; Mores, C.N. Effect of Sequential Exposure on Infection and Dissemination Rates for West Nile and St. Louis Encephalitis Viruses in Culex Quinquefasciatus. Vector Borne Zoonotic Dis. 2009, 9, 281. [Google Scholar] [CrossRef]
- Salazar, M.I.; Richardson, J.H.; Sánchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue Virus Type 2: Replication and Tropisms in Orally Infected Aedes Aegypti Mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Nuñez, A.I.; Talavera, S.; Birnberg, L.; Rivas, R.; Pujol, N.; Verdún, M.; Aranda, C.; Berdugo, M.; Busquets, N. Evidence of Zika Virus Horizontal and Vertical Transmission in Aedes Albopictus from Spain but Not Infectious Virus in Saliva of the Progeny. Emerg. Microbes Infect. 2020, 9, 2236–2244. [Google Scholar] [CrossRef]
- Consoli, R.A.G.B.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária No Brasil, 1st ed.; Coimbra, C.E.A., Jr., Bori, C.M., Pessanha, C., Momen, H., Benchimol, J.L., Carvalheiro, J.d.R., Ferreira, L.F., Struchiner, M., Amarante, P., Gadelha, P., et al., Eds.; Fiocruz: Rio de Janeiro, Brazil, 1994; Volume 1, ISBN 85-85676-03-5. [Google Scholar]
- Vazeille, M.; Mousson, L.; Martin, E.; Failloux, A.-B. Orally Co-Infected Aedes Albopictus from La Reunion Island, Indian Ocean, Can Deliver Both Dengue and Chikungunya Infectious Viral Particles in Their Saliva. PLoS Negl. Trop. Dis. 2010, 4, e706. [Google Scholar] [CrossRef]
- Igarashi, A. Isolation of a Singh’s Aedes Albopictus Cell Clone Sensitive to Dengue and Chikungunya Viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef]
- Gubler, D.J.; Kuno, G.; Sather, G.E.; Velez, M.; Oliver, A. Mosquito Cell Cultures and Specific Monoclonal Antibodies in Surveillance for Dengue Viruses. Am. J. Trop. Med. Hyg. 1984, 33, 158–165. [Google Scholar] [CrossRef]
- Dulbecco, R.; Vogt, M. Some Problems of Animal Virology as Studied by the Plaque Technique. Cold Spring Harb. Symp. Quant. Biol. 1953, 18, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Couto-Lima, D.; Madec, Y.; Bersot, M.I.; Campos, S.S.; Motta, M.d.A.; Dos Santos, F.B.; Vazeille, M.; Vasconcelos, P.F.d.C.; Lourenço-de-Oliveira, R.; Failloux, A.-B. Potential Risk of Re-Emergence of Urban Transmission of Yellow Fever Virus in Brazil Facilitated by Competent Aedes Populations. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcgee, C.E.; Shustov, A.V.; Tsetsarkin, K.; Frolov, I.V.; Mason, P.W.; Vanlandingham, D.L.; Higgs, S. Infection, Dissemination, and Transmission of a West Nile Virus Green Fluorescent Protein Infectious Clone by Culex Pipiens Quinquefasciatus Mosquitoes. Vector Borne Zoonotic Dis. 2010, 10, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudeep, A.B.; Mandar, P.; Ghodke, Y.K.; Gokhale, M.D. Vector Competence of Two Indian Populations of Culex Quinquefasciatus (Diptera: Culicidae) Mosquitoes to Three West Nile Virus Strains. J. Vector Borne Dis. 2015, 52, 185–192. [Google Scholar]
- Micieli, M.V.; Matacchiero, A.C.; Muttis, E.; Fonseca, D.M.; Aliota, M.T.; Kramer, L.D. Vector Competence of Argentine Mosquitoes (Diptera: Culicidae) for West Nile Virus (Flaviviridae: Flavivirus). J. Med. Entomol. 2013, 50, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.L.; Lord, C.C.; Pesko, K.; Tabachnick, W.J. Environmental and Biological Factors Influencing Culex Pipiens Quinquefasciatus Say (Diptera: Culicidae) Vector Competence for Saint Louis Encephalitis Virus. J. Trop. Med. Hyg. 2009, 81, 264. [Google Scholar] [CrossRef] [Green Version]
- Vanlandingham, D.L.; McGee, C.E.; Klingler, K.A.; Galbraith, S.E.; Barrett, A.D.T.; Higgs, S. Short Report: Comparison of Oral Infectious Dose of West Nile Virus Isolates Representing Three Distinct Genotypes in Culex Quinquefasciatus. Am. J. Trop. Med. Hyg. 2008, 79, 951. [Google Scholar] [CrossRef]
- Vanlandingham, D.L.; McGee, C.E.; Klinger, K.A.; Vessey, N.; Fredregillo, C.; Higgs, S. Relative Susceptibilties of South Texas Mosquitoes to Infection with West Nile Virus. Am. J. Trop. Med. Hyg. 2007, 5, 925–928. [Google Scholar] [CrossRef]
- Schneider, C.A.; Calvo, E.; Peterson, K.E. Arboviruses: How Saliva Impacts the Journey from Vector to Host. Int. J. Mol. Sci. 2021, 22, 9173. [Google Scholar] [CrossRef]
- Sanchez-Vargas, I.; Olson, K.E.; Black, W.C. The Genetic Basis for Salivary Gland Barriers to Arboviral Transmission. Insects 2021, 12, 73. [Google Scholar] [CrossRef]
- Styer, L.M.; Bernard, K.A.; Kramer, L.D. Enhanced Early West Nile Virus Infection in Young Chickens Infected by Mosquito Bite: Effect of Viral Dose. Am. J. Trop. Med. Hyg. 2006, 75, 337–345. [Google Scholar] [CrossRef]
- Vanlandingham, D.L.; Schneider, B.S.; Klingler, K.; Fair, J.; Beasley, D.; Huang, J.; Hamilton, P.; Higgs, S. Real-Time Reverse Transcriptase–Polymerase Chain Reaction Quantification of West Nile Virus Transmitted by Culex Pipiens Quinquefasciatus. Am. J. Trop. Med. Hyg. 2004, 71, 120–123. [Google Scholar] [CrossRef]
- Styer, L.M.; Kent, K.A.; Albright, R.G.; Bennett, C.J.; Kramer, L.D.; Bernard, K.A. Mosquitoes Inoculate High Doses of West Nile Virus as They Probe and Feed on Live Hosts. PLoS Pathog. 2007, 3, e132. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, L.A.M.; Silva, E.V.P.d.; Dias, D.D.; Freitas, M.N.O.; Caldeira, R.D.; Araújo, P.A.d.S.; Silva, F.S.d.; Rosa Junior, J.W.; Brandão, R.C.F.; Nascimento, B.L.S.d.; et al. Vector Competence of Culex quinquefasciatus from Brazil for West Nile Virus. Trop. Med. Infect. Dis. 2023, 8, 217. https://doi.org/10.3390/tropicalmed8040217
Reis LAM, Silva EVPd, Dias DD, Freitas MNO, Caldeira RD, Araújo PAdS, Silva FSd, Rosa Junior JW, Brandão RCF, Nascimento BLSd, et al. Vector Competence of Culex quinquefasciatus from Brazil for West Nile Virus. Tropical Medicine and Infectious Disease. 2023; 8(4):217. https://doi.org/10.3390/tropicalmed8040217
Chicago/Turabian StyleReis, Lúcia Aline Moura, Eliana Vieira Pinto da Silva, Daniel Damous Dias, Maria Nazaré Oliveira Freitas, Rossela Damasceno Caldeira, Pedro Arthur da Silva Araújo, Fábio Silva da Silva, José Wilson Rosa Junior, Roberto Carlos Feitosa Brandão, Bruna Laís Sena do Nascimento, and et al. 2023. "Vector Competence of Culex quinquefasciatus from Brazil for West Nile Virus" Tropical Medicine and Infectious Disease 8, no. 4: 217. https://doi.org/10.3390/tropicalmed8040217
APA StyleReis, L. A. M., Silva, E. V. P. d., Dias, D. D., Freitas, M. N. O., Caldeira, R. D., Araújo, P. A. d. S., Silva, F. S. d., Rosa Junior, J. W., Brandão, R. C. F., Nascimento, B. L. S. d., Martins, L. C., & Neto, J. P. N. (2023). Vector Competence of Culex quinquefasciatus from Brazil for West Nile Virus. Tropical Medicine and Infectious Disease, 8(4), 217. https://doi.org/10.3390/tropicalmed8040217