
Malacological Survey and Spatial Distribution of Intermediate Host Snails in Schistosomiasis Endemic Districts of Rwanda

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Snail Sampling and Cercariae Shedding
2.3. Molecular Characterization of Cercariae
2.4. Human Infection Data Analysis
2.5. Parasitological Assessments
2.6. Data Analysis
3. Results
3.1. Snail Species Collected
3.2. Cercariae Shedding Per Snail Species
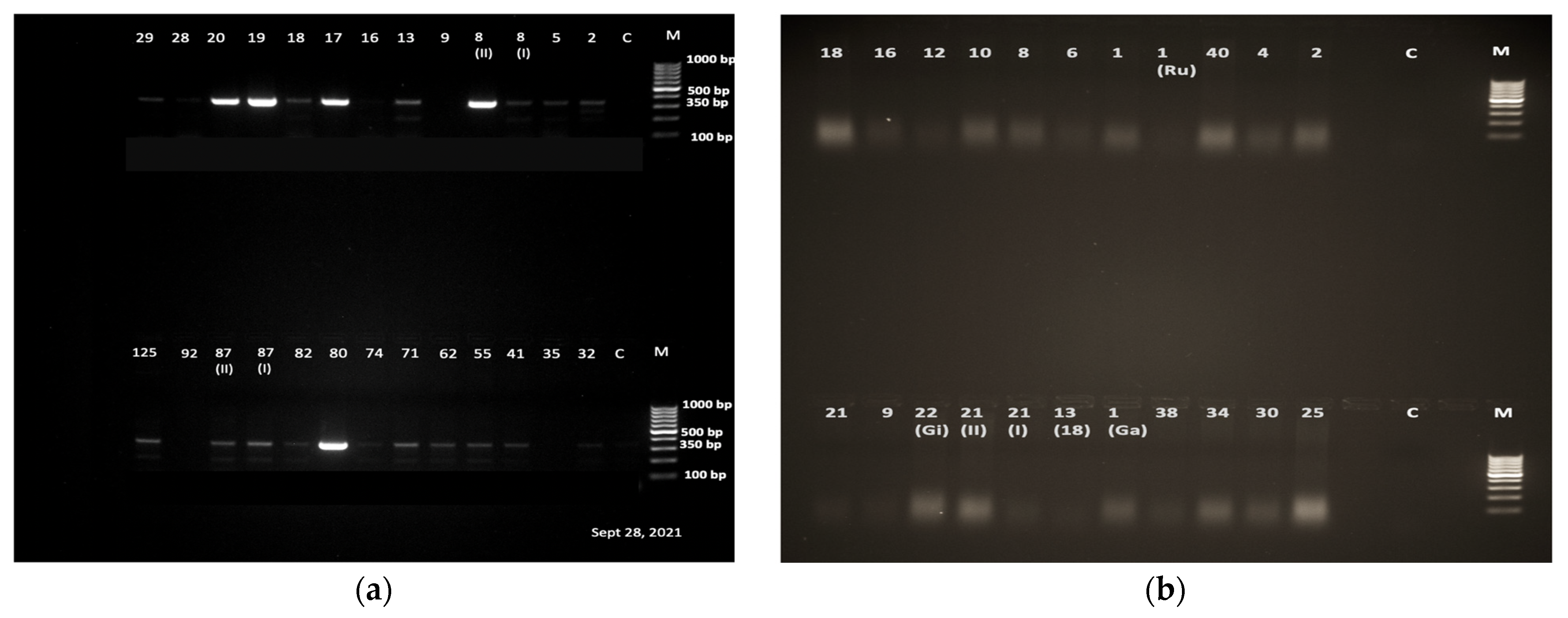
3.3. Schistosoma Cercaria Molecular Characterization
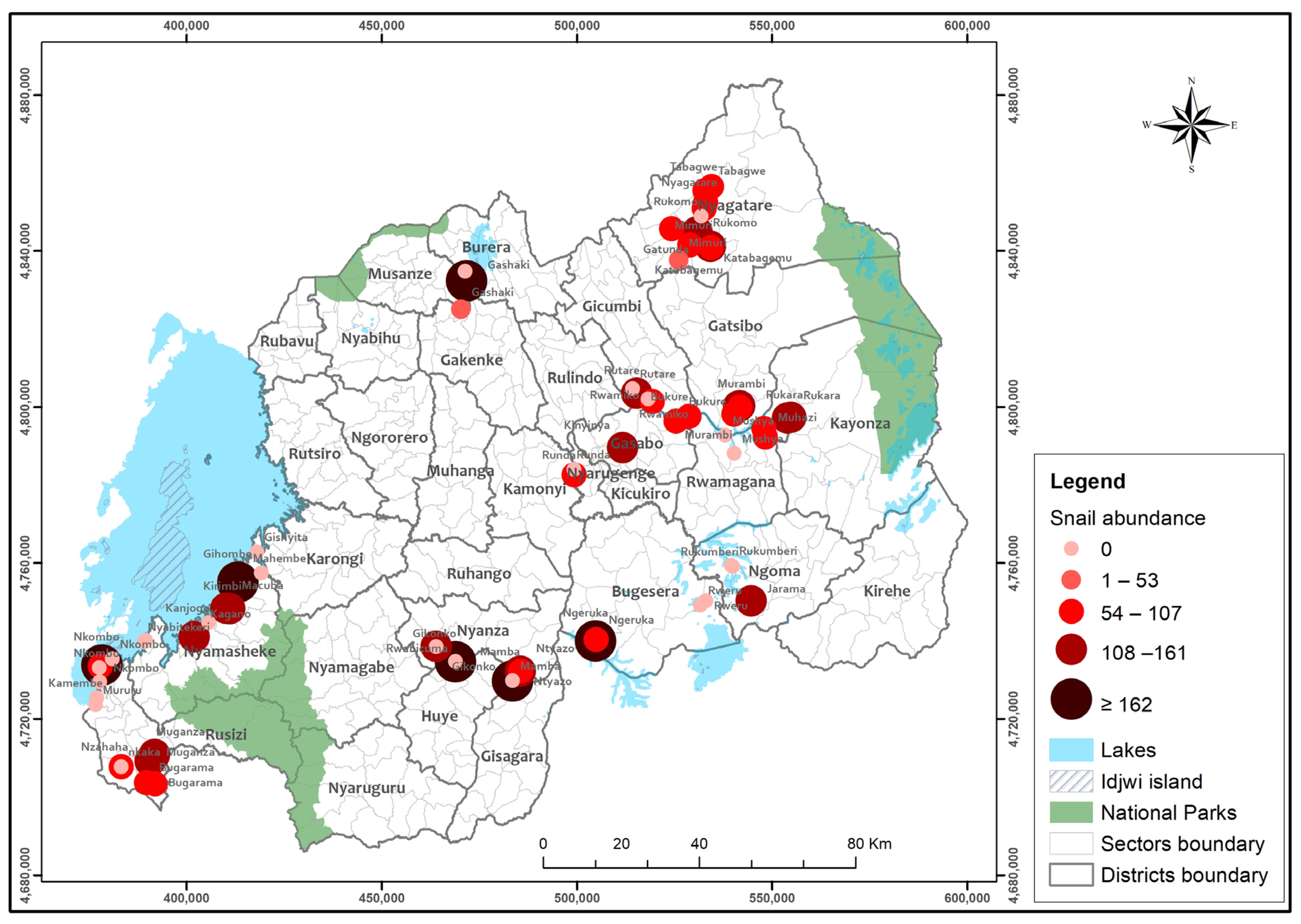
3.4. Biomphalaria and S. mansoni Spatial Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loker, E.S. Gastropod Immunobiology. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2000–2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK45994/ (accessed on 23 April 2023).
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 Diseases and Injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [PubMed]
- King, C.H.H.; Kittur, N.; Binder, S.; Campbell, C.H., Jr.; N’Goran, E.K.; Meite, A.; Utzinger, J.; Olsen, A.; Magnussen, P.; Kinung’Hi, S.; et al. Impact of different mass drug administration strategies for gaining and sustaining control of Schistosoma mansoni and Schistosoma haematobium infection in Africa. Am. J. Trop. Med. Hyg. 2020, 103, 14–23. [Google Scholar] [CrossRef]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef] [PubMed]
- Chitsulo, L.; Engels, D.; Montresor, A.; Savioli, L. The Global Status of Schistosomiasis and Its Control. Acta Trop. 2000, 77, 41–51. Available online: www.elsevier.com/locate/actatropica (accessed on 23 October 2000). [CrossRef] [PubMed]
- World Health Organization 2013; Progress Report 2001–2011 and Strategic Plan 2012–2020; World Health Organization: Geneva, Switzerland, 2010.
- Montresor, A.; Crompton, D.; Bundy, D.; Savioli, L. Guidelines for the Evaluation of Soil-Transmitted Helminthiasis and Schistosomiasis at Community Level; WHO/CTD/SIP/981; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Hotez, P.J.; Fenwick, A. Schistosomiasis in Africa: An emerging tragedy in our new global health decade. PLoS Negl. Trop Dis. 2009, 3, 9–11. [Google Scholar] [CrossRef]
- WHO/Department of Control of Neglected Tropical Diseases. Weekly Epidemiological Record Relevé Épidémiologique Hebdomadaire. Wkly Epidemiol Rec. 2018, 93, 201–220. Available online: https://apps.who.int/iris/handle/10665/272371?locale-attribute=en& (accessed on 14 December 2018).
- Nelwan, M.L. Schistosomiasis: Life Cycle, Diagnosis, and Control. Curr. Ther. Res. Clin. Exp. 2019, 91, 5–9. [Google Scholar] [CrossRef]
- Caldas, I.R.; Campi-Azevedo, A.C.; Oliveira, L.F.A.; Silveira, A.M.S.; Oliveira, R.C.; Gazzinelli, G. Human Schistosomiasis mansoni: Immune Responses during Acute and Chronic Phases of the Infection. Acta Trop. 2008, 108, 109–117. Available online: https://www.sciencedirect.com/science/article/pii/S0001706X08001733 (accessed on 23 November 2008). [CrossRef]
- Assis, A.M.O.; Barreto, M.L.; Prado, M.S.; Reis, M.G.; Parraga, I.M.; Blanton, R.E. Schistosoma mansoni infection and nutritional status in schoolchildren: A randomized, double-blind trial in northeastern Brazil. Am. J. Clin. Nutr. 1998, 68, 1247–1253. [Google Scholar] [CrossRef]
- Ezeamama, A.E.; Bustinduy, A.L.; Nkwata, A.K.; Martinez, L.; Pabalan, N.; Boivin, M.J.; King, C.H. Cognitive deficits and educational loss in children with schistosome infection—A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2018, 12, e0005524. [Google Scholar] [CrossRef]
- David, S.B. Transactions of the Royal Society of Tropical Medicine, and Hygiene. In Freshwater Snails of Africa and Their Medical Importance; CRC Press: Boca Raton, FL, USA, 1981; Volume 75, p. 384. [Google Scholar]
- Isabwe, A.; Ruberanziza, E.; Mupfasoni, D.; Ruxin, J.; Clerinx, J.; White, P.T. Potential for transmission of schistosomiasis in Kayonza district. Rwanda Med. J. 2012, 69, 14–19. [Google Scholar]
- Rujeni, N.; Morona, D.; Ruberanziza, E.; Mazigo, H.D. Schistosomiasis and soil-transmitted helminthiasis in Rwanda: An update on their epidemiology and control. Infect. Dis. Poverty 2017, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Kokaliaris, C.; Garba, A.; Matuska, M.; Bronzan, R.N.; Colley, P.D.G.; Dorkenoo, A.M.; Ekpo, U.F.; Fleming, F.M.; French, M.D.; Kabore, A.; et al. Effect of preventive chemotherapy with praziquantel on schistosomiasis among school-aged children in sub-Saharan Africa: A spatiotemporal modelling study. Lancet Infect Dis. 2022, 22, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Stothard, J.R.; Chitsulo, L.; Kristensen, T.K.; Utzinger, J. Control of schistosomiasis in sub-Saharan Africa: Progress made, new opportunities and remaining challenges. Parasitology 2009, 136, 1665–1675. [Google Scholar] [CrossRef] [PubMed]
- Van, G.Y.; Onasanya, A.; Van Engelen, J.; Oladepo, O.; Diehl, J.C. Improving access to diagnostics for schistosomiasis case management in oyo state, Nigeria: Barriers and opportunities. Diagnostics 2020, 10, 328. [Google Scholar] [CrossRef]
- Doenhoff, M.J.; Hagan, P.; Cioli, D.; Southgate, V.; Pica-Mattoccia, L.; Botros, S.; Coles, G.; Tchuenté, L.A.T.; Mbaye, A.; Engels, D. Praziquantel: Its use in control of schistosomiasis in sub-Saharan Africa and current research needs. Parasitology 2009, 136, 1825–1835. [Google Scholar] [CrossRef]
- Hoekstra, P.T.; Casacuberta, P.M.; Amoah, A.S.; van Lieshout, L.; Corstjens, P.L.A.M.; Tsonaka, S.; Assaré, R.K.; Silué, K.D.; Meite, A.; N’Goran, E.K.; et al. Repeated doses of Praziquantel in Schistosomiasis Treatment (RePST)—Single versus multiple praziquantel treatments in school-aged children in Côte d’Ivoire: A study protocol for an open-label, randomised controlled trial. BMC Infect Dis. 2018, 18, 662. [Google Scholar] [CrossRef]
- Munisi, D.Z.; Buza, J.; Mpolya, E.A.; Angelo, T.; Kinung’hi, S.M. The Efficacy of Single-Dose versus Double-Dose Praziquantel Treatments on Schistosoma mansoni Infections: Its Implication on Undernutrition and Anaemia among Primary Schoolchildren in Two on-Shore Communities, Northwestern Tanzania. Biomed. Res. Int. 2017, 2017, 7035025. [Google Scholar] [CrossRef]
- Rollinson, D.; Knopp, S.; Levitz, S.; Stothard, J.R.; Tchuem Tchuenté, L.A.; Garba, A.; Mohammed, K.A.; Schur, N.; Person, B.; Colley, D.G.; et al. Time to set the agenda for schistosomiasis elimination. Acta Trop. 2013, 128, 423–440. [Google Scholar] [CrossRef]
- Xu, J.; Li, S.Z.; Guo, J.G.; Zhou, X.N.; Djirmay, A.G. The WHO new guideline to control and eliminate human schistosomiasis: Implications for the verification of transmission interruption and surveillance of Schistosoma japonicum in China. Infect. Dis. Poverty 2022, 11, 10–12. [Google Scholar] [CrossRef]
- Grimes, J.E.T.; Croll, D.; Harrison, W.E.; Utzinger, J.; Freeman, M.C.; Templeton, M.R. The roles of water, sanitation and hygiene in reducing schistosomiasis: A review. Parasites Vectors 2015, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Nyandwi, E.; Osei, F.B.; Veldkamp, T.; Amer, S. Modeling schistosomiasis spatial risk dynamics over time in Rwanda using zero-inflated Poisson regression. Sci. Rep. 2020, 10, 19276. [Google Scholar] [CrossRef] [PubMed]
- Nyandwi, E.; Veldkamp, A.; Amer, S.; Karema, C.; Umulisa, I. Schistosomiasis mansoni incidence data in Rwanda can improve prevalence assessments, by providing high-resolution hotspot and risk factors identification. BMC Public Health 2017, 17, 845. [Google Scholar] [CrossRef] [PubMed]
- Ruberanziza, E.; Wittmann, U.; Mbituyumuremyi, A.; Mutabazi, A.; Campbell, C.H.; Colley, D.G.; Fleming, F.M.; Ortu, G.; van Dam, G.J.; Umulisa, I.; et al. Nationwide remapping of Schistosoma mansoni infection in Rwanda using circulating cathodic antigen rapid test: Taking steps toward elimination. Am. J. Trop. Med. Hyg. 2020, 103, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Rabone, M.; Wiethase, J.H.; Allan, F.; Gouvras, A.N.; Pennance, T.; Hamidou, A.A.; Webster, B.L.; Labbo, R.; Emery, A.M.; Garba, A.D.; et al. Freshwater snails of biomedical importance in the Niger River Valley: Evidence of temporal and spatial patterns in abundance, distribution, and infection with Schistosoma spp. Parasites Vectors 2019, 12, 498. [Google Scholar] [CrossRef]
- Ernould, J.C.; Ba, K.; Sellin, B. The impact of the local water-development programme on the abundance of the intermediate hosts of schistosomiasis in three villages of the Senegal River delta. Ann. Trop. Med. Parasitol. 1999, 93, 135–145. [Google Scholar]
- Niyituma, E.; Njunwa, K.J.; Mbugua, A.K.; Karema, C.; Umulisa, I.; Mkoji, G.M. Intestinal Schistosomiasis and the Associated Transmission Factors in Pre-School Aged Children in Villages Surrounding Lake Rweru in Bugesera District, Rwanda. Rwanda J. 2017, 4, 29–35. [Google Scholar] [CrossRef]
- Ruxin, J.; Negin, J. Removing the neglect from neglected tropical diseases: The Rwandan experience 2008–2010. Glob. Public Health 2012, 7, 812–822. [Google Scholar] [CrossRef]
- Madsen, H.; Bloch, P.; Phiri, H.; Kristensen, T.K.; Furu, P. Bulinus nyassanus is an intermediate host for Schistosoma haematobium in Lake Malawi. Ann. Trop. Med. Parasitol. 2001, 95, 353–360. [Google Scholar] [CrossRef]
- Ofoezie, I.E. Distribution of freshwater snails in the man-made Oyan Reservoir, Ogun State, Nigeria. Hydrobiologia 1999, 416, 181–191. [Google Scholar] [CrossRef]
- Gumble, A.; Otori, Y.; Ritchie, L.S.; Hunter, G.W. The Effect of Light, Temperature and pH on the Emergence of Schistosoma japonicum Cercariae from Oncomelania nosophora. Trans. Am. Microsc. Soc. 1957, 76, 87. [Google Scholar] [CrossRef]
- Frandsen, F.; Christensen, N.O. An introductory guide to the identification of cercariae from African freshwater snails with special reference to cercariae of trematode species of medical and veterinary importance. Acta Trop. 1984, 41, 181–202. [Google Scholar] [PubMed]
- Kalinda, C.; Chimbari, M.J.; Malatji, M.P.; Mukaratirwa, S. Influence of desiccation on the survival of Bulinus globosus under laboratory conditions. J. Freshw. Ecol. 2018, 33, 461–473. [Google Scholar] [CrossRef]
- Sandoval, N.; Siles-Lucas, M.; Pérez-Arellano, J.L.; Carranza, C.; Puente, S.; López-Abán, J.; Muro, A. A new PCR-based approach for the specific amplification of DNA from different Schistosoma species applicable to human urine samples. Parasitology 2006, 133, 581–587. [Google Scholar] [CrossRef]
- Kato-Hayashi, N.; Kirinoki, M.; Iwamura, Y.; Kanazawa, T.; Kitikoon, V.; Matsuda, H.; Chigusa, Y. Identification and differentiation of human schistosomes by polymerase chain reaction. Exp. Parasitol. 2010, 124, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, H.C.; Muchiri, E.M.; Clennon, J.A.; Pellegrini, C.; Ouma, J.H.; Sturrock, R.F.; Kitron, U.; Mungai, P.; King, C.; Ndzovu, S.T.M.; et al. Distribution patterns and cercarial shedding of Bulinus nasutus and other snails in the Msambweni area, Coast Province, Kenya. Am. J. Trop. Med. Hyg. 2004, 70, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Rujeni, N.; Bayingana, J.B.; Nyandwi, E.; Ntakarutimana, A.; Kagabo, J.; Rutayisire, R.; Shema, E.; Kanimba, P.; Mbonigaba, J.B.; Ruberanziza, E. Prevalence Mapping of Schistosoma mansoni among Pre-school Age Children in Rwanda. Front. Pediatr. 2022, 10, 906177. [Google Scholar] [CrossRef]
- Barbosa, C.S.; Gomes, E.C.S.; Marcelino, J.M.R.; Cavalcante, K.R.L.J.; Nascimento, W.R.C. Quality control of the slides by Kato-Katz method for the parasitological diagnosis of schistosomiasis infection by Schistosoma mansoni. J. Bras. Patol. Med. Lab. 2017, 53, 110–114. [Google Scholar] [CrossRef]
- Leuenberger, A.; Nassoro, T.; Said, K.; Fenner, L.; Sikalengo, G.; Letang, E.; Montresor, A.; Zhou, X.-N.; Steinmann, P.; Marti, H.; et al. Assessing stool quantities generated by three specific Kato-Katz thick smear templates employed in different settings. Infect. Dis. Poverty 2016, 5, 58. [Google Scholar] [CrossRef]
- Nalugwa, A.; Olsen, A.; Tukahebwa, M.E.; Nuwaha, F. Intestinal Schistosomiasis among Preschool Children along the Shores of Lake Victoria in Uganda. Acta Trop. 2015, 142, 115–121. Available online: https://www.sciencedirect.com/science/article/pii/S0001706X14003830 (accessed on 29 November 2015). [CrossRef]
- Clark, N.J.; Umulisa, I.; Ruberanziza, E.; Owada, K.; Colley, D.; Ortu, G.; Campbell, C.H.; Ruzindana, E.; Lancaster, W.; Mbonigaba, J.B.; et al. Mapping Schistosoma mansoni endemicity in Rwanda: A critical assessment of geographical disparities arising from circulating cathodic antigen versus Kato-Katz diagnostics. PLoS Negl. Trop. Dis. 2019, 13, e0007723. [Google Scholar] [CrossRef] [PubMed]
- Marti, H.P.; Tanner, M. Field observations on the influence of low water velocities on drifting of Bulinus globosus. Hydrobiologia 1988, 157, 119–123. [Google Scholar] [CrossRef]
- Woolhouse, M.E. Passive dispersal of Bulinus globosus. Ann. Trop. Med. Parasitol. 1988, 82, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Kalinda, C.; Chimbari, M.J. Effects of laboratory-induced desiccation on fecundity and survival of Bulinus globosus (Gastropoda: Planorbidae). Aquat. Ecol. 2022, 56, 143–152. [Google Scholar] [CrossRef]
- RoR. Rwanda Irrigation Master Plan. 2010. Available online: https://www.minagri.gov.rw/fileadmin/user_upload/Minagri/Publications/Policies_and_strategies/Rwanda_Irrigation_Master_Plan.pdf (accessed on 16 May 2023).
- Steinmann, P.; Keiser, J.; Bos, R.; Tanner, M.; Utzinger, J. Schistosomiasis and Water Resources Development: Systematic Review, Meta-Analysis, and Estimates of People at Risk. 2006. Available online: http://infection.thelancet.comvol (accessed on 6 July 2006).
- Zhou, Y.-B.; Yang, M.-X.; Yihuo, W.-L.; Liu, G.-M.; Wang, H.-Y.; Wei, J.-G.; Jiang, Q.-W. Effect of habitat fragmentation on the schistosome-transmitting snail Oncomelania hupensis in a mountainous area of China. Trans. Soc. Trop. Med. Hyg. 2011, 105, 189–196. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Waterbody | Bulinus spp. | Biomphalaria spp. | ||||
|---|---|---|---|---|---|---|
| Lakeshore | Tested | Shed | Tested | Shed | PCR tested | PCR positive S. mansoni |
| 253 | 83 (32.8%) | 103 | 43 (41.7%) | 78 | 37 (47.4%) | |
| Wetland | 843 | 406(48.2%) | 332 | 76 (22.9%) | 228 | 92 (40.4%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kagabo, J.; Kalinda, C.; Nshimiyimana, P.; Mbonigaba, J.B.; Ruberanziza, E.; Nyandwi, E.; Rujeni, N. Malacological Survey and Spatial Distribution of Intermediate Host Snails in Schistosomiasis Endemic Districts of Rwanda. Trop. Med. Infect. Dis. 2023, 8, 295. https://doi.org/10.3390/tropicalmed8060295
Kagabo J, Kalinda C, Nshimiyimana P, Mbonigaba JB, Ruberanziza E, Nyandwi E, Rujeni N. Malacological Survey and Spatial Distribution of Intermediate Host Snails in Schistosomiasis Endemic Districts of Rwanda. Tropical Medicine and Infectious Disease. 2023; 8(6):295. https://doi.org/10.3390/tropicalmed8060295
Chicago/Turabian StyleKagabo, Joseph, Chester Kalinda, Project Nshimiyimana, Jean Bosco Mbonigaba, Eugene Ruberanziza, Elias Nyandwi, and Nadine Rujeni. 2023. "Malacological Survey and Spatial Distribution of Intermediate Host Snails in Schistosomiasis Endemic Districts of Rwanda" Tropical Medicine and Infectious Disease 8, no. 6: 295. https://doi.org/10.3390/tropicalmed8060295
APA StyleKagabo, J., Kalinda, C., Nshimiyimana, P., Mbonigaba, J. B., Ruberanziza, E., Nyandwi, E., & Rujeni, N. (2023). Malacological Survey and Spatial Distribution of Intermediate Host Snails in Schistosomiasis Endemic Districts of Rwanda. Tropical Medicine and Infectious Disease, 8(6), 295. https://doi.org/10.3390/tropicalmed8060295