
Effect of 3-Carene and the Micellar Formulation on Leishmania (Leishmania) amazonensis

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Micelle Preparation
2.3. Dynamic Light Scattering (DLS) Analysis
2.4. Rheological Analysis
2.5. Promastigotes Cultivation and Host Cell Cultures Maintenance
2.6. Antipromastigote Activity
2.7. Antiamastigote Activity
2.8. Cytotoxicity to the Host Cells
2.9. Transmission Electron Microscopy Analysis
2.10. Statistical Analysis
2.11. Ethics Statement
3. Results
3.1. Physical–Chemical Characterization of Micelles
3.1.1. Micelles Obtained and Micelle Size
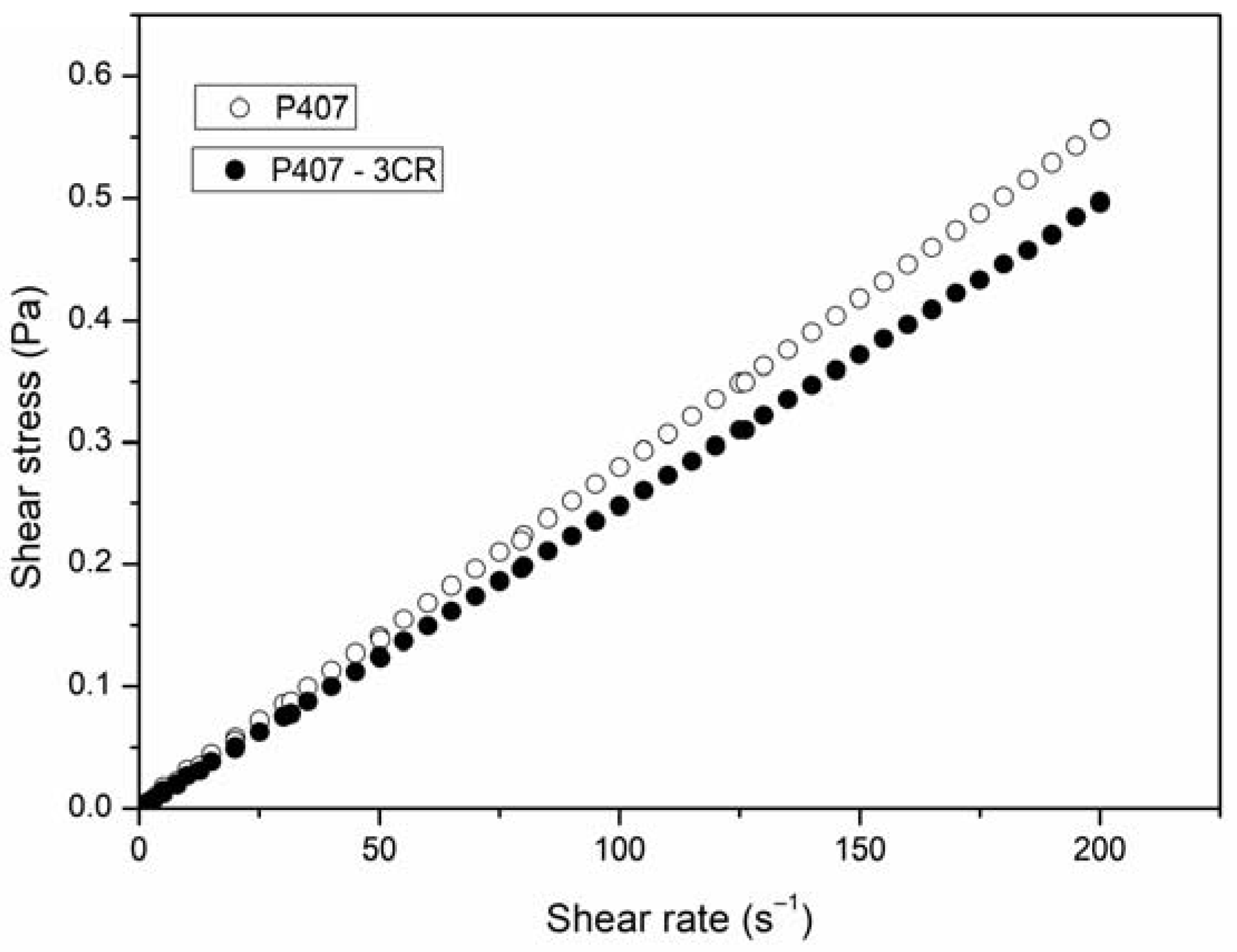
3.1.2. Rheological Behavior
3.2. Bioassays
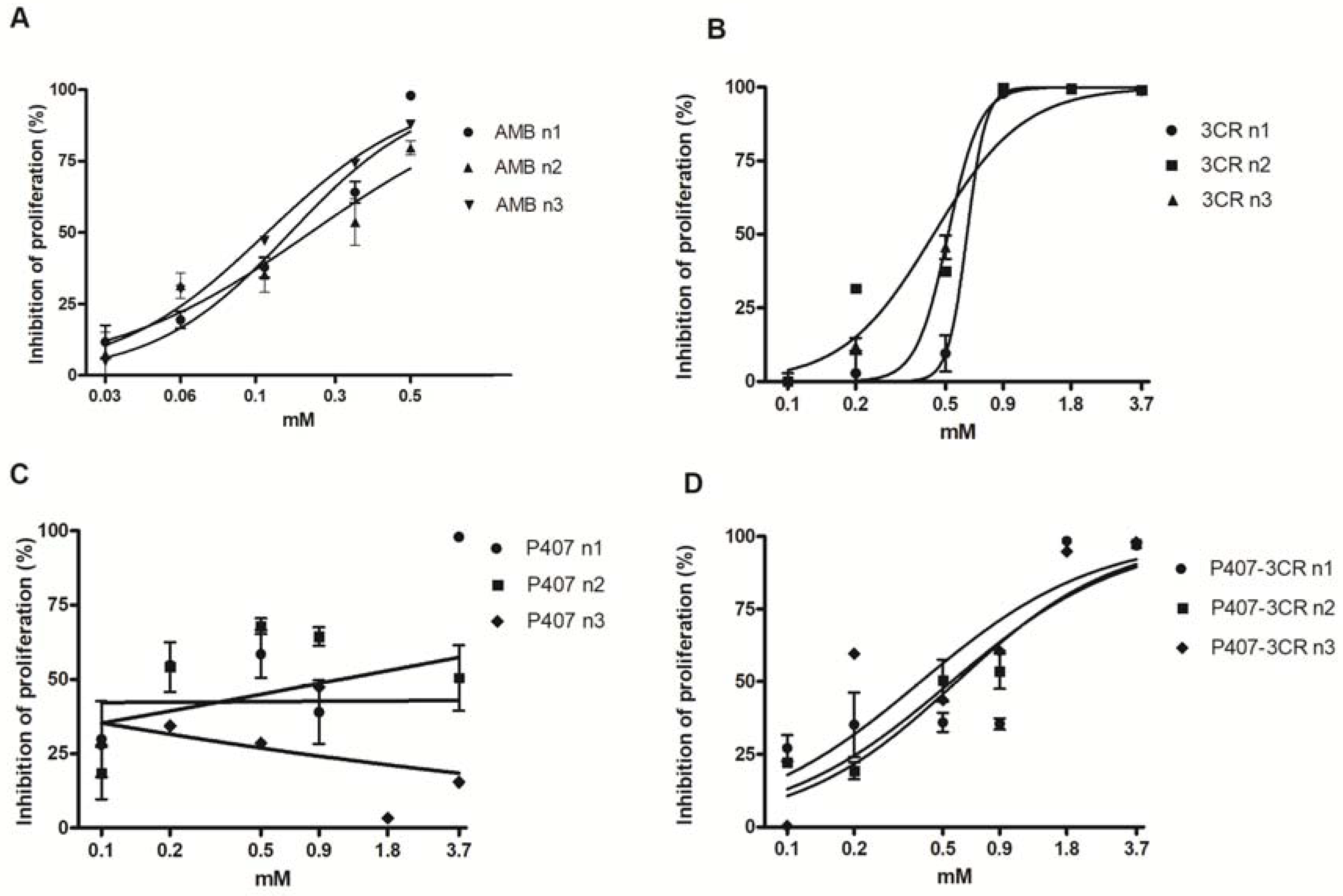
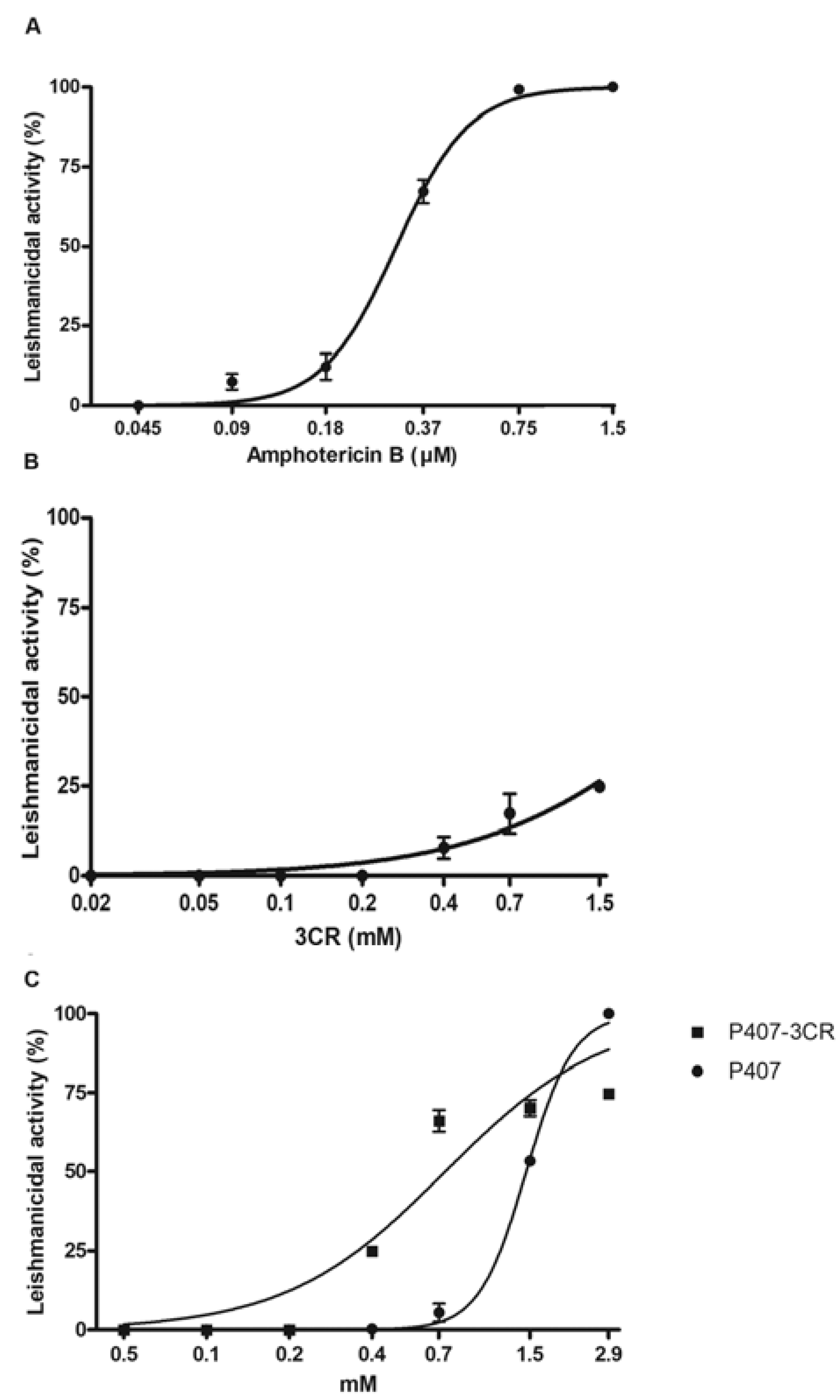
3.2.1. Leishmanicidal Activity in Promastigotes
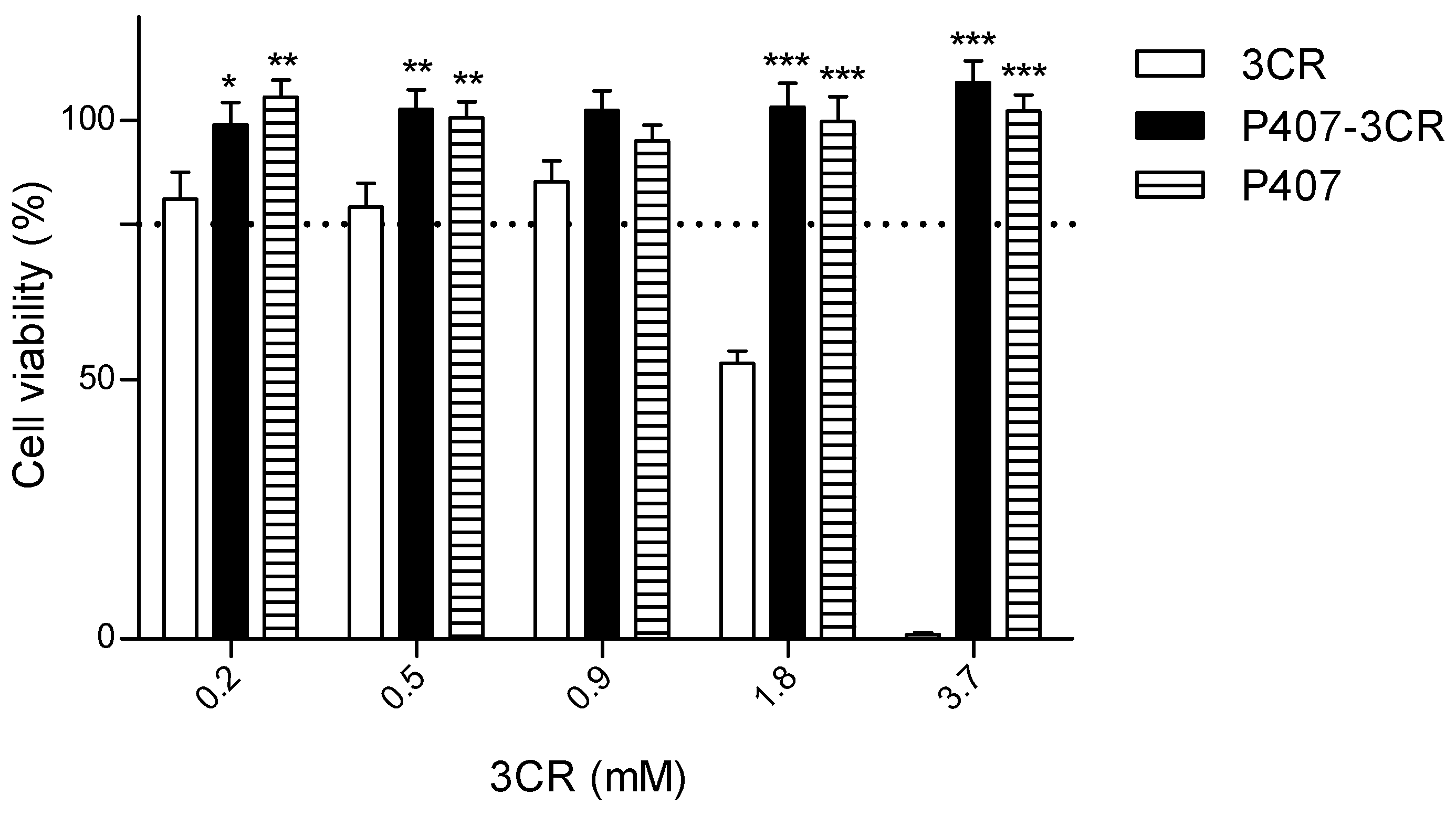
3.2.2. Leishmanicidal Activity on Intracellular Amastigotes and Cytotoxicity to Host Cells
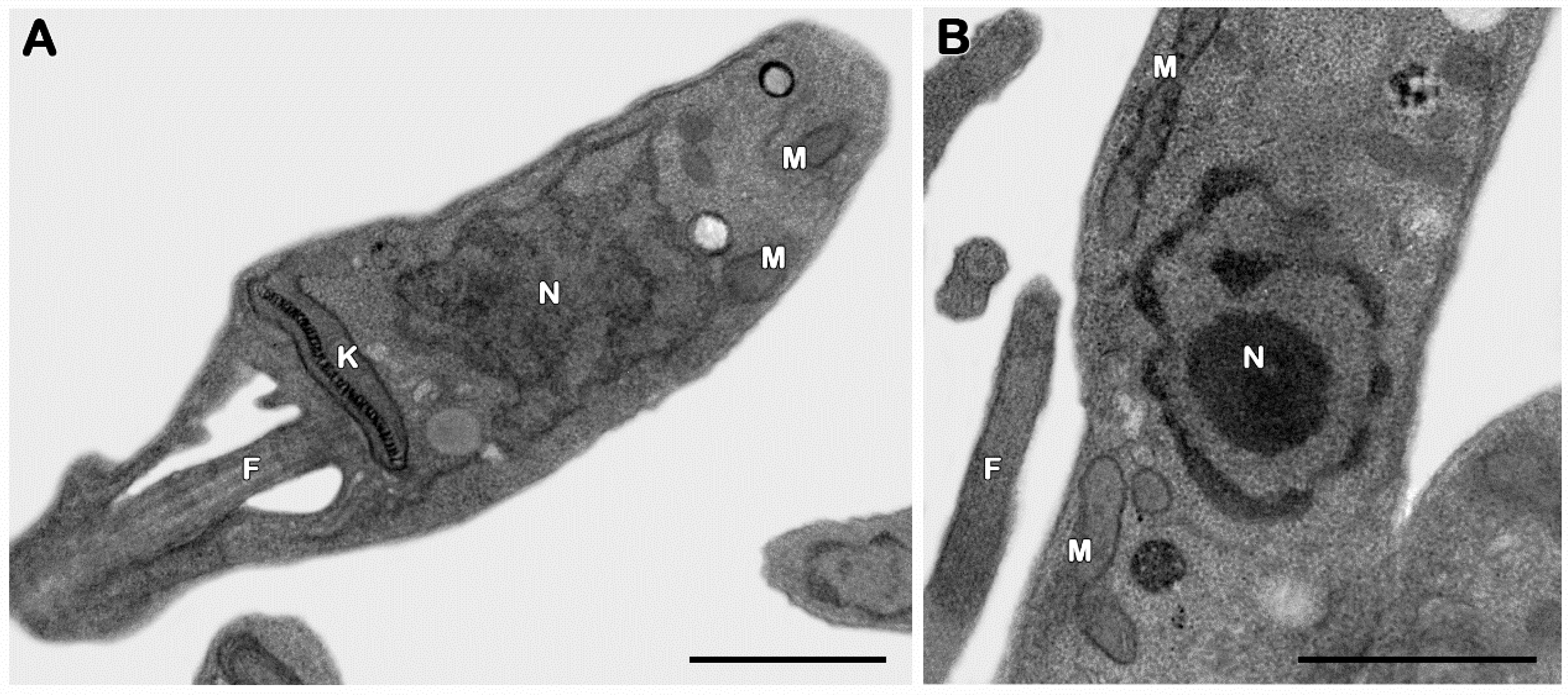
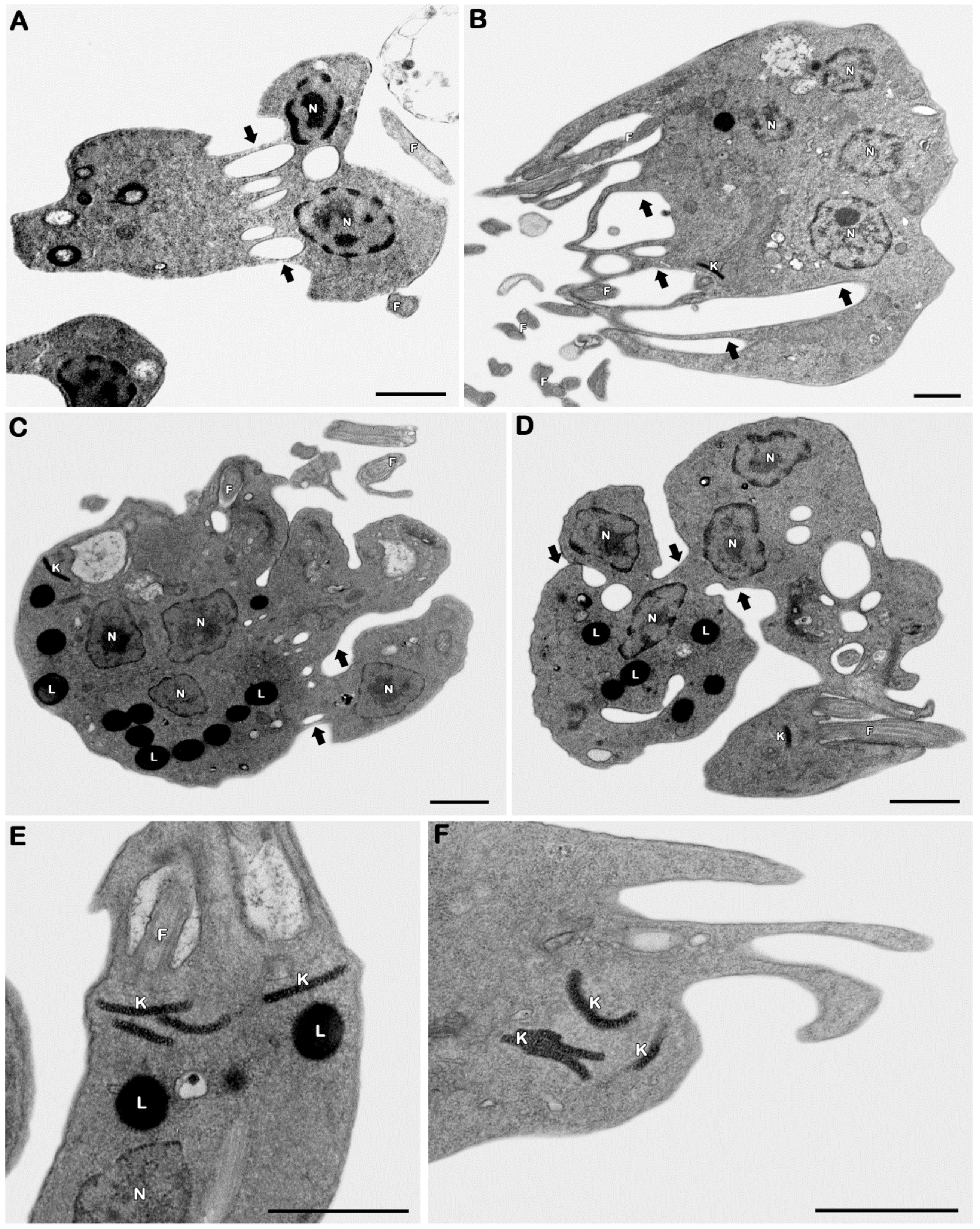
3.2.3. Ultrastructural Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brindha, J.; Balamurali, M.M.; Chanda, K. An Overview on the Therapeutics of Neglected Infectious Diseases—Leishmaniasis and Chagas Diseases. Front. Chem. 2021, 9, 622286. [Google Scholar] [CrossRef]
- WHO Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 1 March 2023).
- Mann, S.; Frasca, K.; Scherrer, S.; Henao-Martínez, A.F.; Newman, S.; Ramanan, P.; Suarez, J.A. A Review of Leishmaniasis: Current Knowledge and Future Directions. Curr. Trop. Med. Rep. 2021, 8, 121–132. [Google Scholar] [CrossRef] [PubMed]
- PAHO. WHO Leishmaniasis: Epidemiological Report for the Americas. No. 11. 12 December 2022. Available online: https://iris.paho.org/handle/10665.2/56831 (accessed on 1 March 2023).
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Bailey, F.; Mondragon-Shem, K.; Haines, L.R.; Olabi, A.; Alorfi, A.; Ruiz-Postigo, J.A.; Alvar, J.; Hotez, P.; Adams, E.R.; Vélez, I.D.; et al. Cutaneous Leishmaniasis and Co-Morbid Major Depressive Disorder: A Systematic Review with Burden Estimates. PLoS Negl. Trop. Dis. 2019, 13, e0007092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roatt, B.M.; de Oliveira Cardoso, J.M.; De Brito, R.C.F.; Coura-Vital, W.; de Oliveira Aguiar-Soares, R.D.; Reis, A.B. Recent Advances and New Strategies on Leishmaniasis Treatment. Appl. Microbiol. Biotechnol. 2020, 104, 8965–8977. [Google Scholar] [CrossRef] [PubMed]
- Scorza, B.M.; Carvalho, E.M.; Wilson, M.E. Cutaneous Manifestations of Human and Murine Leishmaniasis. Int. J. Mol. Sci. 2017, 18, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Savoia, D. Review Recent Updates and Perspectives on Leishmaniasis. J. Infect. Dev. Ctries. 2015, 9, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, S.; Abbasi, B.H.; Uzair, B.; Abbasi, R. Natural Products as Reservoirs of Novel Therapeutic Agents. EXCLI J. 2018, 17, 420–451. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.A.; Kasprzak, K.; Oniszczuk, T.; Oniszczuk, A. Natural Monoterpenes: Much More than Only a Scent. Chem. Biodivers. 2019, 16, e1900434. [Google Scholar] [CrossRef]
- Koziol, A.; Stryjewska, A.; Librowski, T.; Salat, K.; Gawel, M.; Moniczewski, A.; Lochynki, S. An Overview of the Pharmacological Properties and Potential Applications of Natural Monoterpenes. Mini-Rev. Med. Chem. 2014, 14, 1156–1168. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.G.F.; da Silva, A.C.; Citó, A.M.d.G.L.; Borges, A.R.; deLima, S.G.; Lopes, J.A.D.; Figueiredo, R.C.B.Q. In Vitro Antileishmanial Activity and Cytotoxicity of Essential Oil from Lippia Sidoides Cham. Parasitol. Int. 2011, 60, 237–241. [Google Scholar] [CrossRef] [PubMed]
- de Morais, S.M.; Vila-Nova, N.S.; Bevilaqua, C.M.; Rondon, F.C.; Lobo, C.H.; Moura, A.D.; Sales, A.D.; Rodrigues, A.P.; de Figuereido, J.R.; Campello, C.C.; et al. Thymol and Eugenol Derivatives as Potential Antileishmanial Agents. Bioorg. Med. Chem. 2014, 22, 6250–6255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, M.; Pires, P.; Dinis, A.M.; Santos-rosa, M.; Alves, V.; Salgueiro, L.; Cavaleiro, C. Monoterpenic Aldehydes as Potential Anti- Leishmania Agents: Activity of Cymbopogon Citratus and Citral on L. Infantum, L. Tropica and L. Major. Exp. Parasitol. 2012, 130, 223–231. [Google Scholar] [CrossRef]
- Youssef, M.R.; Moghaddas, E.; Tabari, M.A.; Moghadamnia, A.A.; Hosseini, S.M.; Farash, B.R.H.; Ebrahimi, M.A.; Mousavi, N.N.; Fata, A.; Maggi, F.; et al. In Vitro and In Vivo Effectiveness of Carvacrol, Thymol and Linalool against Leishmania Infantum. Molecules 2019, 11, 2072. [Google Scholar] [CrossRef] [Green Version]
- Arruda, D.C.; Miguel, D.C.; Yokoyama-yasunaka, J.K.U.; Katzin, A.M.; Uliana, S.R.B. Inhibitory Activity of Limonene against Leishmania Parasites in Vitro and in Vivo. Biomed. Pharmacother. 2009, 63, 643–649. [Google Scholar] [CrossRef]
- Api, A.M.; Belmonte, F.; Belsito, D.; Botelho, D.; Bruze, M.; Burton, G.A.; Buschmann, J.; Dagli, M.L.; Date, M.; Dekant, W.; et al. RIFM Fragrance Ingredient Safety Assessment, δ-3-Carene, CAS Registry Number 13466-78-9. Food Chem. Toxicol. 2018, 122, S771–S779. [Google Scholar] [CrossRef]
- Edman, K.; Löfstedt, H.; Berg, P.; Eriksson, K.; Axelsson, S.; Bryngelsson, I.; Fedeli, C. Exposure Assessment to α and β-Pinene, Δ3-Carene and Wood Dust in Industrial Production of Wood Pellets. Ann. Occup. Hyg. 2003, 47, 219–226. [Google Scholar] [CrossRef]
- Wang, M.; Chittiboyina, A.G.; Parcher, J.F.; Ali, Z.; Ford, P.; Zhao, J.; Avula, B.; Wang, Y.H.; Khan, I.A. Piper Nigrum Oil—Determination of Selected Terpenes for Quality Evaluation. Planta Med. 2019, 85, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.R.S.T.; Scher, R.; Santos, F.V.; Ferreira, S.R.; Cavalcanti, S.C.H.; Correa, C.B.; Bueno, L.L.; Alves, R.J.; Souza, D.P.; Fujiwara, R.T.; et al. Leishmanicidal Activity and Structure-Activity Relationships of Essential Oil Constituents. Molecules 2017, 22, 815. [Google Scholar] [CrossRef] [Green Version]
- Zheljazkov, V.D.; Cantrell, C.L.; Tekwani, B.; Khan, S.I. Content, Composition, and Bioactivity of the Essential Oils of Three Basil Genotypes as a Function of Harvesting. J. Agric. Food Chem. 2008, 56, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.; Belov, V.N.; Göen, T. Human Metabolism of Delta3-Carene and Renal Elimination of Delta3-Caren-10-Carboxylic Acid (Chaminic Acid) after Oral Administration. Arch. Toxicol. 2015, 89, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Costa, A.; Jain, S.; Coelho, E.; Fujiwara, R.; Scher, R.; Nunes, R.; Dolabella, S. Application of Poloxamers for the Development of Drug Delivery System to Treat Leishmaniasis: A Review. Curr. Drug Targets 2020, 22, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Magalhães, M.; Veiga, F.; Figueiras, A. Poloxamers, Poloxamines and Polymeric Micelles: Definition, Structure and Therapeutic Applications in Cancer. J. Polym. Res. 2018, 25, 1–14. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, E.; Yang, J.; Cao, Z. Strategies to Improve Micelle Stability for Drug Delivery. Nano Res. 2018, 11, 4985–4998. [Google Scholar] [CrossRef]
- Oyama, J.; Lera-Nonose, D.S.S.L.; Ramos-Milaré, Á.C.F.H.; Padilha Ferreira, F.B.; de Freitas, C.F.; Caetano, W.; Hioka, N.; Silveira, T.G.V.; Lonardoni, M.V.C. Potential of Pluronics® P-123 and F-127 as Nanocarriers of Anti-Leishmania Chemotherapy. Acta Trop. 2019, 192, 11–21. [Google Scholar] [CrossRef]
- Pellosi, D.S.; Moret, F.; Fraix, A.; Marino, N.; Maiolino, S.; Gaio, E.; Hioka, N.; Reddi, E.; Sortino, S.; Quaglia, F. Pluronic® P123/F127 Mixed Micelles Delivering Sorafenib and Its Combination with Verteporfin in Cancer Cells. Int. J. Nanomed. 2016, 11, 4479–4494. [Google Scholar] [CrossRef] [Green Version]
- Pereira, G.G.; Dimer, F.A.; Guterres, S.S.; Kechinski, C.P.; Granada, J.E.; Cardozo, N.S.M. Formulation and Chracterization of Poloxamer 407: Thermoreversible Gel Containing Polymeric Microparticles and Hyaluronic Acid. Quim. Nova 2013, 36, 1121–1125. [Google Scholar] [CrossRef] [Green Version]
- Akkari, A.C.S.; Papini, J.Z.B.; Garcia, G.K.; Franco, M.K.K.D.; Cavalcanti, L.P.; Gasperini, A.; Alkschbirs, M.I.; Yokaichyia, F.; De Paula, E.; Tófoli, G.R.; et al. Poloxamer 407/188 Binary Thermosensitive Hydrogels as Delivery Systems for Infiltrative Local Anesthesia: Physico-Chemical Characterization and Pharmacological Evaluation. Mater. Sci. Eng. C 2016, 68, 299–307. [Google Scholar] [CrossRef]
- Andrade-Neto, V.V.; Cunha-Júnior, E.F.; Canto-Cavalheiro, M.M.; Atella, G.C.; Fernandes, T.A.; Costa, P.R.; Torres-Santos, E.C. Antileishmanial Activity of Ezetimibe: Inhibition of Sterol Biosynthesis, In Vitro Synergy with Azoles, and Efficacy in Experimental Cutaneous Leishmaniasis. Antimicrob. Agents Chemother. 2016, 60, 6844–6852. [Google Scholar] [CrossRef] [Green Version]
- Andrade-Neto, V.V.; da Silva Pacheco, J.; Inácio, J.D.; Almeida-Amaral, E.E.; Torres-Santos, E.C.; Cunha-Junior, E.F. Efficacy of Spironolactone Treatment in Murine Models of Cutaneous and Visceral Leishmaniasis. Front. Pharmacol. 2021, 12, 636265. [Google Scholar] [CrossRef] [PubMed]
- Shamana, H.; Grossutti, M.; Papp-Szabo, E.; Miki, C.; Dutcher, J.R. Unusual Polysaccharide Rheology of Aqueous Dispersions of Soft Phytoglycogen Nanoparticles. Soft Matter 2018, 14, 6496–6505. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.K.; Hoepfner, D.; Krastel, P. Natural Products as Probes in Pharmaceutical Research. J. Ind. Microbiol. Biotechnol. 2016, 43, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Perinelli, D.R.; Cespi, M.; Pucciarelli, S.; Casettari, L.; Palmieri, G.F.; Bonacucina, G. Effect of Phosphate Buffer on the Micellisation Process of Poloxamer 407: Microcalorimetry, Acoustic Spectroscopy and Dynamic Light Scattering (DLS) Studies. Colloids Surfaces A Physicochem. Eng. Asp. 2013, 436, 123–129. [Google Scholar] [CrossRef]
- Mendonça, D.V.C.; Lage, L.M.R.; Lage, D.P.; Chávez-Fumagalli, M.A.; Ludolf, F.; Roatt, B.M.; Menezes-Souza, D.; Faraco, A.A.G.; Castilho, R.O.; Tavares, C.A.P.; et al. Poloxamer 407 (Pluronic® F127)-Based Polymeric Micelles for Amphotericin B: In Vitro Biological Activity, Toxicity and in Vivo Therapeutic Efficacy against Murine Tegumentary Leishmaniasis. Exp. Parasitol. 2016, 169, 34–42. [Google Scholar] [CrossRef]
- dos Lage, L.M.R.; Barichello, J.M.; Lage, D.P.; Mendonça, D.V.C.; Carvalho, A.M.R.S.; Rodrigues, M.R.; Menezes-Souza, D.; Roatt, B.M.; Alves, R.J.; Tavares, C.A.P.; et al. An 8-Hydroxyquinoline-Containing Polymeric Micelle System Is Effective for the Treatment of Murine Tegumentary Leishmaniasis. Parasitol. Res. 2016, 115, 4083–4095. [Google Scholar] [CrossRef]
- Tavares, G.S.V.; Mendonça, D.V.C.; Miyazaki, C.K.; Lage, D.P.; Soyer, T.G.; Carvalho, L.M.; Ottoni, F.M.; Dias, D.S.; Ribeiro, P.A.F.; Antinarelli, L.M.R.; et al. A Pluronic® F127-Based Polymeric Micelle System Containing an Antileishmanial Molecule Is Immunotherapeutic and Effective in the Treatment against Leishmania Amazonensis Infection. Parasitol. Int. 2019, 68, 63–72. [Google Scholar] [CrossRef]
- Lucia, A.; Toloza, A.C.; Guzmán, E.; Ortega, F.; Rubio, R.G. Novel Polymeric Micelles for Insect Pest Control: Encapsulation of Essential Oil Monoterpenes inside a Triblock Copolymer Shell for Head Lice Control. PeerJ 2017, 5, e3171. [Google Scholar] [CrossRef] [Green Version]
- Kulthe, S.S.; Inamdar, N.N.; Choudhari, Y.M.; Shirolikar, S.M.; Borde, L.C.; Mourya, V.K. Mixed Micelle Formation with Hydrophobic and Hydrophilic Pluronic Block Copolymers: Implications for Controlled and Targeted Drug Delivery. Colloids Surf. B Biointerfaces 2011, 88, 691–696. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, J.; Watanabe, W. Physical and Chemical Stability of Drug Nanoparticles. Adv. Drug Deliv. Rev. 2011, 63, 456–469. [Google Scholar] [CrossRef]
- Akkari, A.C.S.; Campos, E.V.R.; Keppler, A.F.; Fraceto, L.F.; de Paula, E.; Tófoli, G.R.; de Araujo, D.R. Budesonide-Hydroxypropyl-β-Cyclodextrin Inclusion Complex in Binary Poloxamer 407/403 System for Ulcerative Colitis Treatment: A Physico-Chemical Study from Micelles to Hydrogels. Colloids Surf. B Biointerfaces 2016, 138, 138–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grillo, I.; Morfin, I.; Prévost, S.F. Structural Characterization of Pluronic Micelles Swollen with Perfume Molecules. Langmuir 2018, 34, 13395–13408. [Google Scholar] [CrossRef] [PubMed]
- Fischer, E.; Fieber, W.; Navarro, C.; Sommer, H.; Benczédi, D.; Velazco, M.I.; Schönhoff, M. Partitioning and Localization of Fragrances in Surfactant Mixed Micelles. J. Surfactants Deterg. 2009, 12, 73–84. [Google Scholar] [CrossRef]
- Kabanov, A.V.; Batrakova, E.V.; Alakhov, V.Y. Pluronic Block Copolymers as Novel Polymer Therapeutics for Drug and Gene Delivery. J. Control. Release 2002, 82, 189–212. [Google Scholar] [CrossRef]
- Brugués, A.P.; Naveros, B.C.; Calpena Campmany, A.C.; Pastor, P.H.; Saladrigas, R.F.; Lizandra, C.R. Developing Cutaneous Applications of Paromomycin Entrapped in Stimuli-Sensitive Block Copolymer Nanogel Dispersions. Nanomedicine 2015, 10, 227–240. [Google Scholar] [CrossRef]
- Pasquino, R.; Droghetti, H.; Carbone, P.; Mirzaagha, S.; Grizzuti, N.; Marchisio, D. An Experimental Rheological Phase Diagram of a Tri-Block Co-Polymer in Water Validated against Dissipative Particle Dynamics Simulations. Soft Matter 2019, 15, 1396–1404. [Google Scholar] [CrossRef]
- Freitas de Souza, C.D.; Calabrese, K.D.; Abreu Silva, A.L.; Pereira Carvalho, L.O.; de Oliveira Cardoso, F.; Moraes Cavalheiros Dorval, M.E.; Teruya Oshiro, E.; Quaresma, P.F.; Ferreira Gontijo, C.M.; da Silva Pacheco, R.; et al. Leishmania Amazonensis Isolated from Human Visceral Leishmaniasis: Histopathological Analysis and Parasitological Burden in Different Inbred Mice. Histol. Histopathol. 2018, 33, 705–716. [Google Scholar] [CrossRef]
- Sanches, L.D.; Martini, C.C.; Nakamura, A.A.; Santiago, M.E.; Dolabela de Lima, B.; Lima, V.M. Natural Canine Infection by Leishmania Infantum and Leishmania Amazonensis and Their Implications for Disease Control. Rev. Bras. Parasitol. Veterinária 2016, 25, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.K.; Pawar, V.K.; Jaiswal, A.K.; Singh, Y.; Srikanth, C.H.; Chaurasia, M.; Bora, H.K.; Raval, K.; Meher, J.G.; Gayen, J.R.; et al. Chitosan Coated PluronicF127 Micelles for Effective Delivery of Amphotericin B in Experimental Visceral Leishmaniasis. Int. J. Biol. Macromol. 2017, 105, 1220–1231. [Google Scholar] [CrossRef]
- Costa, A.M.B.; Silva, A.R.S.T.; de Santos, A.J.; Galvão, J.G.; Andrade-Neto, V.V.; Torres-Santos, E.C.; Ueki, M.M.; Almeida, L.E.; Sarmento, V.H.V.; Dolabella, S.S.; et al. Thermosensitive System Formed by Poloxamers Containing Carvacrol: An Effective Carrier System against Leishmania Amazonensis. Acta Trop. 2023, 237, 106744. [Google Scholar] [CrossRef]
- Bogdan, C.; Donhauser, N.; Döring, R.; Röllinghoff, M.; Diefenbach, A.; Rittig, M.G. Fibroblasts as Host Cells in Latent Leishmaniosis. J. Exp. Med. 2000, 191, 2121–2129. [Google Scholar] [CrossRef]
- Lynch, M.D.; Watt, F.M. Fibroblast Heterogeneity: Implications for Human Disease. J. Clin. Investig. 2018, 128, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Niyompanich, J.; Chuysinuan, P.; Pavasant, P.; Supaphol, P. Development of Thermoresponsive Poloxamer in Situ Gel Loaded with Gentamicin Sulfate for Cavity Wounds. J. Polym. Res. 2021, 28, 1–13. [Google Scholar] [CrossRef]
- Borges, V.M.; Lopes, U.G.; De Souza, W.; Vannier-Santos, M.A. Cell Structure and Cytokinesis Alterations in Multidrug-Resistant Leishmania (Leishmania) Amazonensis. Parasitol. Res. 2005, 95, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Grellier, P.; Sinou, V.; Garreau-De Loubresse, N.; Bylèn, E.; Boulard, Y.; Schrével, J. Selective and Reversible Effects of Vinca Alkaloids on Trypanosoma Cruzi Epimastigote Forms: Blockage of Cytokinesis without Inhibition of the Organelle Duplication. Cell Motil. Cytoskeleton 1999, 42, 36–47. [Google Scholar] [CrossRef]
- Moulay, L.; Robert-Gero, M.; Brown, S.; Gendron, M.C.; Tournier, F. Sinefungin and Taxol Effects on Cell Cycle and Cytoskeleton of Leishmania Donovani Promastigotes. Exp. Cell Res. 1996, 226, 283–291. [Google Scholar] [CrossRef]
- Ploubidou, A.; Robinson, D.R.; Docherty, R.C.; Ogbadoyi, E.O.; Gull, K. Evidence for Novel Cell Cycle Checkpoints in Trypanosomes: Kinetoplast Segregation and Cytokinesis in the Absence of Mitosis. J. Cell Sci. 1999, 112, 4641–4650. [Google Scholar] [CrossRef]
- Stefanello, T.F.; Panice, M.R.; Ueda-Nakamura, T.; Sarragiotto, M.H.; Auzély-Velty, R.; Nakamura, C.V. N-Butyl-[1-(4-Methoxy)Phenyl-9H-β-Carboline]-3-Carboxamide Prevents Cytokinesis in Leishmania Amazonensis. Antimicrob. Agents Chemother. 2014, 58, 7112–7120. [Google Scholar] [CrossRef] [Green Version]
- Ambit, A.; Woods, K.L.; Cull, B.; Coombs, G.H.; Mottram, J.C. Morphological Events during the Cell Cycle of Leishmania Major. Eukaryot. Cell 2011, 10, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Havens, C.G.; Bryant, N.; Asher, L.; Lamoreaux, L.; Perfetto, S.; Brendle, J.J.; Werbovetz, K.A. Cellular Effects of Leishmanial Tubulin Inhibitors on L. Donovani. Mol. Biochem. Parasitol. 2000, 110, 223–236. [Google Scholar] [CrossRef]
- Castro, E.V.; Yoneyama, K.G.; Haapalainen, E.F.; Toledo, M.S.; Takahashi, H.K.; Straus, A.H. Myriocin, a Serine Palmitoyltransferase Inhibitor, Blocks Cytokinesis in Leishmania (Viannia) Braziliensis Promastigotes. J. Eukaryot. Microbiol. 2013, 60, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Godinho, J.L.P.; Georgikopoulou, K.; Calogeropoulou, T.; de Souza, W.; Rodrigues, J.C.F. A Novel Alkyl Phosphocholine-Dinitroaniline Hybrid Molecule Exhibits Biological Activity in Vitro against Leishmania Amazonensis. Exp. Parasitol. 2013, 135, 153–165. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulations | T 1 (°C) | HD (nm) | PDI | T (°C) | HD (nm) | PDI |
|---|---|---|---|---|---|---|
| P407 | 25 | 24.16 ± 0.42 | 0.314 ± 0.014 | 37 | 18.48 ± 0.15 | 0.206 ± 0.011 |
| P407-3CR | 25 | 26.81 ± 0.82 * | 0.463 ± 0.023 | 37 | 21.17 ± 0.45 * | 0.326 ± 0.027 |
| Formulations | IC50/48 h (mM) 2 |
|---|---|
| 3CR 1 | 488.1 ± 3.7 3 |
| P407 | - |
| P407-3CR | 419.9 ± 1.5 |
| Amphotericin B | 0.0002 ± 0.1 |
| Formulations | IC50/72 h 2 (mM) | CC50/72 h 3 (mM) | SI 4 |
|---|---|---|---|
| 3CR 1 | >1.5 5 | 5.2 ± 0.5 | - |
| P407 | 1.4 ± 0.1 | 2.9 ± 0.2 | 2.0 |
| P407-3CR | 0.7 ± 0.1 | 3.7 ± 0.1 | 5.2 |
| Amphotericin B | 0.0003 ± 0.0 | >0.02 | >66.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.R.S.T.; Costa, A.M.B.; Scher, R.; Andrade-Neto, V.V.; Sarmento, V.H.V.; Santos, A.d.J.; Torres-Santos, E.C.; Jain, S.; Nunes, R.d.S.; Menna-Barreto, R.F.S.; et al. Effect of 3-Carene and the Micellar Formulation on Leishmania (Leishmania) amazonensis. Trop. Med. Infect. Dis. 2023, 8, 324. https://doi.org/10.3390/tropicalmed8060324
Silva ARST, Costa AMB, Scher R, Andrade-Neto VV, Sarmento VHV, Santos AdJ, Torres-Santos EC, Jain S, Nunes RdS, Menna-Barreto RFS, et al. Effect of 3-Carene and the Micellar Formulation on Leishmania (Leishmania) amazonensis. Tropical Medicine and Infectious Disease. 2023; 8(6):324. https://doi.org/10.3390/tropicalmed8060324
Chicago/Turabian StyleSilva, Audrey Rouse Soares Tavares, Amanda Mendonça Barros Costa, Ricardo Scher, Valter Viana Andrade-Neto, Victor Hugo Vitorino Sarmento, Adriana de Jesus Santos, Eduardo Caio Torres-Santos, Sona Jain, Rogéria de Souza Nunes, Rubem Figueiredo Sadok Menna-Barreto, and et al. 2023. "Effect of 3-Carene and the Micellar Formulation on Leishmania (Leishmania) amazonensis" Tropical Medicine and Infectious Disease 8, no. 6: 324. https://doi.org/10.3390/tropicalmed8060324
APA StyleSilva, A. R. S. T., Costa, A. M. B., Scher, R., Andrade-Neto, V. V., Sarmento, V. H. V., Santos, A. d. J., Torres-Santos, E. C., Jain, S., Nunes, R. d. S., Menna-Barreto, R. F. S., & Dolabella, S. S. (2023). Effect of 3-Carene and the Micellar Formulation on Leishmania (Leishmania) amazonensis. Tropical Medicine and Infectious Disease, 8(6), 324. https://doi.org/10.3390/tropicalmed8060324