
Knobs, Adhesion, and Severe Falciparum Malaria


Abstract
:1. Introduction
2. Knob Structure
2.1. KAHRP Binds to the Submembrane Cytoskeleton of the Host Erythrocyte and May Mediate Its Reorganization
2.2. The Major Cytoadherence Ligand Is Anchored into the Knobs
2.3. Other Parasite Proteins Are Associated with Knobs
3. Knob Formation
3.1. Unique Protein Trafficking Mechanisms Are Used by the Malaria Parasite to Modify the Host Erythrocyte
3.2. KAHRP Binds to the Submembrane Cytoskeleton of the Erythrocyte as a First Step in Knob Formation
3.3. Chaperones and Other Parasite Proteins May Assist in the Movement of PfEMP1 to the Host Erythrocyte Membrane
3.4. Knobs Are Assembled In Situ at the Erythrocyte Membrane
4. Var Genes and PfEMP1 Variants
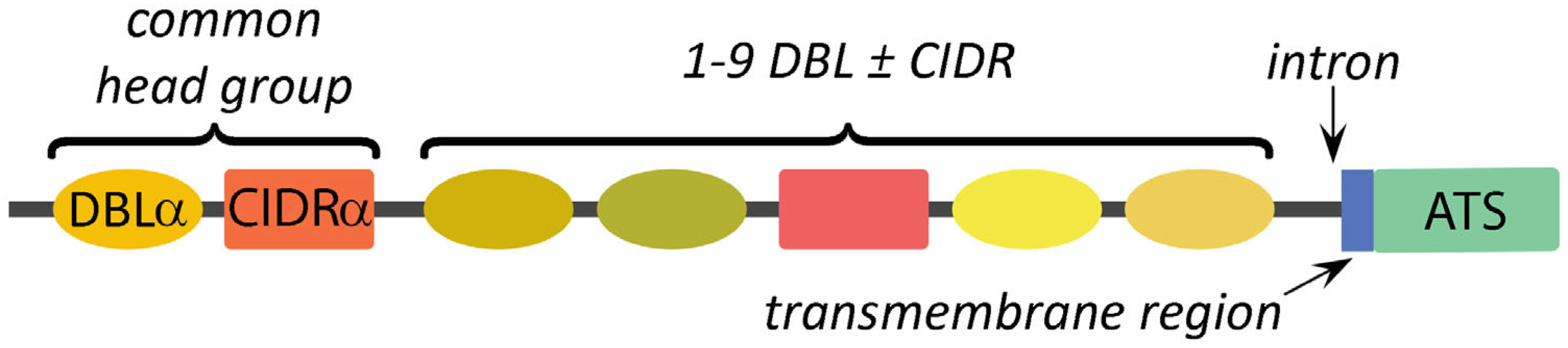
4.1. Subtypes of DBL and CIDR Modules Define the Variation between PfEMP1 Alleles
4.2. Specific Arrangements of DBL and CIDR Modules Define PfEMP1 Domain Cassettes
5. Antigenic Variation
5.1. Epigenetic Mechanisms and cis-DNA Elements Are Involved in the Monoallelic Expression of PfEMP1 Alleles
5.2. Transcripts Originating from PfEMP1 Introns Regulate Gene Expression
6. Cytoadherence Receptors
6.1. Endothelial Cell Receptors Include Adhesion Molecules and Proteins Associated with Inflammation
6.2. DBL and CIDR Domains Determine Cytoadherence Phenotype
6.3. A Single PfEMP1 Allele Binds to Chondroitin Sulfate A in the Placenta
6.4. PfEMP1 Mediates Other Cytoadherence Phenomena
7. Severe Falciparum Malaria
7.1. Cerebral Malaria, Respiratory Distress, and Severe Anemia Are Common Manifestations of Complicated Malaria
7.2. Sequestration of the Infected Erythrocytes in Microvasculature Is a Major Factor in Disease Pathogenesis
7.3. Expression of Specific PfEMP1 Alleles Is Associated with Severe Disease and Organ Specific Clinical Manifestations
8. Summary
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [Green Version]
- McQueen, P.G.; McKenzie, F.E. Age-structured red blood cell susceptibility and the dynamics of malaria infections. Proc. Natl. Acad. Sci. USA 2004, 101, 9161–9166. [Google Scholar] [CrossRef]
- Trager, W.; Rudzinska, M.A.; Bradbury, P.C. The fine structure of Plasmodium falciparum and its host erythrocytes in natural malarial infections in man. Bull. World Health Organ. 1966, 35, 883–885. [Google Scholar] [PubMed]
- Udeinya, I.J.; Schmidt, J.A.; Aikawa, M.; Miller, L.H.; Green, I. Falciparum malaria-infected erythrocytes specifically bind to cultured human endothelial cells. Science 1981, 213, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Raventos-Suarez, C.; Kaul, D.K.; Macaluso, F.; Nagel, R.L. Membrane knobs are required for the microcirculatory obstruction induced by Plasmodium falciparum-infected erythrocytes. Proc. Natl. Acad. Sci. USA 1985, 82, 3829–3833. [Google Scholar] [CrossRef]
- Warncke, J.D.; Beck, H.-P. Host cytoskeleton remodeling throughout the blood stages of Plasmodium falciparum. Microbiol. Mol. Biol. Rev. 2019, 83, e00013-19. [Google Scholar] [CrossRef] [PubMed]
- Buffet, P.A.; Safeukui, I.; Deplaine, G.; Brousse, V.; Prendki, V.; Thellier, M.; Turner, G.D.; Mercereau-Puijalon, O. The pathogenesis of Plasmodium falciparum malaria in humans: Insights from splenic physiology. Blood 2011, 117, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.D.; Rowe, J.A.; Higgins, M.K.; Lavstsen, T. Malaria’s deadly grip: Cytoadhesion of Plasmodium falciparum-infected erythrocytes. Cell. Microbiol. 2013, 15, 1976–1983. [Google Scholar] [CrossRef] [Green Version]
- Watermeyer, J.M.; Hale, V.L.; Hackett, F.; Clare, D.K.; Cutts, E.E.; Vakonakis, I.; Fleck, R.A.; Blackman, M.J.; Saibil, H.R. A spiral scaffold underlies cytoadherent knobs in Plasmodium falciparum-infected erythrocytes. Blood 2016, 127, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Kilejian, A. Characterization of a protein correlated with the production of knob-like protrusions on membranes of erythrocytes infected with Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1979, 76, 4650–4653. [Google Scholar] [CrossRef]
- Pologe, L.G.; Pavlovec, A.; Shio, H.; Ravetch, J.V. Primary structure and subcellular localization of the knob-associated histidine-rich protein of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1987, 84, 7139–7143. [Google Scholar] [CrossRef]
- Looker, O.; Blanch, A.J.; Liu, B.; Nunez-Iglesias, J.; McMillan, P.J.; Tilley, L.; Dixon, M.W.A. The knob protein KAHRP assembles into a ring-shaped structure that underpins virulence complex assembly. PLoS Pathog. 2019, 15, e1007761. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, C.P.; Patra, P.; Chang, S.-Y.S.; Karathanasis, C.; Hanebutte, L.; Kilian, N.; Cyrklaff, M.; Heilemann, M.; Schwarz, U.S.; Kudryashev, M.; et al. KAHRP dynamically relocalizes to remodeled actin junctions and associates with knob spirals in Plasmodium falciparum-infected erythrocytes. Mol. Microbiol. 2022, 117, 274–292. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rug, M.; Prescott, S.W.; Fernandez, K.M.; Cooke, B.M.; Cowman, A.F. The role of KAHRP domains in knob formation and cytoadherence of P. falciparum-infected human erythrocytes. Blood 2006, 108, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Cutts, E.E.; Laasch, N.; Reiter, D.M.; Trenker, R.; Slater, L.M.; Stansfeld, P.J.; Vakonakis, I. Structural analysis of P. falciparum KAHRP and PfEMP1 complexes with host erythrocyte spectrin suggests a model for cytoadherent knob protrusions. PLoS Pathog. 2017, 13, e1006552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cyrklaff, M.; Sanchez, C.P.; Frischknecht, F.; Lanzer, M. Host actin remodeling and protection from malaria by hemoglobinopathies. Trends Parasitol. 2012, 28, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Quadt, K.A.; Barfod, L.; Andersen, D.; Bruun, J.; Gyan, B.; Hassenkam, T.; Ofori, M.F.; Hviid, L. The density of knobs on Plasmodium falciparum-infected erythrocytes depends on developmental age and varies among isolates. PLoS ONE 2012, 7, e45658. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, C.; Kim, S.; Golkaram, M.; Dixon, M.W.A.; Tilley, L.; Li, J.; Zhang, S.; Suresh, S. Multiple stiffening effects of nanoscale knobs on human red blood cells infected with Plasmodium falciparum malaria parasite. Proc. Natl. Acad. Sci. USA 2015, 112, 6068–6073. [Google Scholar] [CrossRef]
- Jensen, A.R.; Adams, Y.; Hviid, L. Cerebral Plasmodium falciparum malaria: The role of PfEMP1 in its pathogenesis and immunity, and PfEMP1-based vaccines to prevent it. Immunol. Rev. 2020, 293, 230–252. [Google Scholar] [CrossRef] [Green Version]
- Waller, K.L.; Cooke, B.M.; Nunomura, W.; Mohandas, N.; Coppel, R.L. Mapping the binding domains involved in the interaction between the Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) and the cytoadherence ligand P. falciparum erythrocyte membrane protein 1 (PfEMP1). J. Biol. Chem. 1999, 274, 23808–23813. [Google Scholar] [CrossRef] [Green Version]
- Voigt, S.; Hanspal, M.; LeRoy, P.J.; Zhao, P.S.; Oh, S.S.; Chishti, A.H.; Liu, S.C. The cytoadherence ligand Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) binds to the P. falciparum knob-associated histidine-rich protein (KAHRP) by electrostatic interactions. Mol. Biochem. Parasitol. 2000, 110, 423–428. [Google Scholar] [CrossRef]
- Ganguly, A.K.; Ranjan, P.; Kumar, A.; Bhavesh, N.S. Dynamic association of PfEMP1 and KAHRP in knobs mediates cytoadherence during Plasmodium invasion. Sci. Rep. 2015, 5, 8617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.S.; Voigt, S.; Fisher, D.; Yi, S.J.; LeRoy, P.J.; Derick, L.H.; Liu, S.; Chishti, A.H. Plasmodium falciparum erythrocyte membrane protein 1 is anchored to the actin-spectrin junction and knob-associated histidine-rich protein in the erythrocyte skeleton. Mol. Biochem. Parasitol. 2000, 108, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Knuepfer, E.; Rug, M.; Klonis, N.; Tilley, L.; Cowman, A.F. Trafficking determinants for PfEMP3 export and assembly under the Plasmodium falciparum-infected red blood cell membrane. Mol. Microbiol. 2005, 58, 1039–1053. [Google Scholar] [CrossRef]
- Oberli, A.; Slater, L.M.; Cutts, E.; Brand, F.; Mundwiler-Pachlatko, E.; Rusch, S.; Masik, M.F.G.; Erat, M.C.; Beck, H.-P.; Vakonakis, I. A Plasmodium falciparum PHIST protein binds the virulence factor PfEMP1 and comigrates to knobs on the host cell surface. FASEB J. 2014, 28, 4420–4433. [Google Scholar] [CrossRef] [PubMed]
- Proellocks, N.I.; Herrmann, S.; Buckingham, D.W.; Hanssen, E.; Hodges, E.K.; Elsworth, B.; Morahan, B.J.; Coppel, R.L.; Cooke, B.M. A lysine-rich membrane-associated PHISTb protein involved in alteration of the cytoadhesive properties of Plasmodium falciparum-infected red blood cells. FASEB J. 2014, 28, 3103–3113. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Slater, L.; Erat, M.C.; Konrat, R.; Vakonakis, I. Structural analysis of the Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) intracellular domain reveals a conserved interaction epitope. J. Biol. Chem. 2012, 287, 7182–7189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberli, A.; Zurbrügg, L.; Rusch, S.; Brand, F.; Butler, M.E.; Day, J.L.; Cutts, E.E.; Lavstsen, T.; Vakonakis, I.; Beck, H.-P. Plasmodium falciparum Plasmodium helical interspersed subtelomeric proteins contribute to cytoadherence and anchor P. falciparum erythrocyte membrane protein 1 to the host cell cytoskeleton. Cell. Microbiol. 2016, 18, 1415–1428. [Google Scholar] [CrossRef] [Green Version]
- Alampalli, S.V.; Grover, M.; Chandran, S.; Tatu, U.; Acharya, P. Proteome and structural organization of the knob complex on the surface of the Plasmodium infected red blood cell. Proteomics Clin. Appl. 2018, 12, 1600177. [Google Scholar] [CrossRef]
- Gruenberg, J.; Allred, D.R.; Sherman, I.W. Scanning electron microscope-analysis of the protrusions (knobs) present on the surface of Plasmodium falciparum-infected erythrocytes. J. Cell Biol. 1983, 97, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Horrocks, P.; Pinches, R.A.; Chakravorty, S.J.; Papakrivos, J.; Christodoulou, Z.; Kyes, S.A.; Urban, B.C.; Ferguson, D.J.P.; Newbold, C.I. PfEMP1 expression is reduced on the surface of knobless Plasmodium falciparum infected erythrocytes. J. Cell Sci. 2005, 118, 2507–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriek, N.; Tilley, L.; Horrocks, P.; Pinches, R.; Elford, B.C.; Ferguson, D.J.P.; Lingelbach, K.; Newbold, C.I. Characterization of the pathway for transport of the cytoadherence-mediating protein, PfEMP1, to the host cell surface in malaria parasite-infected erythrocytes. Mol. Microbiol. 2003, 50, 1215–1227. [Google Scholar] [CrossRef] [Green Version]
- de Koning-Ward, T.F.; Dixon, M.W.A.; Tilley, L.; Gilson, P.R. Plasmodium species: Master renovators of their host cells. Nat. Rev. Microbiol. 2016, 14, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Counihan, N.A.; Modak, J.K.; de Koning-Ward, T.F. How malaria parasites acquire nutrients from their host. Front. Cell Dev. Biol. 2021, 9, 649184. [Google Scholar] [CrossRef]
- Wiser, M.F. Export and trafficking of Plasmodium proteins within the host erythrocyte. Acta Biológica Colomb. 2007, 12, 3–18. [Google Scholar] [CrossRef]
- Bullen, H.E.; Crabb, B.S.; Gilson, P.R. Recent insights into the export of PEXEL/HTS-motif containing proteins in Plasmodium parasites. Curr. Opin. Microbiol. 2012, 15, 699–704. [Google Scholar] [CrossRef]
- Wiser, M.F. Unique endomembrane systems and virulence in pathogenic protozoa. Life 2021, 11, 822. [Google Scholar] [CrossRef]
- Marti, M.; Good, R.T.; Rug, M.; Knuepfer, E.; Cowman, A.F. Targeting malaria virulence and remodeling proteins to the host erythrocyte. Science 2004, 306, 1930–1933. [Google Scholar] [CrossRef]
- Heiber, A.; Kruse, F.; Pick, C.; Grüring, C.; Flemming, S.; Oberli, A.; Schoeler, H.; Retzlaff, S.; Mesén-Ramírez, P.; Hiss, J.A.; et al. Identification of new PNEPs indicates a substantial non-PEXEL exportome and underpins common features in Plasmodium falciparum protein export. PLoS Pathog. 2013, 9, e1003546. [Google Scholar] [CrossRef]
- Schulze, J.; Kwiatkowski, M.; Borner, J.; Schlüter, H.; Bruchhaus, I.; Burmester, T.; Spielmann, T.; Pick, C. The Plasmodium falciparum exportome contains non-canonical PEXEL/HT proteins. Mol. Microbiol. 2015, 97, 301–314. [Google Scholar] [CrossRef]
- Matz, J.M.; Beck, J.R.; Blackman, M.J. The parasitophorous vacuole of the blood-stage malaria parasite. Nat. Rev. Microbiol. 2020, 18, 379–391. [Google Scholar] [CrossRef]
- Mundwiler-Pachlatko, E.; Beck, H.-P. Maurer’s clefts, the enigma of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2013, 110, 19987–19994. [Google Scholar] [CrossRef] [PubMed]
- Wickert, H.; Krohne, G. The complex morphology of Maurer’s clefts: From discovery to three-dimensional reconstructions. Trends Parasitol. 2007, 23, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Pachlatko, E.; Rusch, S.; Müller, A.; Hemphill, A.; Tilley, L.; Hanssen, E.; Beck, H.-P. MAHRP2, an exported protein of Plasmodium falciparum, is an essential component of Maurer’s cleft tethers. Mol. Microbiol. 2010, 77, 1136–1152. [Google Scholar] [CrossRef]
- Knuepfer, E.; Rug, M.; Klonis, N.; Tilley, L.; Cowman, A.F. Trafficking of the major virulence factor to the surface of transfected P. falciparum-infected erythrocytes. Blood 2005, 105, 4078–4087. [Google Scholar] [CrossRef] [PubMed]
- Maier, A.G.; Rug, M.; O’Neill, M.T.; Beeson, J.G.; Marti, M.; Reeder, J.; Cowman, A.F. Skeleton-binding protein 1 functions at the parasitophorous vacuole membrane to traffic PfEMP1 to the Plasmodium falciparum-infected erythrocyte surface. Blood 2007, 109, 1289–1297. [Google Scholar] [CrossRef]
- Spycher, C.; Rug, M.; Pachlatko, E.; Hanssen, E.; Ferguson, D.; Cowman, A.F.; Tilley, L.; Beck, H.-P. The Maurer’s cleft protein MAHRP1 is essential for trafficking of PfEMP1 to the surface of Plasmodium falciparum-infected erythrocytes. Mol. Microbiol. 2008, 68, 1300–1314. [Google Scholar] [CrossRef]
- Külzer, S.; Charnaud, S.; Dagan, T.; Riedel, J.; Mandal, P.; Pesce, E.R.; Blatch, G.L.; Crabb, B.S.; Gilson, P.R.; Przyborski, J.M. Plasmodium falciparum-encoded exported HSP70/HSP40 chaperone/co-chaperone complexes within the host erythrocyte. Cell. Microbiol. 2012, 14, 1784–1795. [Google Scholar] [CrossRef]
- Rug, M.; Cyrklaff, M.; Mikkonen, A.; Lemgruber, L.; Kuelzer, S.; Sanchez, C.P.; Thompson, J.; Hanssen, E.; O’Neill, M.; Langer, C.; et al. Export of virulence proteins by malaria-infected erythrocytes involves remodeling of host actin cytoskeleton. Blood 2014, 124, 3459–3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterkeyn, J.G.; Wickham, M.E.; Davern, K.M.; Cooke, B.M.; Coppel, R.L.; Reeder, J.C.; Culvenor, J.G.; Waller, R.F.; Cowman, A.F. Targeted mutagenesis of Plasmodium falciparum erythrocyte membrane protein 3 (PfEMP3) disrupts cytoadherence of malaria-infected red blood cells. EMBO J. 2000, 19, 2813–2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmo, O.M.S.; Shami, G.J.; Cox, D.; Liu, B.; Blanch, A.J.; Tiash, S.; Tilley, L.; Dixon, M.W.A. Deletion of the Plasmodium falciparum exported protein PTP7 leads to Maurer’s clefts vesiculation, host cell remodeling defects, and loss of surface presentation of EMP1. PLoS Pathog. 2022, 18, e1009882. [Google Scholar] [CrossRef] [PubMed]
- Diehl, M.; Roling, L.; Rohland, L.; Weber, S.; Cyrklaff, M.; Sanchez, C.P.; Beretta, C.A.; Simon, C.S.; Guizetti, J.; Hahn, J.; et al. Co-chaperone involvement in knob biogenesis implicates host-derived chaperones in malaria virulence. PLoS Pathog. 2021, 17, e1009969. [Google Scholar] [CrossRef] [PubMed]
- Kilejian, A.; Rashid, M.A.; Aikawa, M.; Aji, T.; Yang, Y.F. Selective association of a fragment of the knob protein with spectrin, actin and the red cell membrane. Mol. Biochem. Parasitol. 1991, 44, 175–181. [Google Scholar] [CrossRef]
- Pei, X.; An, X.; Guo, X.; Tarnawski, M.; Coppel, R.; Mohandas, N. Structural and functional studies of interaction between Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) and erythrocyte spectrin. J. Biol. Chem. 2005, 280, 31166–31171. [Google Scholar] [CrossRef] [Green Version]
- Magowan, C.; Nunomura, W.; Waller, K.L.; Yeung, J.; Liang, J.; Van Dort, H.; Low, P.S.; Coppel, R.L.; Mohandas, N. Plasmodium falciparum histidine-rich protein 1 associates with the band 3 binding domain of ankyrin in the infected red cell membrane. Biochim. Biophys. Acta-Mol. Basis Dis. 2000, 1502, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Weng, H.; Guo, X.; Papoin, J.; Wang, J.; Coppel, R.; Mohandas, N.; An, X. Interaction of Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) with erythrocyte ankyrin R is required for its attachment to the erythrocyte membrane. Biochim. Biophys. Acta 2014, 1838, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Maier, A.G.; Rug, M.; O’Neill, M.T.; Brown, M.; Chakravorty, S.; Szestak, T.; Chesson, J.; Wu, Y.; Hughes, K.; Coppel, R.L.; et al. Exported proteins required for virulence and rigidity of Plasmodium falciparum-infected human erythrocytes. Cell 2008, 134, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ma, C.; Oberli, A.; Zinz, A.; Engels, S.; Przyborski, J.M. Proteomic analysis of exported chaperone/co-chaperone complexes of P. falciparum reveals an array of complex protein-protein interactions. Sci. Rep. 2017, 7, 42188. [Google Scholar] [CrossRef] [Green Version]
- Florentin, A.; Cobb, D.W.; Kudyba, H.M.; Muralidharan, V. Directing traffic: Chaperone-mediated protein transport in malaria parasites. Cell. Microbiol. 2020, 22, e13215. [Google Scholar] [CrossRef]
- Blatch, G.L. Plasmodium falciparum molecular chaperones: Guardians of the malaria parasite proteome and renovators of the host proteome. Front. Cell Dev. Biol. 2022, 10, 921739. [Google Scholar] [CrossRef]
- Grover, M.; Chaubey, S.; Ranade, S.; Tatu, U. Identification of an exported heat shock protein 70 in Plasmodium falciparum. Parasite 2013, 20, 2. [Google Scholar] [CrossRef] [Green Version]
- Almaazmi, S.Y.; Singh, H.; Dutta, T.; Blatch, G.L. Exported J domain proteins of the human malaria parasite. Front. Mol. Biosci. 2022, 9, 978663. [Google Scholar] [CrossRef]
- Jonsdottir, T.K.; Counihan, N.A.; Modak, J.K.; Kouskousis, B.; Sanders, P.R.; Gabriela, M.; Bullen, H.E.; Crabb, B.S.; de Koning-Ward, T.F.; Gilson, P.R. Characterisation of complexes formed by parasite proteins exported into the host cell compartment of Plasmodium falciparum infected red blood cells. Cell. Microbiol. 2021, 23, e13332. [Google Scholar] [CrossRef]
- Petersen, W.; Külzer, S.; Engels, S.; Zhang, Q.; Ingmundson, A.; Rug, M.; Maier, A.G.; Przyborski, J.M. J-dot targeting of an exported HSP40 in Plasmodium falciparum-infected erythrocytes. Int. J. Parasitol. 2016, 46, 519–525. [Google Scholar] [CrossRef]
- Frankland, S.; Adisa, A.; Horrocks, P.; Taraschi, T.F.; Schneider, T.; Elliott, S.R.; Rogerson, S.J.; Knuepfer, E.; Cowman, A.F.; Newbold, C.I.; et al. Delivery of the malaria virulence protein PfEMP1 to the erythrocyte surface requires cholesterol-rich domains. Eukaryot. Cell 2006, 5, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Udeinya, I.J.; Miller, L.H.; McGregor, I.A.; Jensen, J.B. Plasmodium falciparum strain-specific antibody blocks binding of infected erythrocytes to amelanotic melanoma cells. Nature 1983, 303, 429–431. [Google Scholar] [CrossRef]
- Leech, J.H.; Barnwell, J.W.; Miller, L.H.; Howard, R.J. Identification of a strain-specific malarial antigen exposed on the surface of Plasmodium falciparum-infected erythrocytes. J. Exp. Med. 1984, 159, 1567–1575. [Google Scholar] [CrossRef]
- Howard, R.J.; Barnwell, J.W.; Rock, E.P.; Neequaye, J.; Ofori-Adjei, D.; Maloy, W.L.; Lyon, J.A.; Saul, A. Two approximately 300 kilodalton Plasmodium falciparum proteins at the surface membrane of infected erythrocytes. Mol. Biochem. Parasitol. 1988, 27, 207–223. [Google Scholar] [CrossRef]
- Su, X.Z.; Heatwole, V.M.; Wertheimer, S.P.; Guinet, F.; Herrfeldt, J.A.; Peterson, D.S.; Ravetch, J.A.; Wellems, T.E. The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infected erythrocytes. Cell 1995, 82, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Otto, T.D.; Bohme, U.; Sanders, M.J.; Reid, A.J.; Bruske, E.I.; Duffy, C.W.; Bull, P.C.; Pearson, R.D.; Abdi, A.I.; Dimonte, S.; et al. Long read assemblies of geographically dispersed Plasmodium falciparum isolates reveal highly structured subtelomeres. Wellcome Open Res. 2018, 3, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claessens, A.; Hamilton, W.L.; Kekre, M.; Otto, T.D.; Faizullabhoy, A.; Rayner, J.C.; Kwiatkowski, D. Generation of antigenic diversity in Plasmodium falciparum by structured rearrangement of var genes during mitosis. PLOS Genet. 2014, 10, e1004812. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-C.; Russell, B.; Rénia, L. Sticking for a cause: The falciparum malaria parasites cytoadherence paradigm. Front. Immunol. 2019, 10, 1444. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.S.; Miller, L.H.; Wellems, T.E. Isolation of multiple sequences from the Plasmodium falciparum genome that encode conserved domains homologous to those in erythrocyte-binding proteins. Proc. Natl. Acad. Sci. USA 1995, 92, 7100–7104. [Google Scholar] [CrossRef] [PubMed]
- Iyer, J.; Grüner, A.C.; Rénia, L.; Snounou, G.; Preiser, P.R. Invasion of host cells by malaria parasites: A tale of two protein families. Mol. Microbiol. 2007, 65, 231–249. [Google Scholar] [CrossRef]
- Baruch, D.I.; Ma, X.C.; Singh, H.B.; Bi, X.; Pasloske, B.L.; Howard, R.J. Identification of a region of PfEMP1 that mediates adherence of Plasmodium falciparum infected erythrocytes to CD36: Conserved function with variant sequence. Blood 1997, 90, 3766–3775. [Google Scholar] [CrossRef] [Green Version]
- Rask, T.S.; Hansen, D.A.; Theander, T.G.; Gorm Pedersen, A.; Lavstsen, T. Plasmodium falciparum erythrocyte membrane protein 1 diversity in seven genomes–divide and conquer. PLoS Comput. Biol. 2010, 6, e1000933. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Heddini, A.; Barragan, A.; Fernandez, V.; Pearce, S.F.; Wahlgren, M. The semiconserved head structure of Plasmodium falciparum erythrocyte membrane protein 1 mediates binding to multiple independent host receptors. J. Exp. Med. 2000, 192, 1–10. [Google Scholar] [CrossRef]
- Trimnell, A.R.; Kraemer, S.M.; Mukherjee, S.; Phippard, D.J.; Janes, J.H.; Flamoe, E.; Su, X.; Awadalla, P.; Smith, J.D. Global genetic diversity and evolution of var genes associated with placental and severe childhood malaria. Mol. Biochem. Parasitol. 2006, 148, 169–180. [Google Scholar] [CrossRef]
- Roberts, D.J.; Craig, A.G.; Berendt, A.R.; Pinches, R.; Nash, G.; Marsh, K.; Newbold, C.I. Rapid switching to multiple antigenic and adhesive phenotypes in malaria. Nature 1992, 357, 689–692. [Google Scholar] [CrossRef] [Green Version]
- Crompton, P.D.; Moebius, J.; Portugal, S.; Waisberg, M.; Hart, G.; Garver, L.S.; Miller, L.H.; Barillas-Mury, C.; Pierce, S.K. Malaria immunity in man and mosquito: Insights into unsolved mysteries of a deadly infectious disease. Annu. Rev. Immunol. 2014, 32, 157–187. [Google Scholar] [CrossRef]
- Guizetti, J.; Scherf, A. Silence, activate, poise and switch! Mechanisms of antigenic variation in Plasmodium falciparum. Cell. Microbiol. 2013, 15, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Deitsch, K.W.; Dzikowski, R. Variant gene expression and antigenic variation by malaria parasites. Annu. Rev. Microbiol. 2017, 71, 625–641. [Google Scholar] [CrossRef]
- Scherf, A.; Hernandez-Rivas, R.; Buffet, P.; Bottius, E.; Benatar, C.; Pouvelle, B.; Gysin, J.; Lanzer, M. Antigenic variation in malaria: In situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocytic development in Plasmodium falciparum. EMBO J. 1998, 17, 5418–5426. [Google Scholar] [CrossRef] [Green Version]
- Duraisingh, M.T.; Voss, T.S.; Marty, A.J.; Duffy, M.F.; Good, R.T.; Thompson, J.K.; Freitas-Junior, L.H.; Scherf, A.; Crabb, B.S.; Cowman, A.F. Heterochromatin silencing and locus repositioning linked to regulation of virulence genes in Plasmodium falciparum. Cell 2005, 121, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Ralph, S.A.; Scheidig-Benatar, C.; Scherf, A. Antigenic variation in Plasmodium falciparum is associated with movement of var loci between subnuclear locations. Proc. Natl. Acad. Sci. USA 2005, 102, 5414–5419. [Google Scholar] [CrossRef]
- Grewal, S.I.S.; Moazed, D. Heterochromatin and epigenetic control of gene expression. Science 2003, 301, 798–802. [Google Scholar] [CrossRef] [Green Version]
- Turner, B.M. Cellular memory and the histone code. Cell 2002, 111, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.; Dzikowski, R.; Costantini, D.; Amulic, B.; Berdougo, E.; Deitsch, K. Strict pairing of var promoters and introns is required for var gene silencing in the malaria parasite Plasmodium falciparum. J. Biol. Chem. 2006, 281, 9942–9952. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, M.S.; Gannoun-Zaki, L.; Wellems, T.E.; Deitsch, K.W. Plasmodium falciparum var genes are regulated by two regions with separate promoters, one upstream of the coding region and a second within the intron. J. Biol. Chem. 2003, 278, 34125–34132. [Google Scholar] [CrossRef] [Green Version]
- Epp, C.; Li, F.; Howitt, C.A.; Chookajorn, T.; Deitsch, K.W. Chromatin associated sense and antisense noncoding RNAs are transcribed from the var gene family of virulence genes of the malaria parasite Plasmodium falciparum. RNA 2009, 15, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Amit-Avraham, I.; Pozner, G.; Eshar, S.; Fastman, Y.; Kolevzon, N.; Yavin, E.; Dzikowski, R. Antisense long noncoding RNAs regulate var gene activation in the malaria parasite Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2015, 112, E982–E991. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.A.; Claessens, A.; Corrigan, R.A.; Arman, M. Adhesion of Plasmodium falciparum-infected erythrocytes to human cells: Molecular mechanisms and therapeutic implications. Expert Rev. Mol. Med. 2009, 11, e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ockenhouse, C.F.; Tandon, N.N.; Magowan, C.; Jamieson, G.A.; Chulay, J.D. Identification of a platelet membrane glycoprotein as a falciparum malaria sequestration receptor. Science 1989, 243, 1469–1471. [Google Scholar] [CrossRef] [PubMed]
- Lenahan, C.; Huang, L.; Travis, Z.D.; Zhang, J.H. Scavenger receptor class B type 1 (SR-B1) and the modifiable risk factors of stroke. Chinese Neurosurg. J. 2019, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, J.; Cui, W.; Silverstein, R.L. CD36, a signaling receptor and fatty acid transporter that regulates immune cell metabolism and fate. J. Exp. Med. 2022, 219, e20211314. [Google Scholar] [CrossRef]
- Turner, L.; Lavstsen, T.; Berger, S.S.; Wang, C.W.; Petersen, J.E.V.; Avril, M.; Brazier, A.J.; Freeth, J.; Jespersen, J.S.; Nielsen, M.A.; et al. Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature 2013, 498, 502–505. [Google Scholar] [CrossRef] [Green Version]
- Pendurthi, U.R.; Rao, L.V.M. Endothelial cell protein C receptor-dependent signaling. Curr. Opin. Hematol. 2018, 25, 219–226. [Google Scholar] [CrossRef]
- Berendt, A.R.; Simmons, D.L.; Tansey, J.; Newbold, C.I.; Marsh, K. Intercellular adhesion molecule-1 is an endothelial cell adhesion receptor for Plasmodium falciparum. Nature 1989, 341, 57–59. [Google Scholar] [CrossRef]
- van de Stolpe, A.; van der Saag, P.T. Intercellular adhesion molecule-1. J. Mol. Med. 1996, 74, 13–33. [Google Scholar] [CrossRef]
- Ockenhouse, C.F.; Tegoshi, T.; Maeno, Y.; Benjamin, C.; Ho, M.; Kan, K.E.; Thway, Y.; Win, K.; Aikawa, M.; Lobb, R.R. Human vascular endothelial cell adhesion receptors for Plasmodium falciparum-infected erythrocytes: Roles for endothelial leukocyte adhesion molecule 1 and vascular cell adhesion molecule 1. J. Exp. Med. 1992, 176, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Treutiger, C.J.; Heddini, A.; Fernandez, V.; Muller, W.A.; Wahlgren, M. PECAM-1/CD31, an endothelial receptor for binding Plasmodium falciparum-infected erythrocytes. Nat. Med. 1997, 3, 1405–1408. [Google Scholar] [CrossRef]
- Pouvelle, B.; Matarazzo, V.; Jurzynski, C.; Nemeth, J.; Ramharter, M.; Rougon, G.; Gysin, J. Neural cell adhesion molecule, a new cytoadhesion receptor for Plasmodium falciparum-infected erythrocytes capable of aggregation. Infect. Immun. 2007, 75, 3516–3522. [Google Scholar] [CrossRef] [Green Version]
- Sowparani, S.; Mahalakshmi, P.; Sweety, J.P.; Francis, A.P.; Dhanalekshmi, U.M.; Selvasudha, N. Ubiquitous neural cell adhesion molecule (NCAM): Potential mechanism and valorisation in cancer pathophysiology, drug targeting and molecular transductions. Mol. Neurobiol. 2022, 59, 5902–5924. [Google Scholar] [CrossRef]
- Ley, K. The role of selectins in inflammation and disease. Trends Mol. Med. 2003, 9, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Senczuk, A.M.; Reeder, J.C.; Kosmala, M.M.; Ho, M. Plasmodium falciparum erythrocyte membrane protein 1 functions as a ligand for P-selectin. Blood 2001, 98, 3132–3135. [Google Scholar] [CrossRef]
- Roberts, D.D.; Sherwood, J.A.; Spitalnik, S.L.; Panton, L.J.; Howard, R.J.; Dixit, V.M.; Frazier, W.A.; Miller, L.H.; Ginsburg, V. Thrombospondin binds falciparum malaria parasitized erythrocytes and may mediate cytoadherence. Nature 1985, 318, 64–66. [Google Scholar] [CrossRef]
- Siano, J.P.; Grady, K.K.; Millet, P.; Wick, T.M. Plasmodium falciparum: Cytoadherence to alpha(v)beta3 on human microvascular endothelial cells. Am. J. Trop. Med. Hyg. 1998, 59, 77–79. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [Green Version]
- Eda, S.; Sherman, I.W. Plasmodium falciparum-infected erythrocytes bind to the RGD motif of fibronectin via the band 3-related adhesin. Exp. Parasitol. 2004, 107, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.A.; Moulds, J.M.; Newbold, C.I.; Miller, L.H. P. falciparum rosetting mediated by a parasite-variant erythrocyte membrane protein and complement-receptor 1. Nature 1997, 388, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Khera, R.; Das, N. Complement receptor 1: Disease associations and therapeutic implications. Mol. Immunol. 2009, 46, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.K.; Hafiz, A.; Banerjee, B.; Kim, K.S.; Datta, K.; Chitnis, C.E. Plasmodium falciparum uses gC1qR/HABP1/p32 as a receptor to bind to vascular endothelium and for platelet-mediated clumping. PLoS Pathog. 2007, 3, 1271–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egusquiza-Alvarez, C.A.; Robles-Flores, M. An approach to p32/gC1qR/HABP1: A multifunctional protein with an essential role in cancer. J. Cancer Res. Clin. Oncol. 2022, 148, 1831–1854. [Google Scholar] [CrossRef]
- Hatabu, T.; Kawazu, S.-I.; Aikawa, M.; Kano, S. Binding of Plasmodium falciparum-infected erythrocytes to the membrane-bound form of fractalkine/CX3CL1. Proc. Natl. Acad. Sci. USA 2003, 100, 15942–15946. [Google Scholar] [CrossRef]
- Rogerson, S.J.; Chaiyaroj, S.C.; Ng, K.; Reeder, J.C.; Brown, G. V Chondroitin sulfate A is a cell surface receptor for Plasmodium falciparum-infected erythrocytes. J. Exp. Med. 1995, 182, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Fried, M.; Duffy, P.E. Adherence of Plasmodium falciparum to chondroitin sulfate A in the human placenta. Science 1996, 272, 1502–1504. [Google Scholar] [CrossRef]
- Beeson, J.G.; Rogerson, S.J.; Cooke, B.M.; Reeder, J.C.; Chai, W.; Lawson, A.M.; Molyneux, M.E.; Brown, G.V. Adhesion of Plasmodium falciparum-infected erythrocytes to hyaluronic acid in placental malaria. Nat. Med. 2000, 6, 86–90. [Google Scholar] [CrossRef]
- Chen, Q.; Barragan, A.; Fernandez, V.; Sundström, A.; Schlichtherle, M.; Sahlén, A.; Carlson, J.; Datta, S.; Wahlgren, M. Identification of Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) as the rosetting ligand of the malaria parasite P. falciparum. J. Exp. Med. 1998, 187, 15–23. [Google Scholar] [CrossRef]
- Carlson, J.; Wahlgren, M. Plasmodium falciparum erythrocyte rosetting is mediated by promiscuous lectin-like interactions. J. Exp. Med. 1992, 176, 1311–1317. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, A.; Metwally, N.G.; Allweier, J.; Cronshagen, J.; Del Pilar Martinez Tauler, M.; Murk, A.; Roth, L.K.; Torabi, H.; Wu, Y.; Gutsmann, T.; et al. CD36—A host receptor necessary for malaria parasites to establish and maintain infection. Microorganisms 2022, 10, 2356. [Google Scholar] [CrossRef]
- Robinson, B.A.; Welch, T.L.; Smith, J.D. Widespread functional specialization of Plasmodium falciparum erythrocyte membrane protein 1 family members to bind CD36 analysed across a parasite genome. Mol. Microbiol. 2003, 47, 1265–1278. [Google Scholar] [CrossRef]
- Hsieh, F.-L.; Turner, L.; Bolla, J.R.; Robinson, C.V.; Lavstsen, T.; Higgins, M.K. The structural basis for CD36 binding by the malaria parasite. Nat. Commun. 2016, 7, 12837. [Google Scholar] [CrossRef]
- Springer, A.L.; Smith, L.M.; Mackay, D.Q.; Nelson, S.O.; Smith, J.D. Functional interdependence of the DBLbeta domain and c2 region for binding of the Plasmodium falciparum variant antigen to ICAM-1. Mol. Biochem. Parasitol. 2004, 137, 55–64. [Google Scholar] [CrossRef]
- Lennartz, F.; Adams, Y.; Bengtsson, A.; Olsen, R.W.; Turner, L.; Ndam, N.T.; Ecklu-Mensah, G.; Moussiliou, A.; Ofori, M.F.; Gamain, B.; et al. Structure-guided identification of a family of dual receptor-binding PfEMP1 that is associated with cerebral malaria. Cell Host Microbe 2017, 21, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Oleinikov, A.V.; Amos, E.; Frye, I.T.; Rossnagle, E.; Mutabingwa, T.K.; Fried, M.; Duffy, P.E. High throughput functional assays of the variant antigen PfEMP1 reveal a single domain in the 3D7 Plasmodium falciparum genome that binds ICAM1 with high affinity and is targeted by naturally acquired neutralizing antibodies. PLOS Pathog. 2009, 5, e1000386. [Google Scholar] [CrossRef]
- Bengtsson, A.; Joergensen, L.; Rask, T.S.; Olsen, R.W.; Andersen, M.A.; Turner, L.; Theander, T.G.; Hviid, L.; Higgins, M.K.; Craig, A.; et al. A novel domain cassette identifies Plasmodium falciparum PfEMP1 proteins binding ICAM-1 and is a target of cross-reactive, adhesion-inhibitory antibodies. J. Immunol. 2013, 190, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.S.; Turner, L.; Wang, C.W.; Petersen, J.E.V.; Kraft, M.; Lusingu, J.P.A.; Mmbando, B.; Marquard, A.M.; Bengtsson, D.B.A.C.; Hviid, L.; et al. Plasmodium falciparum expressing domain cassette 5 type PfEMP1 (DC5-PfEMP1) bind PECAM1. PLoS ONE 2013, 8, e69117. [Google Scholar] [CrossRef] [Green Version]
- Magallón-Tejada, A.; Machevo, S.; Cisteró, P.; Lavstsen, T.; Aide, P.; Rubio, M.; Jiménez, A.; Turner, L.; Valmaseda, A.; Gupta, H.; et al. Cytoadhesion to gC1qR through Plasmodium falciparum erythrocyte membrane protein-1 in severe malaria. PLoS Pathog. 2016, 12, e1006011. [Google Scholar] [CrossRef] [Green Version]
- Vogt, A.M.; Barragan, A.; Chen, Q.; Kironde, F.; Spillmann, D.; Wahlgren, M. Heparan sulfate on endothelial cells mediates the binding of Plasmodium falciparum-infected erythrocytes via the DBL1alpha domain of PfEMP1. Blood 2003, 101, 2405–2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlbäck, M.; Jørgensen, L.M.; Nielsen, M.A.; Clausen, T.M.; Ditlev, S.B.; Resende, M.; Pinto, V.V.; Arnot, D.E.; Theander, T.G.; Salanti, A. The chondroitin sulfate A-binding site of the VAR2CSA protein involves multiple N-terminal domains. J. Biol. Chem. 2011, 286, 15908–15917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clausen, T.M.; Christoffersen, S.; Dahlbäck, M.; Langkilde, A.E.; Jensen, K.E.; Resende, M.; Agerbæk, M.Ø.; Andersen, D.; Berisha, B.; Ditlev, S.B.; et al. Structural and functional insight into how the Plasmodium falciparum VAR2CSA protein mediates binding to chondroitin sulfate A in placental malaria. J. Biol. Chem. 2012, 287, 23332–23345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avril, M.; Bernabeu, M.; Benjamin, M.; Brazier, A.J.; Smith, J.D. Interaction between endothelial protein-C receptor and intercellular adhesion molecule-1 to mediate binding of Plasmodium falciparum-infected erythrocytes to endothelial cells. MBio 2016, 7, e00615-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, C.J.; Craig, A.; Roberts, D.; Newbold, C.I.; Berendt, A.R. Intercellular adhesion molecule-1 and CD36 synergize to mediate adherence of Plasmodium falciparum-infected erythrocytes to cultured human microvascular endothelial cells. J. Clin. Invest. 1997, 100, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Chakravorty, S.J.; Craig, A. The role of ICAM-1 in Plasmodium falciparum cytoadherence. Eur. J. Cell Biol. 2005, 84, 15–27. [Google Scholar] [CrossRef]
- Sasisekharan, R.; Raman, R.; Prabhakar, V. Glycomics approach to structure-function relationships of glycosaminoglycans. Annu. Rev. Biomed. Eng. 2006, 8, 181–231. [Google Scholar] [CrossRef]
- Viebig, N.K.; Gamain, B.; Scheidig, C.; Lépolard, C.; Przyborski, J.; Lanzer, M.; Gysin, J.; Scherf, A. A single member of the Plasmodium falciparum var multigene family determines cytoadhesion to the placental receptor chondroitin sulphate A. EMBO Rep. 2005, 6, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.F.; Maier, A.G.; Byrne, T.J.; Marty, A.J.; Elliott, S.R.; O’Neill, M.T.; Payne, P.D.; Rogerson, S.J.; Cowman, A.F.; Crabb, B.S.; et al. VAR2CSA is the principal ligand for chondroitin sulfate A in two allogeneic isolates of Plasmodium falciparum. Mol. Biochem. Parasitol. 2006, 148, 117–124. [Google Scholar] [CrossRef]
- Viebig, N.K.; Levin, E.; Dechavanne, S.; Rogerson, S.J.; Gysin, J.; Smith, J.D.; Scherf, A.; Gamain, B. Disruption of var2csa gene impairs placental malaria associated adhesion phenotype. PLoS ONE 2007, 2, e910. [Google Scholar] [CrossRef]
- McQuaid, F.; Rowe, J.A. Rosetting revisited: A critical look at the evidence for host erythrocyte receptors in Plasmodium falciparum rosetting. Parasitology 2020, 147, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, A.; Kataria, P.; Surela, N.; Das, J. Pathophysiology of cerebral malaria: Implications of MSCs as a regenerative medicinal tool. Bioengineering 2022, 9, 263. [Google Scholar] [CrossRef] [PubMed]
- Pain, A.; Ferguson, D.J.; Kai, O.; Urban, B.C.; Lowe, B.; Marsh, K.; Roberts, D.J. Platelet-mediated clumping of Plasmodium falciparum-infected erythrocytes is a common adhesive phenotype and is associated with severe malaria. Proc. Natl. Acad. Sci. USA 2001, 98, 1805–1810. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, S.C.; Lépolard, C.; Traoré, B.; Pouvelle, B.; Gysin, J.; Grau, G.E. Platelets reorient Plasmodium falciparum-infected erythrocyte cytoadhesion to activated endothelial cells. J. Infect. Dis. 2004, 189, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassmer, S.C.; Taylor, T.; Maclennan, C.A.; Kanjala, M.; Mukaka, M.; Molyneux, M.E.; Grau, G.E. Platelet-induced clumping of Plasmodium falciparum-infected erythrocytes from Malawian patients with cerebral malaria-possible modulation in vivo by thrombocytopenia. J. Infect. Dis. 2008, 197, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Angchaisuksiri, P. Coagulopathy in malaria. Thromb. Res. 2014, 133, 5–9. [Google Scholar] [CrossRef]
- Duangchan, T.; Kotepui, M.; Sukati, S.; Rattanapan, Y.; Wangdi, K. A systematic review and meta-analysis of the proportion estimates of disseminated intravascular coagulation (DIC) in malaria. Trop. Med. Infect. Dis. 2023, 8, 289. [Google Scholar] [CrossRef]
- White, N.J.; Pukrittayakamee, S.; Hien, T.T.; Faiz, M.A.; Mokuolu, O.A.; Dondorp, A.M. Malaria. Lancet 2014, 383, 723–735. [Google Scholar] [CrossRef]
- Mer, M.; Dünser, M.W.; Giera, R.; Dondorp, A.M. Severe malaria. Current concepts and practical overview: What every intensivist should know. Intensive Care Med. 2020, 46, 907–918. [Google Scholar] [CrossRef]
- Severe malaria. Trop. Med. Int. Health 2014, 19, 7–131. [CrossRef]
- Walker, I.S.; Rogerson, S.J. Pathogenicity and virulence of malaria: Sticky problems and tricky solutions. Virulence 2023, 14, 2150456. [Google Scholar] [CrossRef]
- Dondorp, A.M.; Desakorn, V.; Pongtavornpinyo, W.; Sahassananda, D.; Silamut, K.; Chotivanich, K.; Newton, P.N.; Pitisuttithum, P.; Smithyman, A.M.; White, N.J.; et al. Estimation of the total parasite biomass in acute falciparum malaria from plasma PfHRP2. PLoS Med. 2005, 2, e204. [Google Scholar] [CrossRef] [Green Version]
- Marsh, K.; Forster, D.; Waruiru, C.; Mwangi, I.; Winstanley, M.; Marsh, V.; Newton, C.; Winstanley, P.; Warn, P.; Peshu, N. Indicators of life-threatening malaria in African children. N. Engl. J. Med. 1995, 332, 1399–1404. [Google Scholar] [CrossRef]
- Erunkulu, O.A.; Hill, A.V.; Kwiatkowski, D.P.; Todd, J.E.; Iqbal, J.; Berzins, K.; Riley, E.M.; Greenwood, B.M. Severe malaria in Gambian children is not due to lack of previous exposure to malaria. Clin. Exp. Immunol. 1992, 89, 296–300. [Google Scholar] [CrossRef]
- Brejt, J.A.; Golightly, L.M. Severe malaria: Update on pathophysiology and treatment. Curr. Opin. Infect. Dis. 2019, 32, 413–418. [Google Scholar] [CrossRef]
- Trivedi, S.; Chakravarty, A. Neurological complications of malaria. Curr. Neurol. Neurosci. Rep. 2022, 22, 499–513. [Google Scholar] [CrossRef]
- Patel, H.; Dunican, C.; Cunnington, A.J. Predictors of outcome in childhood Plasmodium falciparum malaria. Virulence 2020, 11, 199–221. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-H.; Stevenson, M.M. Malarial anaemia: Mechanisms and implications of insufficient erythropoiesis during blood-stage malaria. Int. J. Parasitol. 2004, 34, 1501–1516. [Google Scholar] [CrossRef]
- Katsoulis, O.; Georgiadou, A.; Cunnington, A.J. Immunopathology of acute kidney injury in severe malaria. Front. Immunol. 2021, 12, 651739. [Google Scholar] [CrossRef]
- Kaul, D.K.; Roth, E.F.J.; Nagel, R.L.; Howard, R.J.; Handunnetti, S.M. Rosetting of Plasmodium falciparum-infected red blood cells with uninfected red blood cells enhances microvascular obstruction under flow conditions. Blood 1991, 78, 812–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planche, T.; Dzeing, A.; Ngou-Milama, E.; Kombila, M.; Stacpoole, P.W. Metabolic complications of severe malaria. In Malaria: Drugs, Disease and Post-genomic Biology; Compans, R.W., Cooper, M.D., Honjo, T., Koprowski, H., Melchers, F., Oldstone, M.B.A., Olsnes, S., Potter, M., Vogt, P.K., Wagner, H., Sullivan, D.J., Krishna, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 105–136. ISBN 978-3-540-29088-9. [Google Scholar]
- Moxon, C.A.; Chisala, N.V.; Wassmer, S.C.; Taylor, T.E.; Seydel, K.B.; Molyneux, M.E.; Faragher, B.; Kennedy, N.; Toh, C.-H.; Craig, A.G.; et al. Persistent endothelial activation and inflammation after Plasmodium falciparum infection in Malawian children. J. Infect. Dis. 2014, 209, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idro, R.; Marsh, K.; John, C.C.; Newton, C.R.J. Cerebral malaria: Mechanisms of brain injury and strategies for improved neurocognitive outcome. Pediatr. Res. 2010, 68, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau, G.E.; Mackenzie, C.D.; Carr, R.A.; Redard, M.; Pizzolato, G.; Allasia, C.; Cataldo, C.; Taylor, T.E.; Molyneux, M.E. Platelet accumulation in brain microvessels in fatal pediatric cerebral malaria. J. Infect. Dis. 2003, 187, 461–466. [Google Scholar] [CrossRef]
- Montgomery, J.; Mphande, F.A.; Berriman, M.; Pain, A.; Rogerson, S.J.; Taylor, T.E.; Molyneux, M.E.; Craig, A. Differential var gene expression in the organs of patients dying of falciparum malaria. Mol. Microbiol. 2007, 65, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Storm, J.; Craig, A.G. Pathogenesis of cerebral malaria—inflammation and cytoadherence. Front. Cell. Infect. Microbiol. 2014, 4, 100. [Google Scholar] [CrossRef] [Green Version]
- Ochola, L.B.; Siddondo, B.R.; Ocholla, H.; Nkya, S.; Kimani, E.N.; Williams, T.N.; Makale, J.O.; Liljander, A.; Urban, B.C.; Bull, P.C.; et al. Specific receptor usage in Plasmodium falciparum cytoadherence is associated with disease outcome. PLoS ONE 2011, 6, e14741. [Google Scholar] [CrossRef] [Green Version]
- Lavstsen, T.; Turner, L.; Saguti, F.; Magistrado, P.; Rask, T.S.; Jespersen, J.S.; Wang, C.W.; Berger, S.S.; Baraka, V.; Marquard, A.M.; et al. Plasmodium falciparum erythrocyte membrane protein 1 domain cassettes 8 and 13 are associated with severe malaria in children. Proc. Natl. Acad. Sci. USA 2012, 109, E1791-800. [Google Scholar] [CrossRef]
- Storm, J.; Jespersen, J.S.; Seydel, K.B.; Szestak, T.; Mbewe, M.; Chisala, N.V.; Phula, P.; Wang, C.W.; Taylor, T.E.; Moxon, C.A.; et al. Cerebral malaria is associated with differential cytoadherence to brain endothelial cells. EMBO Mol. Med. 2019, 11, e9164. [Google Scholar] [CrossRef]
- Avril, M.; Tripathi, A.K.; Brazier, A.J.; Andisi, C.; Janes, J.H.; Soma, V.L.; Sullivan, D.J.J.; Bull, P.C.; Stins, M.F.; Smith, J.D. A restricted subset of var genes mediates adherence of Plasmodium falciparum-infected erythrocytes to brain endothelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, E1782–E1790. [Google Scholar] [CrossRef]
- Claessens, A.; Adams, Y.; Ghumra, A.; Lindergard, G.; Buchan, C.C.; Andisi, C.; Bull, P.C.; Mok, S.; Gupta, A.P.; Wang, C.W.; et al. A subset of group A-like var genes encodes the malaria parasite ligands for binding to human brain endothelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, E1772–E1781. [Google Scholar] [CrossRef]
- Joste, V.; Guillochon, E.; Fraering, J.; Vianou, B.; Watier, L.; Jafari-Guemouri, S.; Cot, M.; Houzé, S.; Aubouy, A.; Faucher, J.F.; et al. PfEMP1 A-type ICAM-1-binding domains are not associated with cerebral malaria in Beninese children. MBio 2020, 11, e02103-20. [Google Scholar] [CrossRef] [PubMed]
- Heddini, A.; Pettersson, F.; Kai, O.; Shafi, J.; Obiero, J.; Chen, Q.; Barragan, A.; Wahlgren, M.; Marsh, K. Fresh isolates from children with severe Plasmodium falciparum malaria bind to multiple receptors. Infect. Immun. 2001, 69, 5849–5856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, A.; Hafiz, A.; Bassat, Q.; Rovira-Vallbona, E.; Sanz, S.; Machevo, S.; Aguilar, R.; Cisteró, P.; Sigaúque, B.; Menéndez, C.; et al. Association of severe malaria outcomes with platelet-mediated clumping and adhesion to a novel host receptor. PLoS ONE 2011, 6, e19422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanti, A.; Staalsoe, T.; Lavstsen, T.; Jensen, A.T.R.; Sowa, M.P.K.; Arnot, D.E.; Hviid, L.; Theander, T.G. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol. Microbiol. 2003, 49, 179–191. [Google Scholar] [CrossRef]
- Tuikue Ndam, N.G.; Salanti, A.; Bertin, G.; Dahlbäck, M.; Fievet, N.; Turner, L.; Gaye, A.; Theander, T.; Deloron, P. High level of var2csa transcription by Plasmodium falciparum isolated from the placenta. J. Infect. Dis. 2005, 192, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.F.; Caragounis, A.; Noviyanti, R.; Kyriacou, H.M.; Choong, E.K.; Boysen, K.; Healer, J.; Rowe, J.A.; Molyneux, M.E.; Brown, G.V.; et al. Transcribed var genes associated with placental malaria in Malawian women. Infect. Immun. 2006, 74, 4875–4883. [Google Scholar] [CrossRef] [Green Version]
- Rogerson, S.J.; Desai, M.; Mayor, A.; Sicuri, E.; Taylor, S.M.; van Eijk, A.M. Burden, pathology, and costs of malaria in pregnancy: New developments for an old problem. Lancet Infect. Dis. 2018, 18, e107–e118. [Google Scholar] [CrossRef]
- Ricke, C.H.; Staalsoe, T.; Koram, K.; Akanmori, B.D.; Riley, E.M.; Theander, T.G.; Hviid, L. Plasma antibodies from malaria-exposed pregnant women recognize variant surface antigens on Plasmodium falciparum-infected erythrocytes in a parity-dependent manner and block parasite adhesion to chondroitin sulfate A. J. Immunol. 2000, 165, 3309–3316. [Google Scholar] [CrossRef] [Green Version]
- Healy, S.A.; Fried, M.; Richie, T.; Bok, K.; Little, M.; August, A.; Riley, L.; Swamy, G.K.; Wylie, B.J.; Menendez, C.; et al. Malaria vaccine trials in pregnant women: An imperative without precedent. Vaccine 2019, 37, 763–770. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Possible Role |
|---|---|
| PfSBP1 [47] | May function in the formation of the Maurer’s clefts at the parasitophorous vacuole membrane and the loading of PfEMP1 onto the clefts. |
| MAHRP1 [48] | Plays a role in the export of PfEMP1 from the parasite and its association with the Maurer’s clefts. |
| PfHSP70-x [49] | A HSP70 unique to P. falciparum that is exported to the host erythrocyte where it forms a complex with PfHSP40 and PfEMP1 in structures known as J-dots. |
| PfPTP1 [50] | Disruption of this gene leads to morphological abnormalities of the Maurer’s clefts and prevents the expression of PfEMP1 on the erythrocyte surface. |
| PfEMP3 [51] | May function in the translocation of PfEMP1 from the Maurer’s clefts to the surface of the erythrocyte. |
| PTP7 [52] | Required for recruitment or formation of vesicles at the Maurer’s clefts and the transfer of PfEMP1 to the host erythrocyte membrane. |
| PFE1605w [26] | A member of the PHIST family that binds the ATS of PfEMP1 and possibly comigrates with PfEMP1 during trafficking within the infected erythrocyte. |
| PFA66 [53] | A J-domain protein (i.e., HSP40) that is needed for the proper assembly of knobs and the expression of PfEMP1 on the erythrocyte surface. May also interact with host erythrocyte HSP70. |
| Receptor | Description |
|---|---|
| CD36 [94] | A scavenger receptor [95] expressed on the surface of many cell types including endothelial cells in most tissues [96]. |
| Endothelial protein C receptor [97] | A protein on the surface of endothelial cells that binds to protein C and regulates thrombosis and inflammation [98]. |
| Intercellular adhesion molecule-1 [99] | A member of the immunoglobulin-like superfamily that is expressed on the surface of many cell types and that is upregulated during inflammation [100]. |
| Vascular cell adhesion molecule-1 [101] | A member of the immunoglobulin-like superfamily expressed on endothelial cells that plays a role in the adhesion of leukocytes [102]. |
| Platelet endothelial cell adhesion molecule-1 [103] | A member of the immunoglobulin-like superfamily that is expressed on endothelial cells as well as platelets, monocytes, and granulocytes. |
| Neural cell adhesion molecule [104] | A member of the immunoglobulin-like superfamily that plays a wide range of roles in cellular adhesion [105]. |
| Endothelial leukocyte adhesion molecule-1 [101] | A selectin, also known as E-selectin, expressed on endothelial cells that mediates adherence of leukocytes during inflammation [106]. |
| P-selectin [107] | An adhesion protein expressed on the surface of activated platelets and endothelial cells during inflammation [106]. |
| Thrombospondin [108] | A glycoprotein that is released into the blood plasma following thrombin-mediated activation of platelets that may function as a bridging molecule between infected erythrocytes and endothelial cells. |
| αVβ3 integrin [109] | A member of a large family of adhesins that facilitate cell–cell interactions and interactions with the extracellular matrix [110]. |
| Fibronectin [111] | An extracellular matrix protein. Binding may be related to the Arg–Gly–Asp (RGD) sequence present in thrombospondin and other serum proteins. |
| Complement receptor-1 [112] | A protein on the surface of erythrocytes and other cells that binds to complement proteins C3b and C4b and inhibits complement activation [113]. |
| gC1qR/HABP1/p32 [114] | A 32 kDa protein (p32) that binds multiple proteins including complement C1q, hyaluronic acid (HA), and several other proteins [115]. |
| Fractalkine [116] | A membrane-bound chemokine expressed on the surface of endothelial cells and associated with inflammation. |
| Chondroitin sulfate A [117,118] | A glycosaminoglycan that is secreted by placental cells and that lines the intervillous space that makes up the maternal–fetal interface. |
| Hyaluronic acid [119] | Another glycosaminoglycan of the placental intervillous space. |
| Heparin sulfate [120] | A glycosaminoglycan found on many cell types that may participate in rosetting. |
| Type A or B blood group antigens [121] | Trisaccharides on glycoproteins and glycolipids of the erythrocyte surface that determine ABO blood types and that may participate in rosetting. |
| Receptor | PfEMP1 Domain(s) That Bind Receptor |
|---|---|
| CD36 | DBLα/CIDRα head group [78], CIDRα2–6 [123,124] |
| Endothelial protein C receptor | CIDRα1 associated with DC8 or DC13 [97] |
| Intercellular adhesion molecule-1 | DBLβ [125,126,127], DBLβ3 in DC4 [128] |
| Platelet endothelial cell adhesion molecule-1 | DBLα/CIDRα head group or DBLδ2 [78], DC5 [129] |
| gC1qR | DBLβ12 associated with DC8 [130] |
| Complement receptor-1 | DBLα [112] |
| Heparin sulfate | DBLα/CIDRα head group [78], DBLα1 [131] |
| Blood group antigens | DBLα/CIDRα head group [78] |
| Chondroitin sulfate A | First four modules including three unique DBLs and unique CIDR in var2csa [132], second unique DBL in var2csa [133] |
| Manifestation | Features |
|---|---|
| Severe anemia | Primarily in young children and defined as hematocrit < 15% or hemoglobin < 50 g/L in the presence of parasitemia. |
| Cerebral malaria | An unrousable coma in the presence of parasitemia and not attributable to another cause. |
| Respiratory distress | Defined by labored breathing and pulmonary edema that can progress to an acute respiratory distress syndrome requiring mechanical ventilation. |
| Impaired consciousness | An impaired consciousness that is less pronounced than the unrousable coma associated with cerebral malaria. |
| Prostration or weakness | Patients are unable to sit or walk, which is not attributable to neurological or other explanations. |
| Convulsions | Three or more repeated generalized convulsions observed within 24 h. |
| Acidosis | An important cause of death due to the accumulation of organic acids, including lactic acid, and compounded by ketoacidosis and acute kidney injury. |
| Hypoglycemia | Results from increased glucose consumption in the tissues and impaired hepatic glucogenesis. Often concomitant with lactic acidosis. |
| Jaundice | Results from a combination of hemolysis and hepatocyte damage and defined by elevated serum bilirubin in the presence of parasitemia. |
| Renal impairment | Defined by low urine output and high serum creatinine or urea levels despite adequate hydration. |
| Abnormal bleeding | Recurrent or prolonged bleeding from nose, gums, or venipuncture sites. |
| Coagulopathy | Activation of blood coagulation including disseminated intravascular coagulation or depletion of platelets. |
| Circulatory collapse (shock) | Defined as systolic blood pressure < 70 mm Hg in malaria patients and accompanied by cold clammy skin. |
| Hyperpyrexia | Core body temperature > 40 °C and may be associated with rapid heart rate and occasionally delirium. |
| Hyperparasitemia | Poor prognosis associated with >10% parasitized erythrocytes. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiser, M.F. Knobs, Adhesion, and Severe Falciparum Malaria. Trop. Med. Infect. Dis. 2023, 8, 353. https://doi.org/10.3390/tropicalmed8070353
Wiser MF. Knobs, Adhesion, and Severe Falciparum Malaria. Tropical Medicine and Infectious Disease. 2023; 8(7):353. https://doi.org/10.3390/tropicalmed8070353
Chicago/Turabian StyleWiser, Mark F. 2023. "Knobs, Adhesion, and Severe Falciparum Malaria" Tropical Medicine and Infectious Disease 8, no. 7: 353. https://doi.org/10.3390/tropicalmed8070353
APA StyleWiser, M. F. (2023). Knobs, Adhesion, and Severe Falciparum Malaria. Tropical Medicine and Infectious Disease, 8(7), 353. https://doi.org/10.3390/tropicalmed8070353