Multispectral, Aerial Disease Detection for Myrtle Rust (Austropuccinia psidii) on a Lemon Myrtle Plantation

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Spectral Data
2.2. Image Processing
2.3. Data Preparation
2.4. Random Forest Classification
2.5. Accuracy Assessment
3. Results
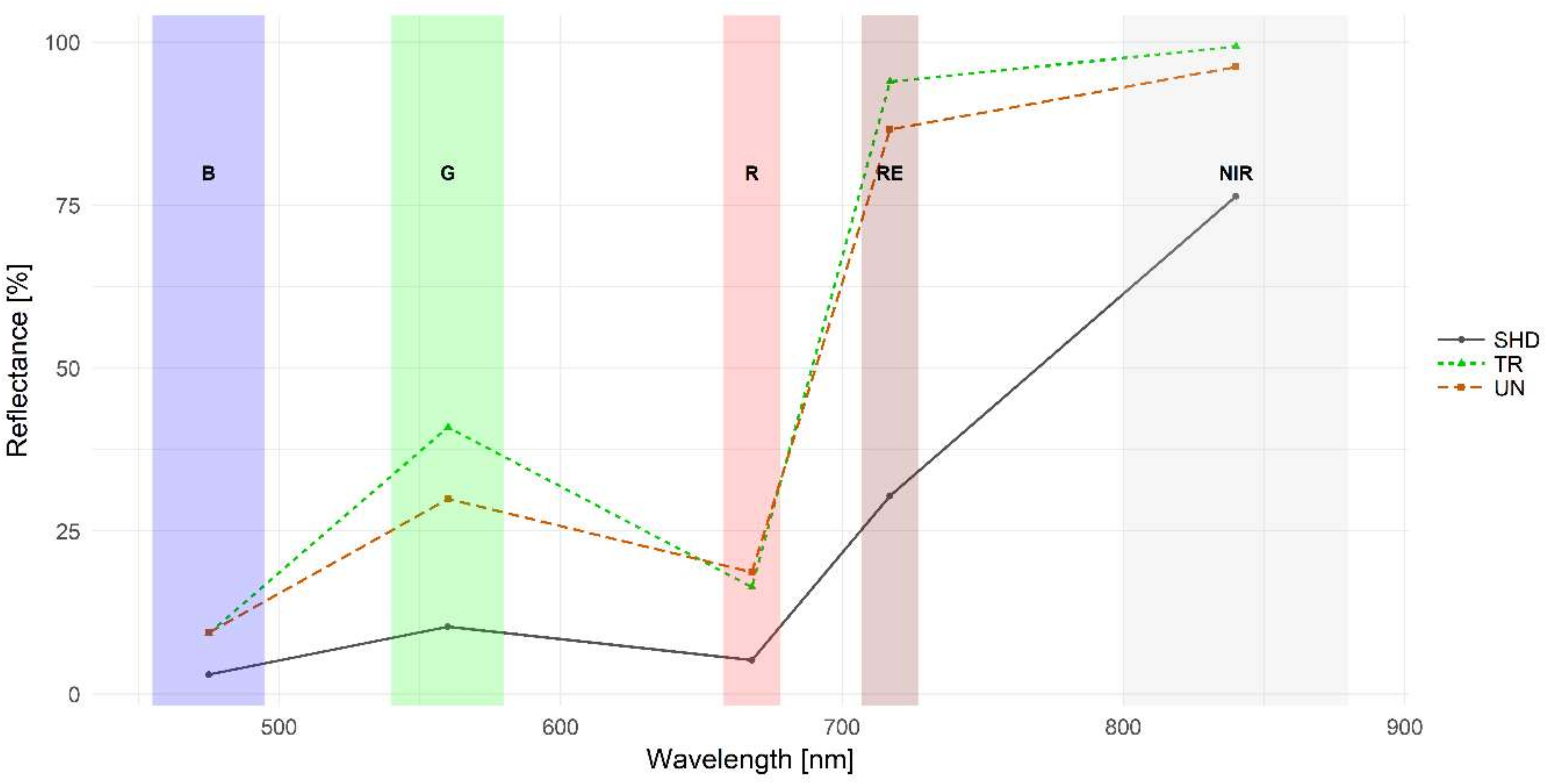
3.1. Multispectral Reflectance Signatures
3.2. Classification
3.3. Important Classification Features
4. Discussion
4.1. Overall Accuracy
4.2. Feature Selection
4.3. Hyperspectral versus Multispectral Sensors for Disease Detection
4.4. Processing Shadow Areas in Land Cover Classification Problems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crist, E.; Mora, C.; Engelman, R. The interaction of human population, food production, and biodiversity protection. Science 2017, 356, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Gebbers, R.; Adamchuk, V.I. Precision Agriculture and Food Security. Science 2010, 327, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kovacs, J.M. The application of small unmanned aerial systems for precision agriculture: A review. Precis. Agric. 2012, 13, 693–712. [Google Scholar] [CrossRef]
- Mulla, D.J. Twenty five years of remote sensing in precision agriculture: Key advances and remaining knowledge gaps. Biosyst. Eng. 2013, 114, 358–371. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Perspectives for Remote Sensing with Unmanned Aerial Vehicles in Precision Agriculture. Trends Plant Sci. 2018, 1–13. [Google Scholar] [CrossRef] [PubMed]
- West, J.S.; Bravo, C.; Oberti, R.; Lemaire, D.; Moshou, D.; McCartney, H.A. The potential of optical canopy measurement for targeted control of field crop diseases. Annu. Rev. Phytopathol. 2003, 41, 593–614. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S. Deadly new wheat disease threatens Europe’s crops. Nature 2017, 542, 145–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahlein, A.-K. Plant Disease Detection by Imaging Sensors – Parallels and Specific Demands for Precision Agriculture and Plant Phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Lechenet, M.; Dessaint, F.; Py, G.; Makowski, D.; Munier-Jolain, N. Reducing pesticide use while preserving crop productivity and profitability on arable farms. Nat. Plants 2017, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.-C.; Mahlein, A.-K.; Steiner, U. Detection and Diagnostics of Plant Pathogens, 5th ed.; Gullino, M.L., Bonants, P.J.M., Eds.; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-017-9019-2. [Google Scholar]
- Dash, J.P.; Watt, M.S.; Pearse, G.D.; Heaphy, M.; Dungey, H.S. Assessing very high resolution UAV imagery for monitoring forest health during a simulated disease outbreak. ISPRS J. Photogramm. Remote Sens. 2017, 131, 1–14. [Google Scholar] [CrossRef]
- Calderón, R.; Montes-Borrego, M.; Landa, B.B.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Detection of downy mildew of opium poppy using high-resolution multi-spectral and thermal imagery acquired with an unmanned aerial vehicle. Precis. Agric. 2014, 15, 639–661. [Google Scholar] [CrossRef] [Green Version]
- Carnegie, A.J.; Lidbetter, J.R.; Walker, J.; Horwood, M.A.; Tesoriero, L.; Glen, M.; Priest, M.J. Uredo rangelii, a taxon in the guava rust complex, newly recorded on Myrtaceae in Australia. Australas. Plant Pathol. 2010, 39, 463–466. [Google Scholar] [CrossRef]
- Glen, M.; Alfenas, A.C.; Zauza, E.A.V.; Wingfield, M.J.; Mohammed, C. Puccinia psidii: A threat to the Australian environment and economy—A review. Australas. Plant Pathol. 2007, 36, 1–16. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Kathuria, A.; Pegg, G.S.; Entwistle, P.; Nagel, M.; Giblin, F.R. Impact of the invasive rust Puccinia psidii (myrtle rust) on native Myrtaceae in natural ecosystems in Australia. Biol. Invasions 2016, 18, 127–144. [Google Scholar] [CrossRef]
- Berthon, K.; Esperon-Rodriguez, M.; Beaumont, L.J.; Carnegie, A.J.; Leishman, M.R. Assessment and prioritisation of plant species at risk from myrtle rust (Austropuccinia psidii) under current and future climates in Australia. Biol. Conserv. 2018, 218, 154–162. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Pegg, G.S. Lessons from the Incursion of Myrtle Rust in Australia. Annu. Rev. Phytopathol. 2018, 56, 457–478. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M. Australian Native Food Industry Stocktake; Rural Industries Research and Development Corporation: Barton, Australia, 2012; ISBN 9781742544090. [Google Scholar]
- Doran, J.; Lea, D.; Bush, D. Assessing Myrtle Rust in a Lemon Myrtle Provenance Trial; Rural Industries Research and Development Corporation: Barton, Australia, 2012; ISBN 9781742544403. [Google Scholar]
- Heim, R.H.J.; Wright, I.J.; Chang, H.-C.; Carnegie, A.J.; Pegg, G.S.; Lancaster, E.K.; Falster, D.S.; Oldeland, J. Detecting myrtle rust (Austropuccinia psidii) on lemon myrtle trees using spectral signatures and machine learning. Plant Pathol. 2018, 67, 1114–1121. [Google Scholar] [CrossRef]
- Bureau of Meteorology Climate Data Online. Available online: http://www.bom.gov.au/climate/data/index.shtml (accessed on 1 August 2018).
- Horwood, M.; Carnegie, A.; Park, R. Gathering Efficacy Data to Indentify the Most Effective Chemicals for Controlling Myrtle Rust (Uredo Rangelii); Plant Health Australia: Canberra, Australia, 2013. [Google Scholar]
- USGS National UAS Project Office. Unmanned Aircraft Systems Data Post-Processing; U.S. Geological Survey (USGS) National Unmanned Aircraft Systems (UAS) Project Office (NUPO): Lakewood, CO, USA, 2017. [Google Scholar]
- Wang, C.; Myint, S.W. A Simplified Empirical Line Method of Radiometric Calibration for Small Unmanned Aircraft Systems-Based Remote Sensing. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 1876–1885. [Google Scholar] [CrossRef]
- QGIS Geographic Information System, Open Source Geospatial Foundation. 2009. Available online: http://qgis.osgeo.org (accessed on 7 March 2019).
- Hijmans, R.J. raster: Geographic Data Analysis and Modeling; 2017. R package version 2.6-7. 2017. Available online: https://CRAN.R-project.org/package=raster (accessed on 7 March 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Davies, K. Plant Pigments and their Manipulation, 1st ed.; Davies, K., Ed.; Blackwell: Oxford, UK, 2004; Volume 14, ISBN 0849323509. [Google Scholar]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical Properties and Nondestructive Estimation of Anthocyanin Content in Plant Leaves. Photochem. Photobiol. 2007, 74, 38–45. [Google Scholar] [CrossRef]
- Lee, D.; Brawner, J.; Pegg, G. Screening Eucalyptus cloeziana and E. argophloia populations for resistance to Puccinia psidii. Plant Dis. 2014, 99, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.R.; McRoberts, N. Plants and biotrophs: A pivotal role for cytokinins? Trends Plant Sci. 2006, 11, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Ramsfield, T.; Dick, M.; Bulman, L.; Ganley, R. Briefing Document on Myrtle Rust, a Member of the Guava Rust Complex, and the Risk To New Zealand; SCION Next generation biomaterials: Rotorua, New Zealand, 2010. [Google Scholar]
- Wang, E.; Dong, C.; Park, R.F.; Roberts, T.H. Carotenoid pigments in rust fungi: Extraction, separation, quantification and characterisation. Fungal Biol. Rev. 2018, 32, 166–180. [Google Scholar] [CrossRef]
- Penuelas, J.; Baret, F.; Filella, I. Semi-empirical indices to assess carotenoids/chlorophyll a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Morin, L.; Talbot, M.J.; Glen, M. Quest to elucidate the life cycle of Puccinia psidii sensu lato. Fungal Biol. 2014, 118, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W.; Hass, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the great plains with ERTS. In Proceedings of the Third Earth Resources Technology Satellite Symposium, Washington, DC, USA, 10–14 December 1973; Volume 1, pp. 309–317. [Google Scholar]
- Di Gennaro, S.F.; Battiston, E.; Di Marco, S.; Facini, O.; Matese, A. Unmanned Aerial Vehicle (UAV)-based remote sensing to monitor grapevine leaf stripe disease within a vineyard affected by esca complex. Phytopathol. Mediterr. 2016, 55, 262–275. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning, 2nd ed.; Springer Series in Statistics; Springer: New York, NY, USA, 2009; ISBN 978-0-387-84857-0. [Google Scholar]
- Belgiu, M.; Drăgu, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.-M.; Tuleau-Malot, C. VSURF: An R Package for Variable Selection Using Random Forests. R J. 2015, 7, 19–33. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.; Tuleau-Malot, C. Variable selection using random forests. Pattern Recognit. Lett. 2010, 31, 2225–2236. [Google Scholar] [CrossRef] [Green Version]
- Story, M.; Congalton, R.G. Accuracy assessment: A user’s perspective. Photogramm. Eng. Remote Sens. 1986, 52, 397–399. [Google Scholar] [CrossRef]
- Government New Zealand Protection and Response: Myrtle Rust. Available online: https://www.mpi.govt.nz/protection-and-response/responding/alerts/myrtle-rust (accessed on 10 September 2018).
- Lambert, S.; Waipara, N.; Black, A.; Mark-Shadbolt, M.; Wood, W. Indigenous Biosecurity: Māori Responses to Kauri Dieback and Myrtle Rust in Aotearoa New Zealand. In The Human Dimensions of Forest and Tree Health; Springer International Publishing: Cham, Switzerland, 2018; pp. 109–137. [Google Scholar] [Green Version]
- Sandino, J.; Pegg, G.; Gonzalez, F.; Smith, G. Aerial Mapping of Forests Affected by Pathogens Using UAVs, Hyperspectral Sensors, and Artificial Intelligence. Sensors 2018, 18, 944. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.-K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral Sensors and Imaging Technologies in Phytopathology: State of the Art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.K.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Spectral signatures of sugar beet leaves for the detection and differentiation of diseases. Precis. Agric. 2010, 11, 413–431. [Google Scholar] [CrossRef]
- Albetis, J.; Duthoit, S.; Guttler, F.; Jacquin, A.; Goulard, M.; Poilvé, H.; Féret, J.B.; Dedieu, G. Detection of Flavescence dorée grapevine disease using Unmanned Aerial Vehicle (UAV) multispectral imagery. Remote Sens. 2017, 9, 1–20. [Google Scholar] [CrossRef]
- Gates, D.M.; Keegan, H.J.; Schleter, J.C.; Weidner, V.R. Spectral Properties of Plants. Appl. Opt. 1965, 4, 11. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N.; Lichtenthaler, H.K. Detection of Red Edge Position and Chlorophyll Content by Reflectance Measurements Near 700 nm. J. Plant Physiol. 1996, 148, 501–508. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and Physiological Basis for the Reflectance of Visible and Near Infrared Radiation from Vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Skidmore, A.K. Plant phenolics and absorption features in vegetation reflectance spectra near 1.66 μm. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 55–83. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Kattenborn, T.; Fassnacht, F.E. How canopy shadow affects invasive plant species classification in high spatial resolution remote sensing. Remote Sens. Ecol. Conserv. 2019, 1–16. [Google Scholar] [CrossRef]
- Mostafa, Y. A Review on Various Shadow Detection and Compensation Techniques in Remote Sensing Images. Can. J. Remote Sens. 2017, 43, 545–562. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectral Vegetation Index (SVI) | SVI Abbreviation | Formula | Reference |
|---|---|---|---|
| Normalized Difference Vegetation Index | NDVI | [37] | |
| Structure Insensitive Pigment Index | SIPI | [35] | |
| Anthocyanin Reflectance Index | ARI | [30] | |
| Green/Red Simple Ratio Index | G/R | [13] |
| Error Matrix | Reference | |||||
|---|---|---|---|---|---|---|
| Shadow | Treated | Untreated | Total | UA | ||
| Prediction | Shadow | 1158 | 19 | 24 | 1201 | 96.4% |
| Treated | 14 | 1128 | 37 | 1179 | 95.7% | |
| Untreated | 30 | 56 | 1142 | 1228 | 93.0% | |
| Total | 1202 | 1203 | 1203 | 3608 | ||
| PA | 96.3% | 93.8% | 94.9% | 95.0% | ||
| Rank | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Band | RE | NIR | R | G/R | ARI | G | NDVI | B | SIPI |
| Abs. Imp. | 0.3 | 0.26 | 0.13 | 0.13 | 0.11 | 0.09 | 0.09 | 0.07 | 0.04 |
| Rel. Imp. | 1 | 0.85 | 0.36 | 0.34 | 0.26 | 0.21 | 0.17 | 0.13 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heim, R.H.J.; Wright, I.J.; Scarth, P.; Carnegie, A.J.; Taylor, D.; Oldeland, J. Multispectral, Aerial Disease Detection for Myrtle Rust (Austropuccinia psidii) on a Lemon Myrtle Plantation. Drones 2019, 3, 25. https://doi.org/10.3390/drones3010025
Heim RHJ, Wright IJ, Scarth P, Carnegie AJ, Taylor D, Oldeland J. Multispectral, Aerial Disease Detection for Myrtle Rust (Austropuccinia psidii) on a Lemon Myrtle Plantation. Drones. 2019; 3(1):25. https://doi.org/10.3390/drones3010025
Chicago/Turabian StyleHeim, René H.J., Ian J. Wright, Peter Scarth, Angus J. Carnegie, Dominique Taylor, and Jens Oldeland. 2019. "Multispectral, Aerial Disease Detection for Myrtle Rust (Austropuccinia psidii) on a Lemon Myrtle Plantation" Drones 3, no. 1: 25. https://doi.org/10.3390/drones3010025
APA StyleHeim, R. H. J., Wright, I. J., Scarth, P., Carnegie, A. J., Taylor, D., & Oldeland, J. (2019). Multispectral, Aerial Disease Detection for Myrtle Rust (Austropuccinia psidii) on a Lemon Myrtle Plantation. Drones, 3(1), 25. https://doi.org/10.3390/drones3010025