
Unfolded Lipase at Interfaces Studied via Interfacial Dilational Rheology: The Impact of Urea

 ,
,  ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
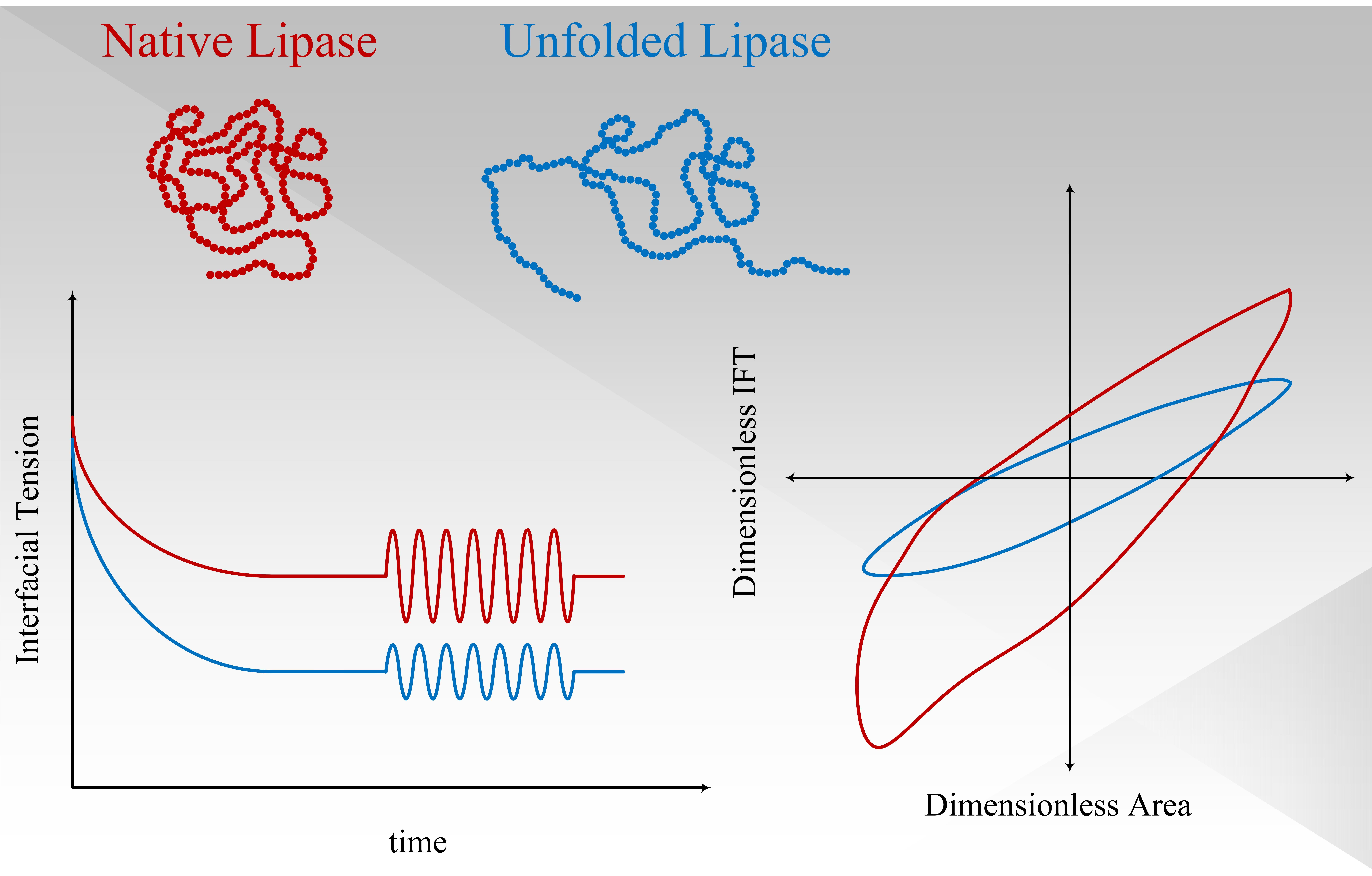
1. Introduction
2. Materials and Methods
2.1. Materials
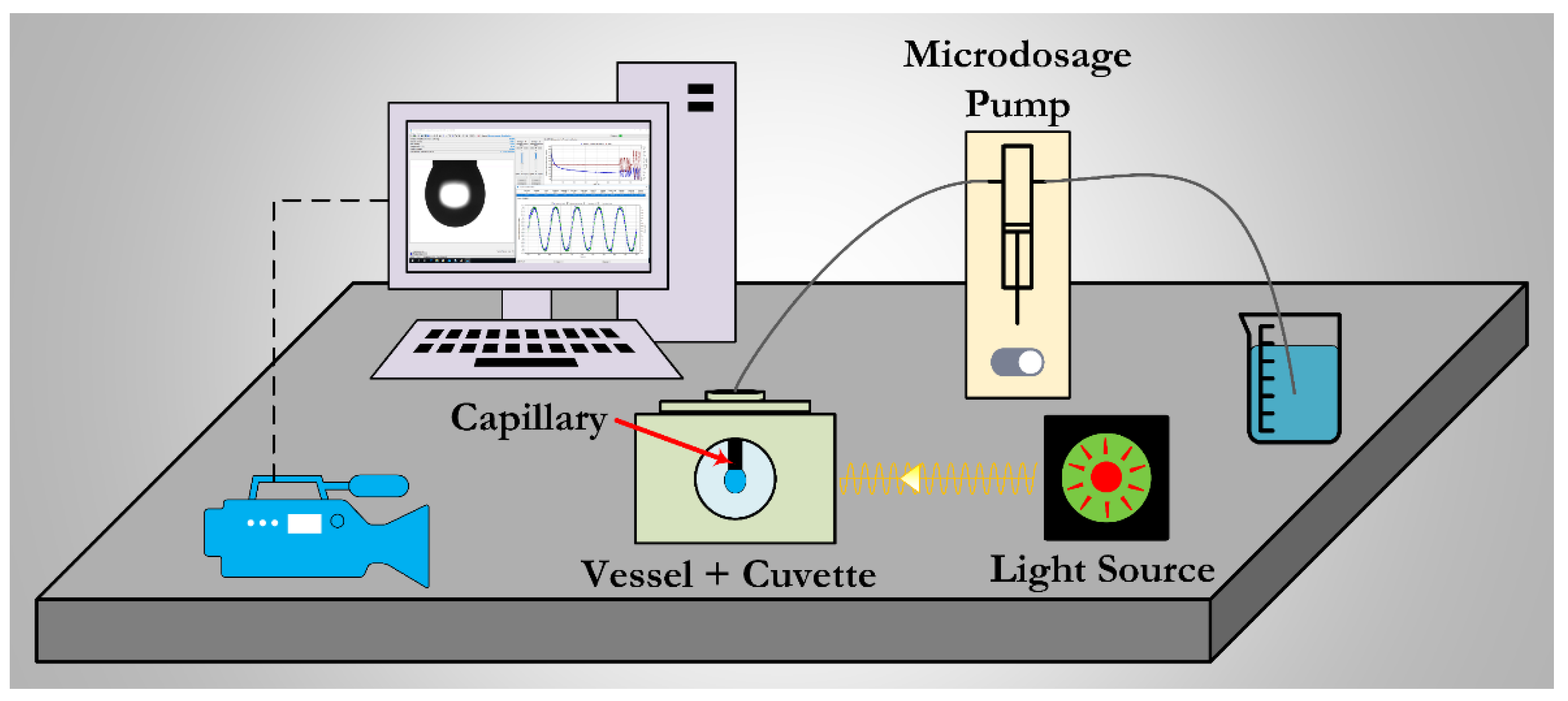
2.2. Interfacial Dilational Rheology
3. Results and Discussion
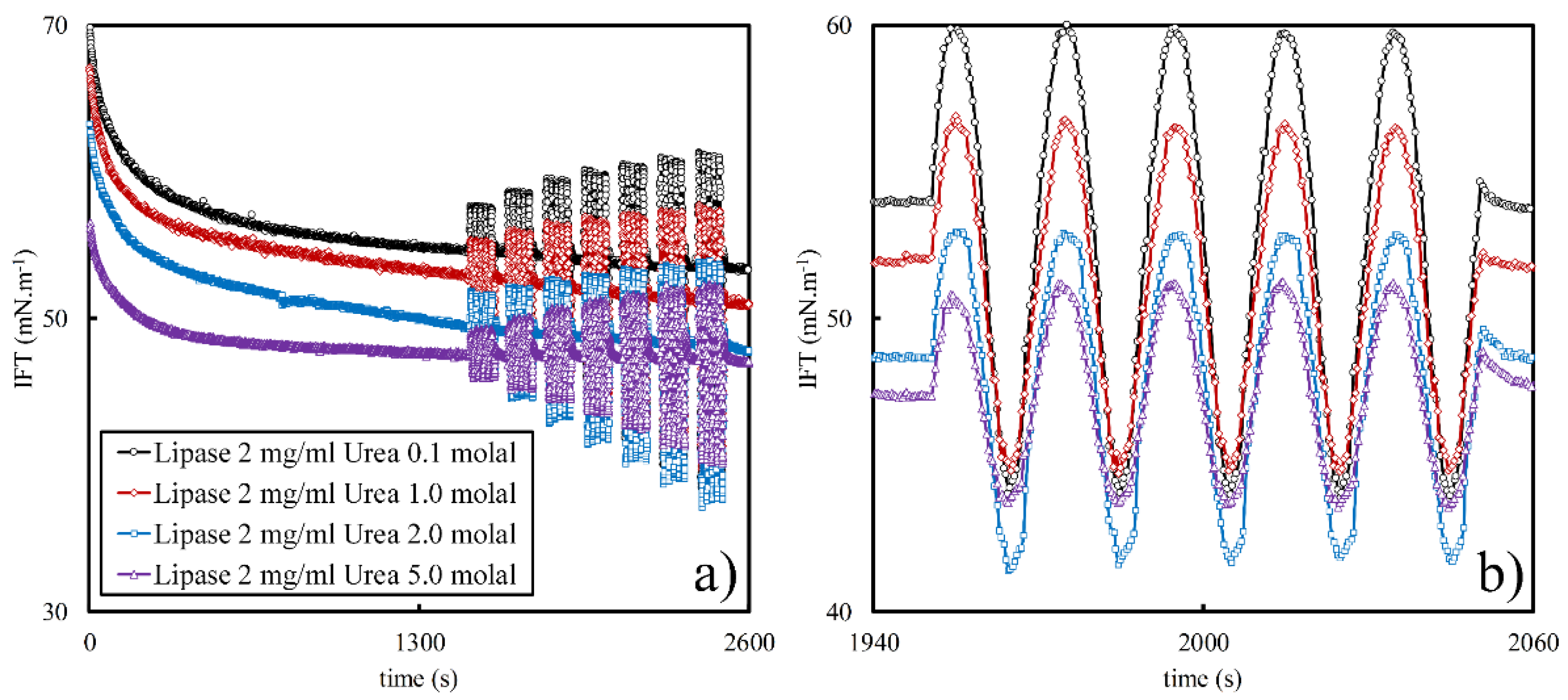
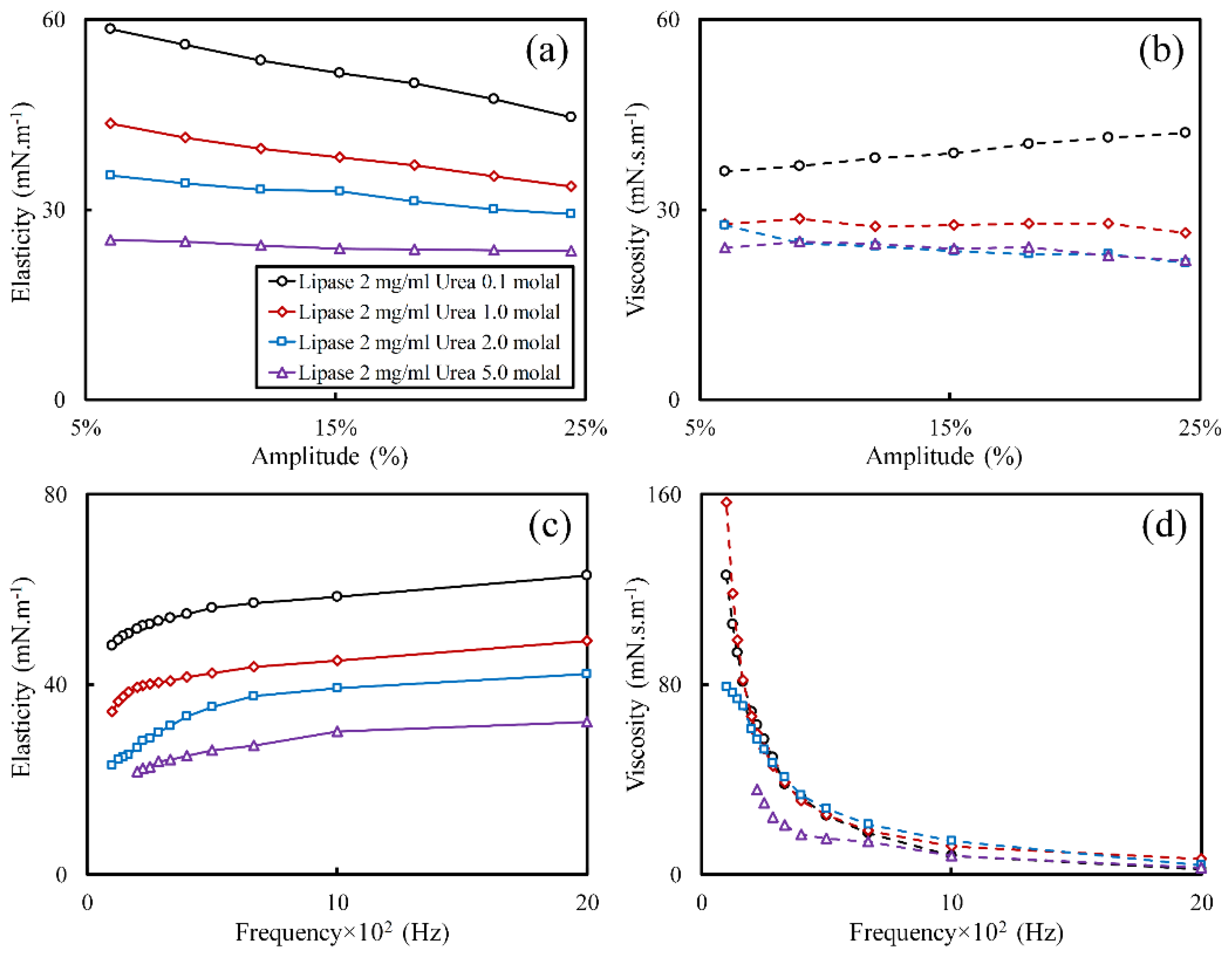
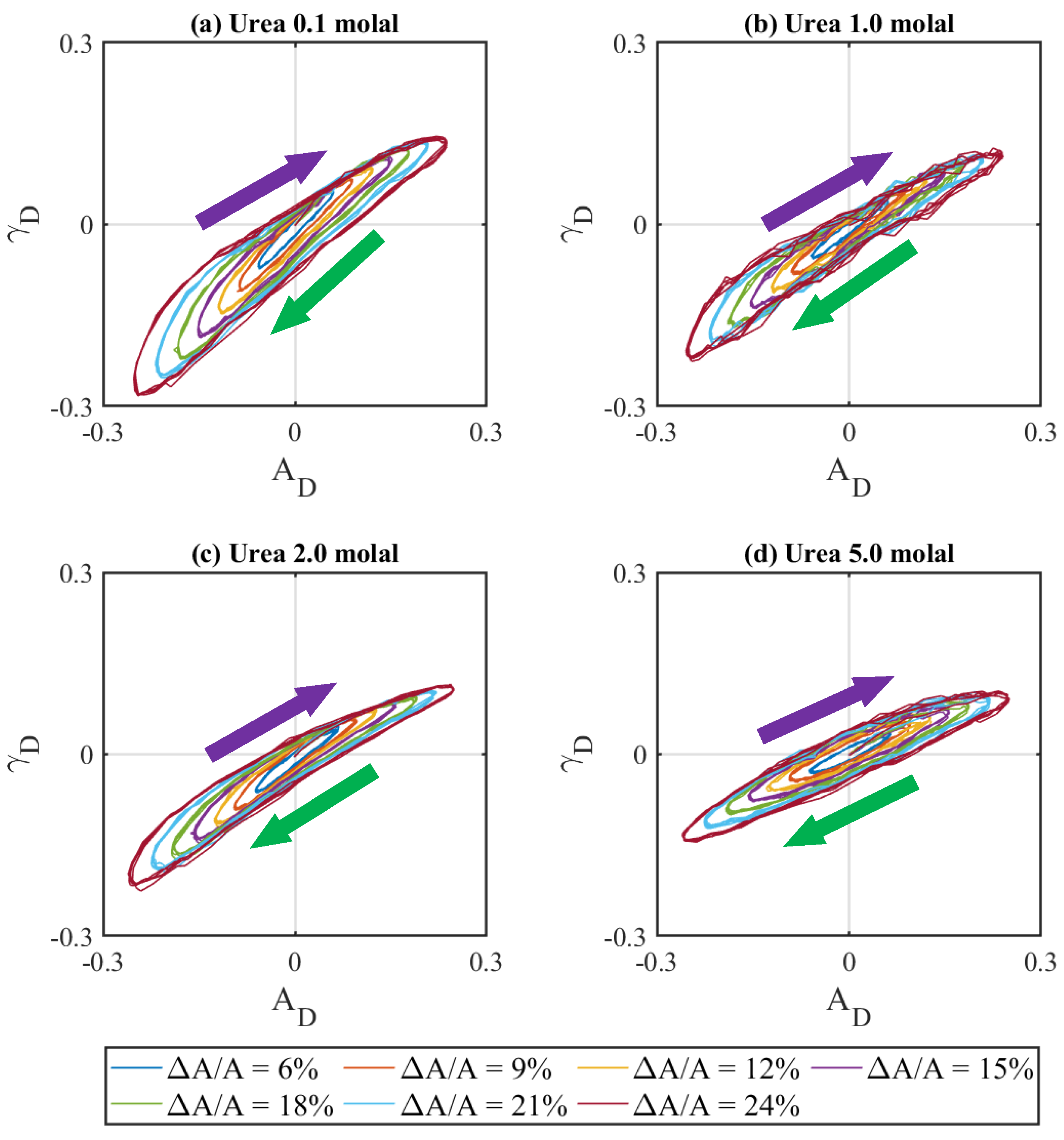
3.1. Effect of Urea Concentration on Viscoelasticity of Lipase Adsorbed Layers
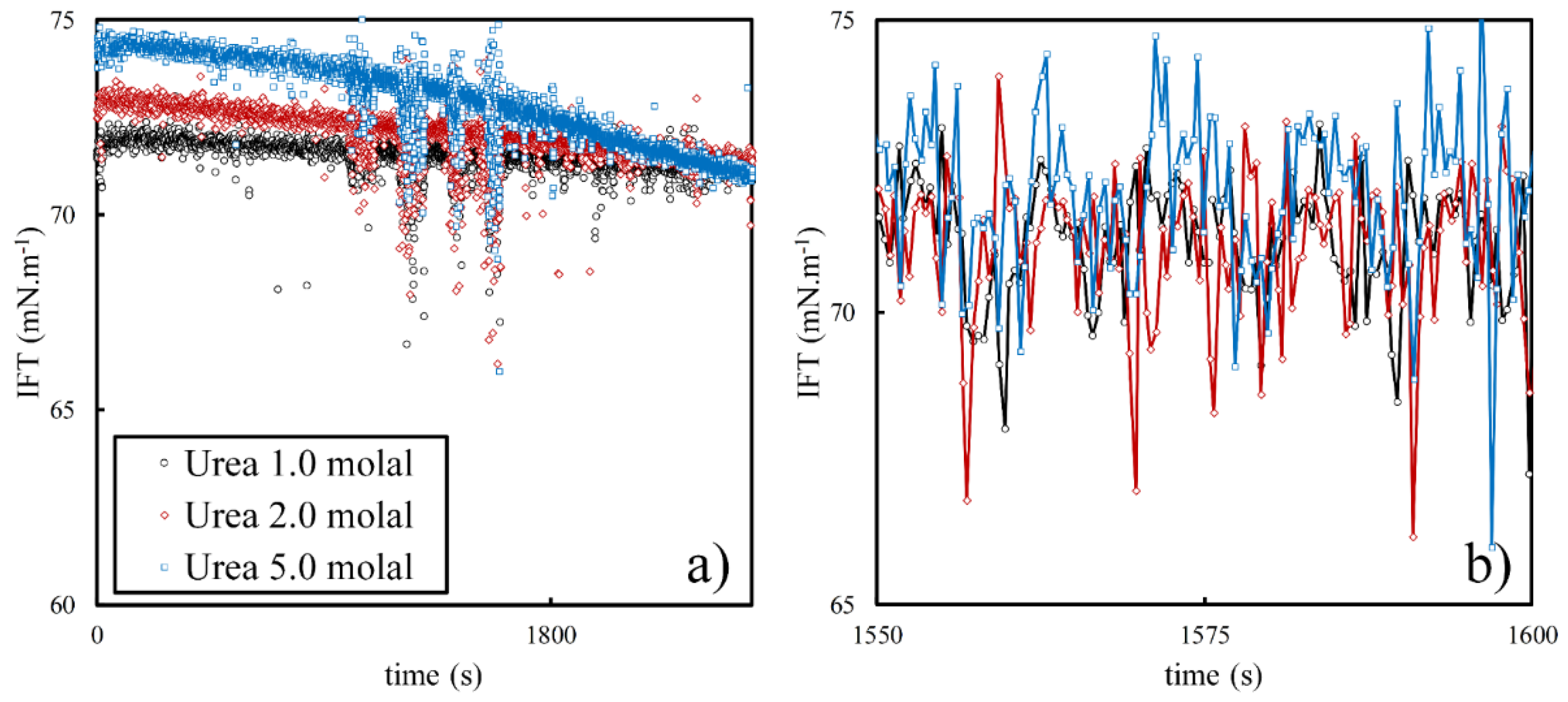
3.2. Effect of Urea on Viscoelasticity of Droplet Surfaces
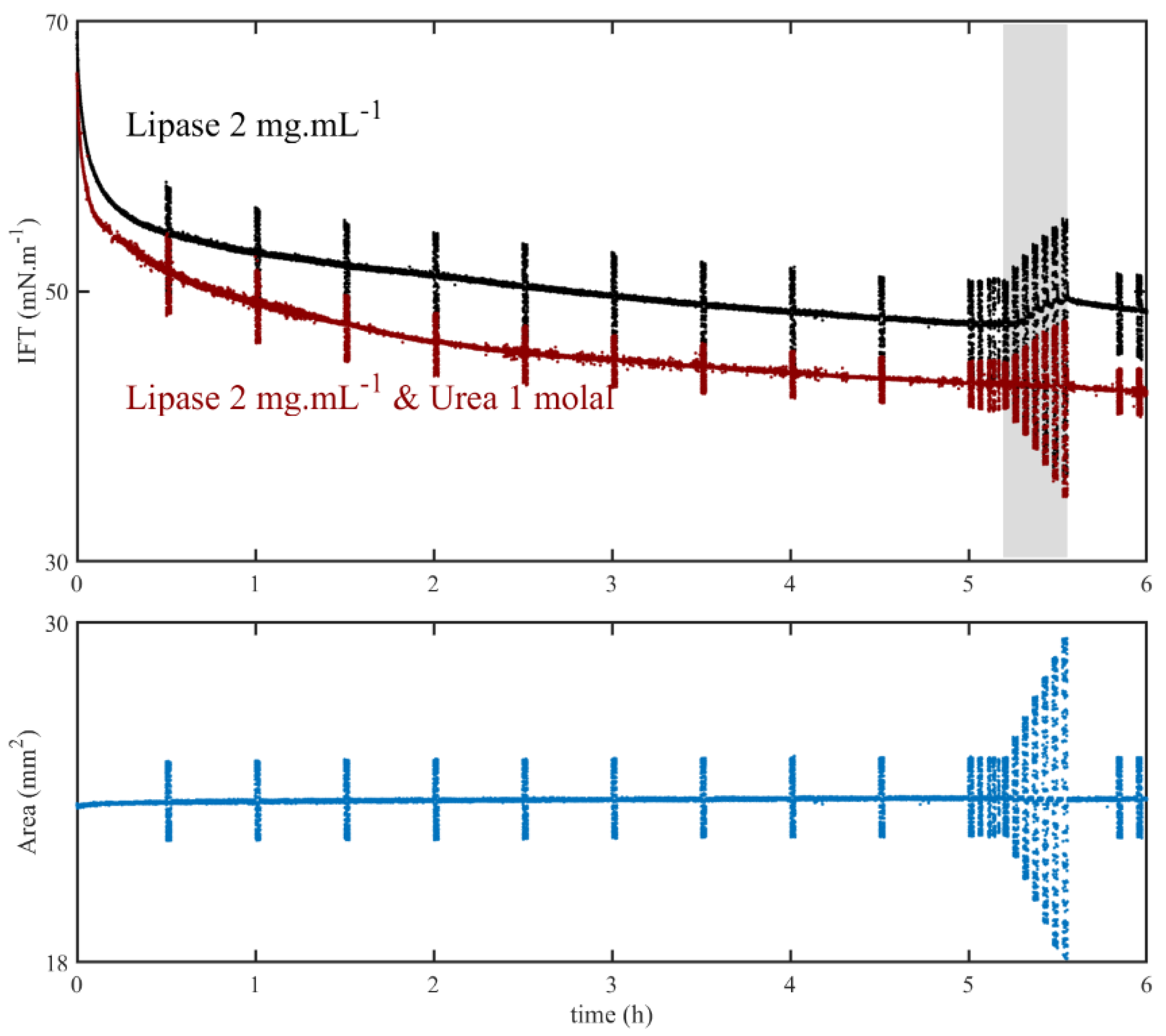
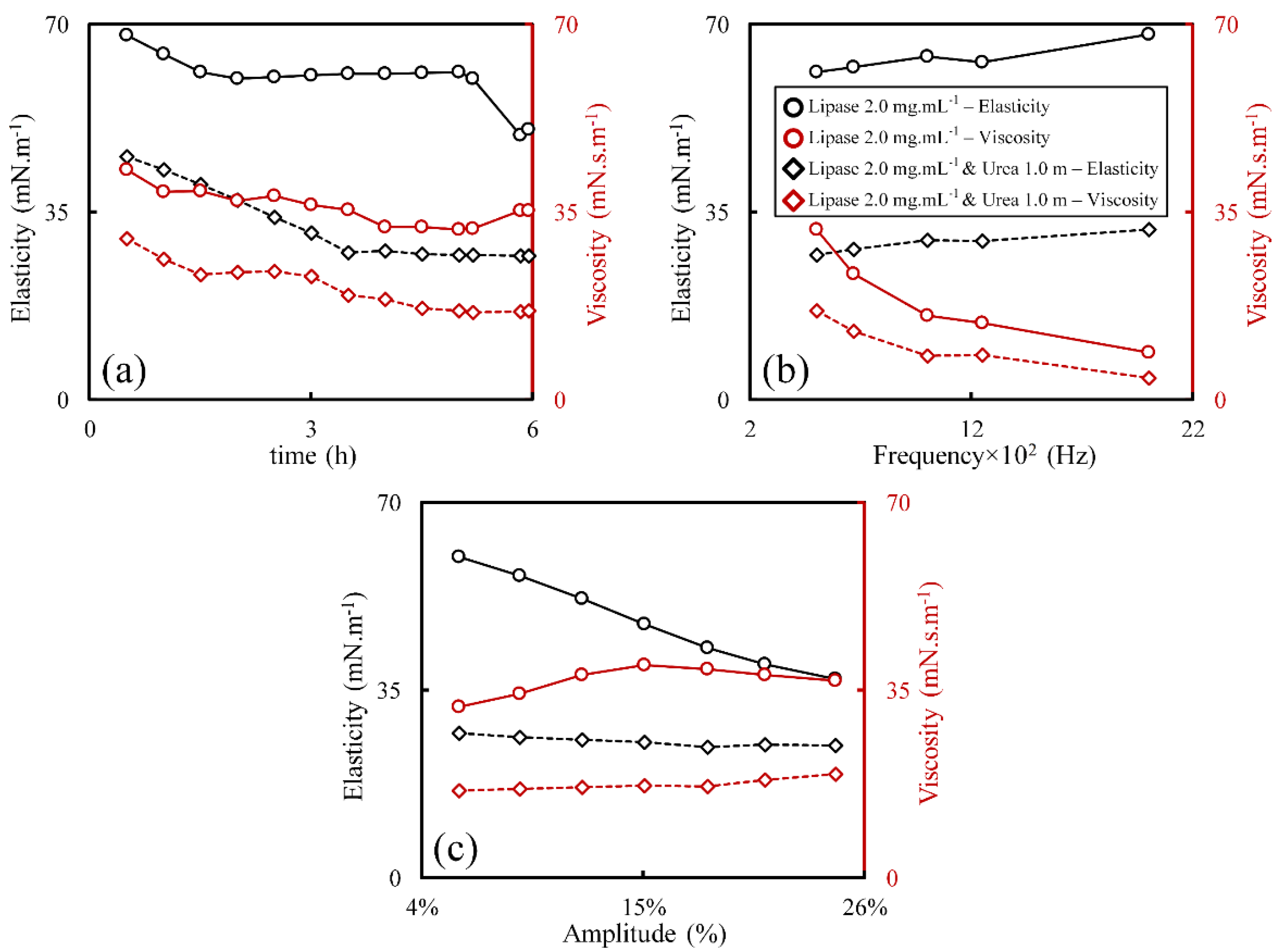
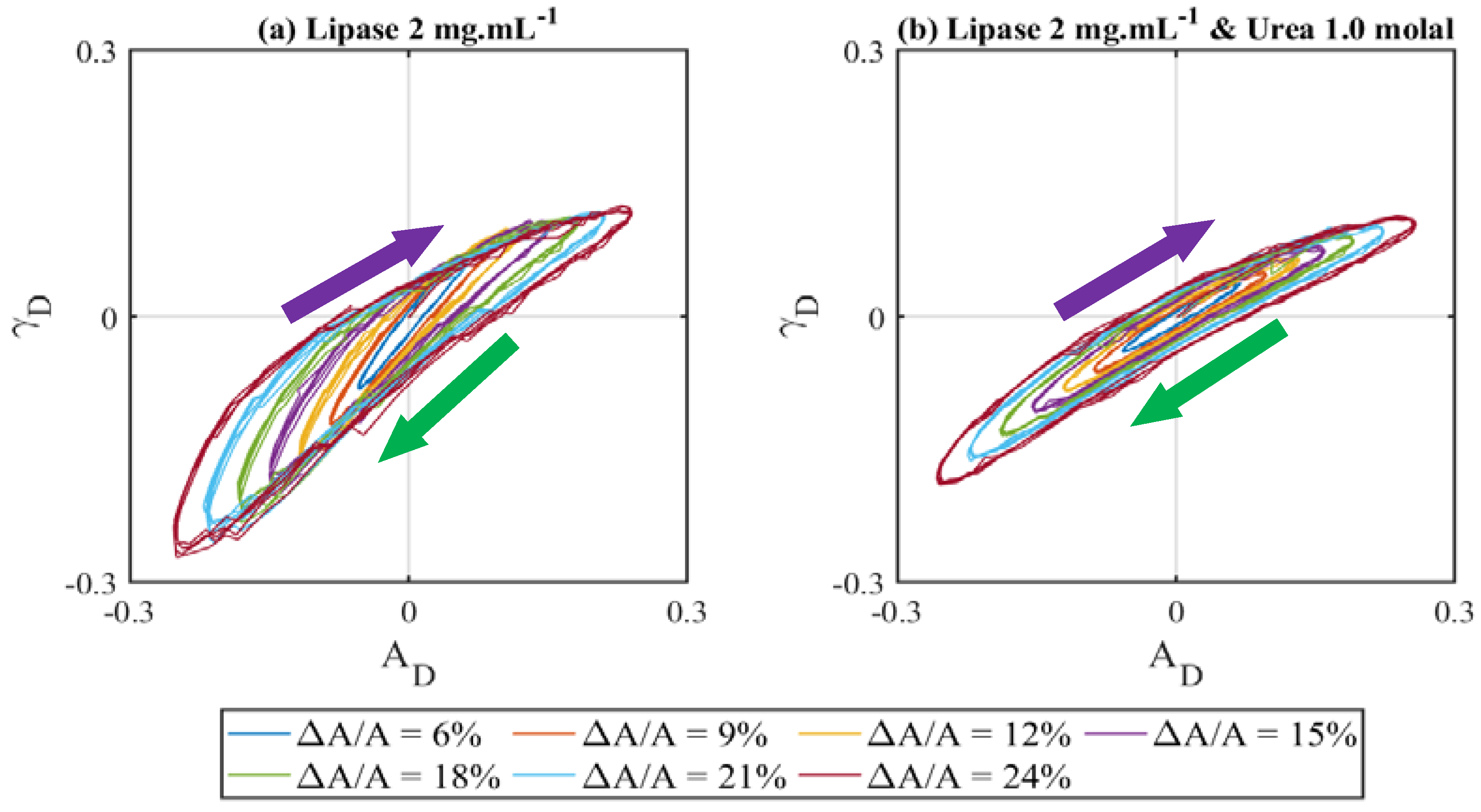
3.3. Effect of Interface Age
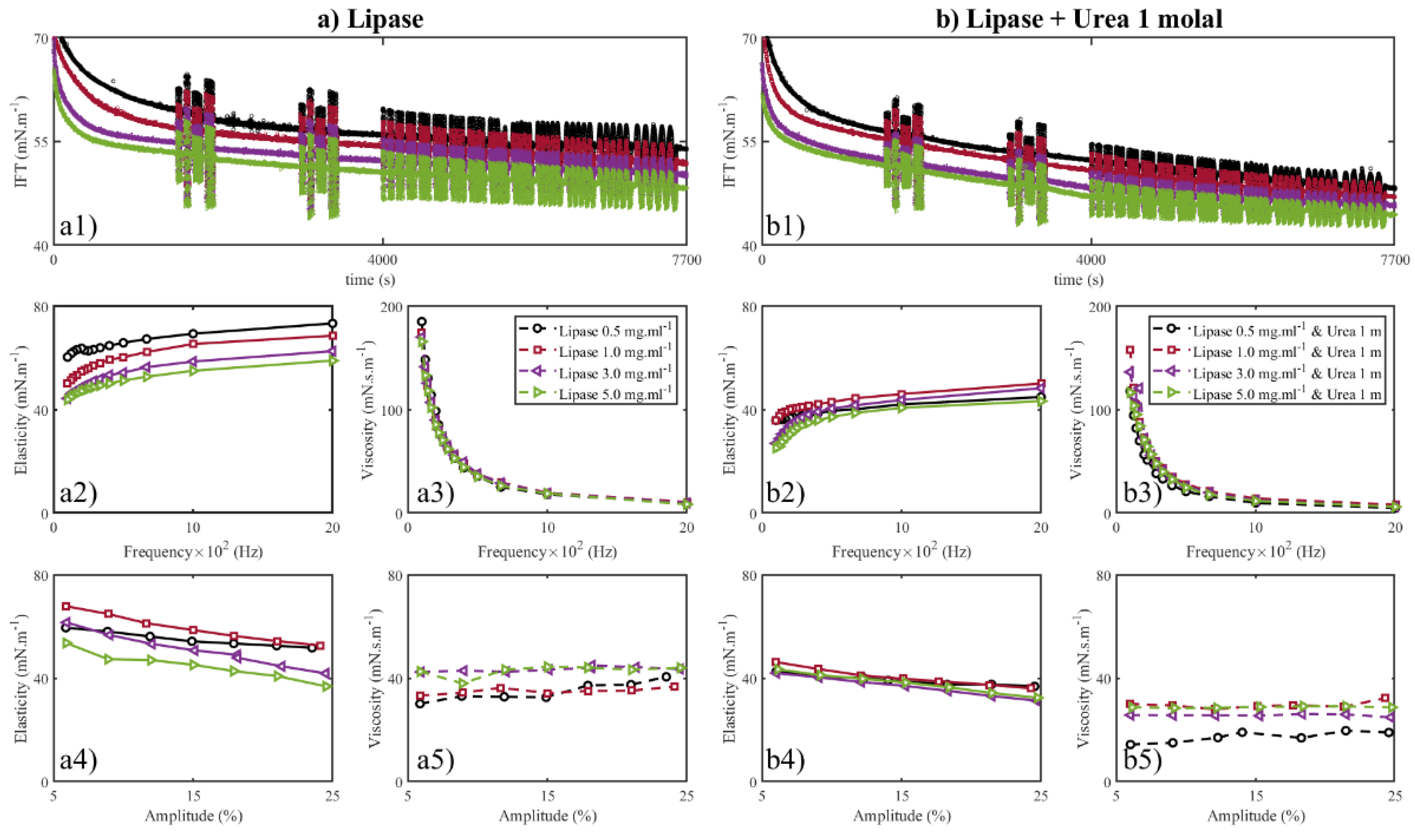
3.4. Effect of Lipase Concentration
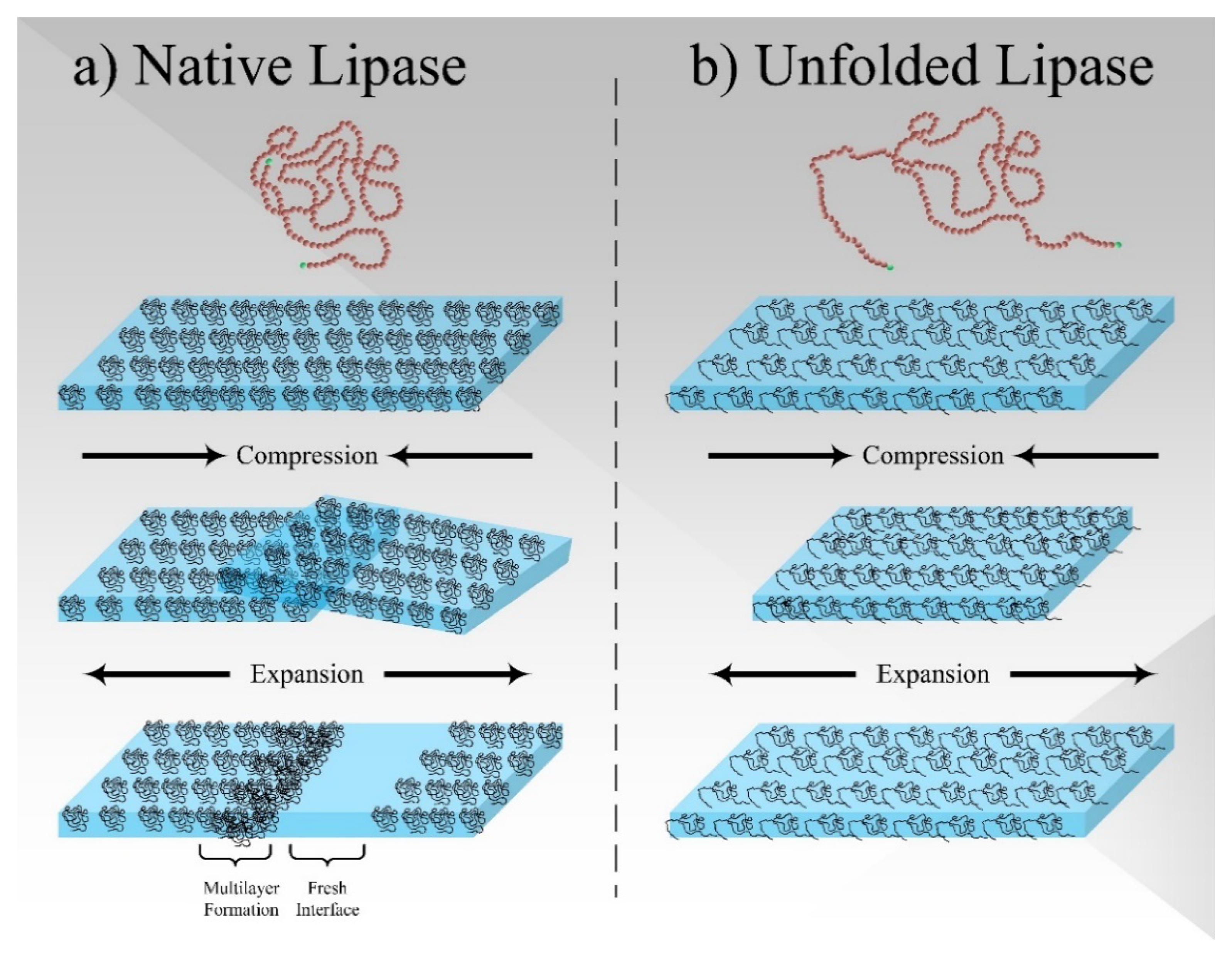
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Svendsen, A. Lipase protein engineering. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2000, 1543, 223–238. [Google Scholar] [CrossRef]
- Freedman, B.; Butterfield, R.O.; Pryde, E.H. Transesterification kinetics of soybean oil 1. J. Am. Oil Chem. Soc. 1986, 63, 1375–1380. [Google Scholar] [CrossRef]
- Reis, P.; Holmberg, K.; Watzke, H.; Leser, M.E.; Miller, R. Lipases at interfaces: A review. Adv. Colloid Interface Sci. 2009, 147, 237–250. [Google Scholar] [CrossRef]
- Rader, A.J.; Hespenheide, B.M.; Kuhn, L.A.; Thorpe, M.F. Protein unfolding: Rigidity lost. Proc. Natl. Acad. Sci. USA 2002, 99, 3540–3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocco, A.G.; Mollica, L.; Ricchiuto, P.; Baptista, A.M.; Gianazza, E.; Eberini, I. Characterization of the Protein Unfolding Processes Induced by Urea and Temperature. Biophys. J. 2008, 94, 2241–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busti, P.A.; Scarpeci, S.; Gatti, C.; Delorenzi, N. Use of fluorescence methods to monitor unfolding transitions in β-lactoglobulin. Food Res. Int. 2002, 35, 871–877. [Google Scholar] [CrossRef]
- Czarnik-Matusewicz, B.; Bin Kim, S.; Jung, Y.M. A Study of Urea-dependent Denaturation of β-Lactoglobulin by Principal Component Analysis and Two-dimensional Correlation Spectroscopy. J. Phys. Chem. B 2008, 113, 559–566. [Google Scholar] [CrossRef]
- D’Alfonso, L.; Collini, M.; Baldini, G. Does β-Lactoglobulin Denaturation Occur via an Intermediate State? Biochemistry 2002, 41, 326–333. [Google Scholar] [CrossRef]
- Mikhailovskaya, A.; Noskov, B.; Nikitin, E.; Lin, S.-Y.; Loglio, G.; Miller, R. Dilational surface viscoelasticity of protein solutions. Impact of urea. Food Hydrocoll. 2014, 34, 98–103. [Google Scholar] [CrossRef]
- Tihonov, M.M.; Milyaeva, O.Y.; Noskov, B.A. Dynamic surface properties of lysozyme solutions. Impact of urea and guanidine hydrochloride. Colloids Surf. B Biointerfaces 2015, 129, 114–120. [Google Scholar] [CrossRef]
- Das, A.; Chitra, R.; Choudhury, R.R.; Ramanadham, M. Structural changes during the unfolding of Bovine serum albumin in the presence of urea: A small-angle neutron scattering study. Pramana 2004, 63, 363–368. [Google Scholar] [CrossRef]
- Leggio, C.; Galantini, L.; Konarev, P.V.; Pavel, N.V. Urea-Induced Denaturation Process on Defatted Human Serum Albumin and in the Presence of Palmitic Acid. J. Phys. Chem. B 2009, 113, 12590–12602. [Google Scholar] [CrossRef] [PubMed]
- Chaitanya, P.K.; Prabhu, N.P. Stability and Activity of Porcine Lipase Against Temperature and Chemical Denaturants. Appl. Biochem. Biotechnol. 2014, 174, 2711–2724. [Google Scholar] [CrossRef] [PubMed]
- Rashno, F.; Khajeh, K.; Capitini, C.; Sajedi, R.H.; Shokri, M.M.; Chiti, F. Very rapid amyloid fibril formation by a bacterial lipase in the absence of a detectable lag phase. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2017, 1865, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Qafary, M.; Khajeh, K.; Ramazzotti, M.; Moosavi-Movahedi, A.A.; Chiti, F. Urea titration of a lipase from Pseudomonas sp. reveals four different conformational states, with a stable partially folded state explaining its high aggregation propensity. Int. J. Biol. Macromol. 2021, 174, 32–41. [Google Scholar] [CrossRef]
- Eberini, I.; Emerson, A.; Sensi, C.; Ragona, L.; Ricchiuto, P.; Pedretti, A.; Gianazza, E.; Tramontano, A. Simulation of urea-induced protein unfolding: A lesson from bovine β-lactoglobulin. J. Mol. Graph. Model. 2011, 30, 24–30. [Google Scholar] [CrossRef]
- Hédoux, A.; Krenzlin, S.; Paccou, L.; Guinet, Y.; Flament, M.-P.; Siepmann, J. Influence of urea and guanidine hydrochloride on lysozyme stability and thermal denaturation; a correlation between activity, protein dynamics and conformational changes. Phys. Chem. Chem. Phys. 2010, 12, 13189–13196. [Google Scholar] [CrossRef]
- Kuwajima, K. The molten globule state as a clue for understanding the folding and cooperativity of globular-protein structure. Proteins Struct. Funct. Bioinform. 1989, 6, 87–103. [Google Scholar] [CrossRef]
- Maldonado-Valderrama, J.; Martín-Rodriguez, A.; Gálvez-Ruiz, M.J.; Miller, R.; Langevin, D.; Cabrerizo-Vílchez, M.A. Foams and emulsions of β-casein examined by interfacial rheology. Colloids Surf. A Physicochem. Eng. Asp. 2008, 323, 116–122. [Google Scholar] [CrossRef]
- Noskov, B.A. Protein conformational transitions at the liquid–gas interface as studied by dilational surface rheology. Adv. Colloid Interface Sci. 2014, 206, 222–238. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, Y.; Li, Y.; Liu, Y.; Wang, C.; Xu, B.; Zhao, L.; Xu, B. Interfacial rheological properties of cholesteryl-oligopeptide surfactants: Effects of hydrophilic group structure. J. Mol. Liq. 2022, 365, 120198. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, P.; Chen, Z.; Liu, Y.; Zhao, L.; Wang, N.; Xu, B. Effects of fatty acyl chains on the interfacial rheological behaviors of amino acid surfactants. J. Mol. Liq. 2021, 325, 114823. [Google Scholar] [CrossRef]
- Reis, P.; Malmsten, M.; Nydén, M.; Folmer, B.; Holmberg, K. Interactions between lipases and amphiphiles at interfaces. J. Surfactants Deterg. 2019, 22, 1047–1058. [Google Scholar] [CrossRef]
- Javadi, A.; Dowlati, S.; Miller, R.; Schneck, E.; Eckert, K.; Kraume, M. Dynamics of Competitive Adsorption of Lipase and Ionic Surfactants at the Water–Air Interface. Langmuir 2020, 36, 12010–12022. [Google Scholar] [CrossRef]
- Mekkaoui, A.; Liu, Y.; Zhang, P.; Ullah, S.; Wang, C.; Xu, B. Effect of Bile Salts on the Interfacial Dilational Rheology of Lecithin in the Lipid Digestion Process. J. Oleo Sci. 2021, 70, 1069–1080. [Google Scholar] [CrossRef]
- Noskov, B.A.; Mikhailovskaya, A.A.; Lin, S.-Y.; Loglio, G.; Miller, R. Bovine Serum Albumin Unfolding at the Air/Water Interface as Studied by Dilational Surface Rheology. Langmuir 2010, 26, 17225–17231. [Google Scholar] [CrossRef]
- Javadi, A.; Dowlati, S.; Shourni, S.; Rusli, S.; Eckert, K.; Miller, R.; Kraume, M. Enzymatic Hydrolysis of Triglycerides at the Water–Oil Interface Studied via Interfacial Rheology Analysis of Lipase Adsorption Layers. Langmuir 2021, 37, 12919–12928. [Google Scholar] [CrossRef]
- Hoorfar, M.; Neumann, A.W. Recent progress in Axisymmetric Drop Shape Analysis (ADSA). Adv. Colloid Interface Sci. 2006, 121, 25–49. [Google Scholar] [CrossRef]
- Loglio, G.; Pandolfini, P.; Miller, R.; Makievski, A.V.; Krägel, J.; Ravera, F.; Noskov, B.A. Perturbation–response relationship in liquid interfacial systems: Non-linearity assessment by frequency–domain analysis. Colloids Surfaces A Physicochem. Eng. Asp. 2005, 261, 57–63. [Google Scholar] [CrossRef]
- Loglio, G.; Tesei, U.; Cini, R. Dilational properties of monolayers at the oil-air interface. J. Colloid Interface Sci. 1984, 100, 393–396. [Google Scholar] [CrossRef]
- Loglio, G.; Tesei, U.; Cini, R. Viscoelastic dilatation processes of fluid/fluid interfaces: Time-domain representation. Colloid Polym. Sci. 1986, 264, 712–718. [Google Scholar] [CrossRef]
- Campbell, R.A.; Tummino, A.; Varga, I.; Milyaeva, O.Y.; Krycki, M.M.; Lin, S.-Y.; Laux, V.; Haertlein, M.; Forsyth, V.T.; Noskov, B.A. Adsorption of Denaturated Lysozyme at the Air–Water Interface: Structure and Morphology. Langmuir 2018, 34, 5020–5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bykov, A.; Liggieri, L.; Noskov, B.; Pandolfini, P.; Ravera, F.; Loglio, G. Surface dilational rheological properties in the nonlinear domain. Adv. Colloid Interface Sci. 2015, 222, 110–118. [Google Scholar] [CrossRef]
- Sears, D.F. Surface activity of urea. J. Colloid Interface Sci. 1969, 29, 288–295. [Google Scholar] [CrossRef]
- Rusli, S.; Grabowski, J.; Drews, A.; Kraume, M. A Multi-Scale Approach to Modeling the Interfacial Reaction Kinetics of Lipases with Emphasis on Enzyme Adsorption at Water-Oil Interfaces. Processes 2020, 8, 1082. [Google Scholar] [CrossRef]
- Bykov, A.G.; Noskov, B.A.; Loglio, G.; Lyadinskaya, V.V.; Miller, R. Dilational surface elasticity of spread monolayers of polystyrene microparticles. Soft Matter 2014, 10, 6499–6505. [Google Scholar] [CrossRef]
- Yazhgur, P.A.; Noskov, B.A.; Liggieri, L.; Lin, S.-Y.; Loglio, G.; Miller, R.; Ravera, F. Dynamic properties of mixed nanoparticle/surfactant adsorption layers. Soft Matter 2013, 9, 3305–3314. [Google Scholar] [CrossRef]
- Hinderink, E.B.; Meinders, M.B.; Miller, R.; Sagis, L.; Schroën, K.; Berton-Carabin, C.C. Interfacial protein-protein displacement at fluid interfaces. Adv. Colloid Interface Sci. 2022, 305, 102691. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dowlati, S.; Javadi, A.; Miller, R.; Eckert, K.; Kraume, M. Unfolded Lipase at Interfaces Studied via Interfacial Dilational Rheology: The Impact of Urea. Colloids Interfaces 2022, 6, 56. https://doi.org/10.3390/colloids6040056
Dowlati S, Javadi A, Miller R, Eckert K, Kraume M. Unfolded Lipase at Interfaces Studied via Interfacial Dilational Rheology: The Impact of Urea. Colloids and Interfaces. 2022; 6(4):56. https://doi.org/10.3390/colloids6040056
Chicago/Turabian StyleDowlati, Saeid, Aliyar Javadi, Reinhard Miller, Kerstin Eckert, and Matthias Kraume. 2022. "Unfolded Lipase at Interfaces Studied via Interfacial Dilational Rheology: The Impact of Urea" Colloids and Interfaces 6, no. 4: 56. https://doi.org/10.3390/colloids6040056
APA StyleDowlati, S., Javadi, A., Miller, R., Eckert, K., & Kraume, M. (2022). Unfolded Lipase at Interfaces Studied via Interfacial Dilational Rheology: The Impact of Urea. Colloids and Interfaces, 6(4), 56. https://doi.org/10.3390/colloids6040056