
ESR Dating Ungulate Teeth and Molluscs from the Paleolithic Site Marathousa 1, Megalopolis Basin, Greece

 , ,
, ,
Abstract
:1. Introduction
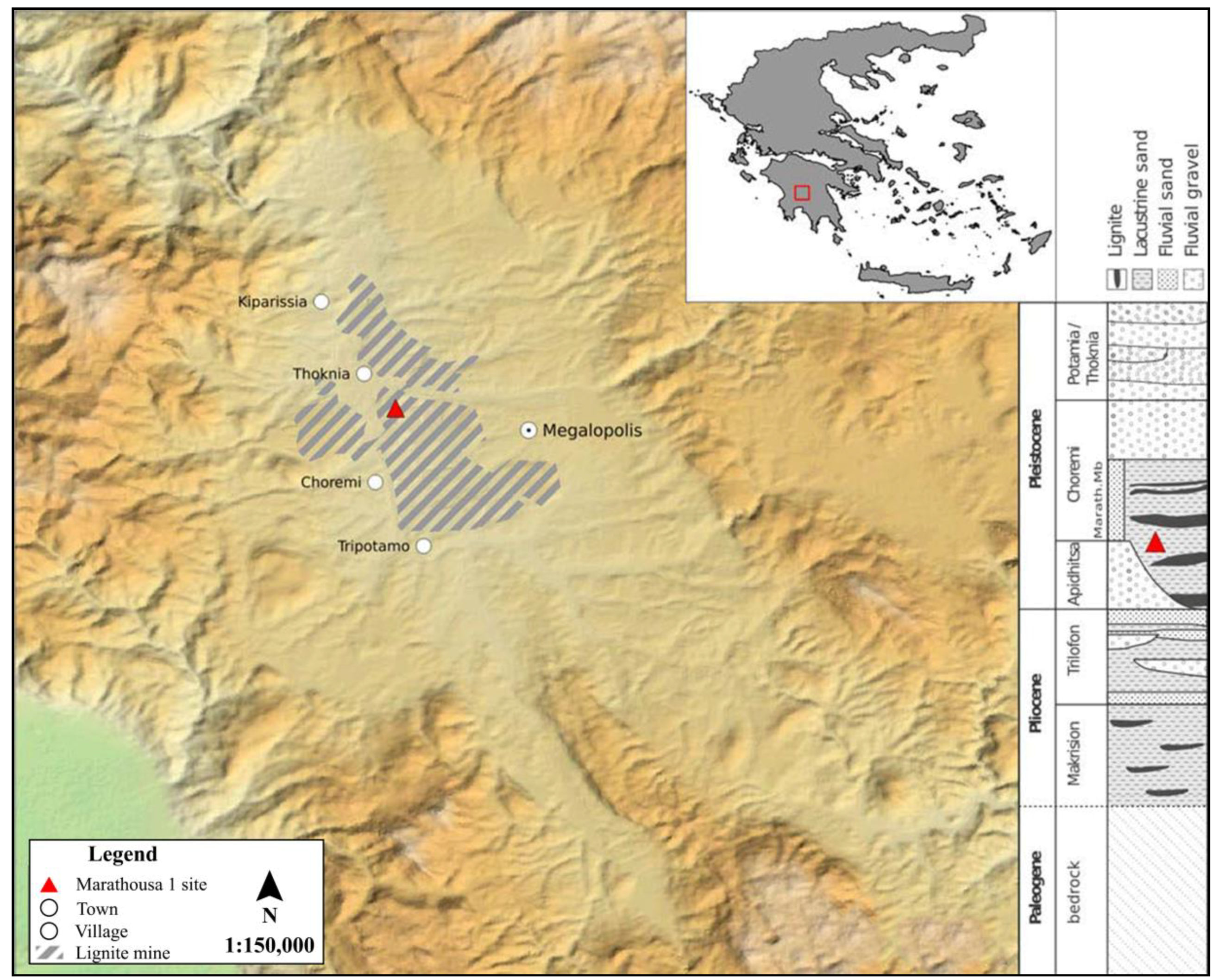
2. Marathousa 1, Megalopolis Basin, Greece
3. ESR Dating
| 𝖠Σ | = | the total accumulated radiation dose in the sample, |
| 𝖠int | = | the internally derived accumulated dose component, |
| 𝖠ext | = | the externally derived accumulated dose component, |
| DΣ(t) | = | the total dose rate, |
| Dint(t) | = | the dose rate from U, its daughters, and other radioisotopes inside the tooth, |
| Dsed(t) | = | the dose rate from sedimentary U, Th, and K, around the fossil, |
| Dcos(t) | = | the dose rate from cosmic sources, |
| t1 | = | the sample’s age, |
| t0 | = | today [33]. |
3.1. Internal Dose Rates
3.2. External Dose Rates
3.3. Sample Preparation
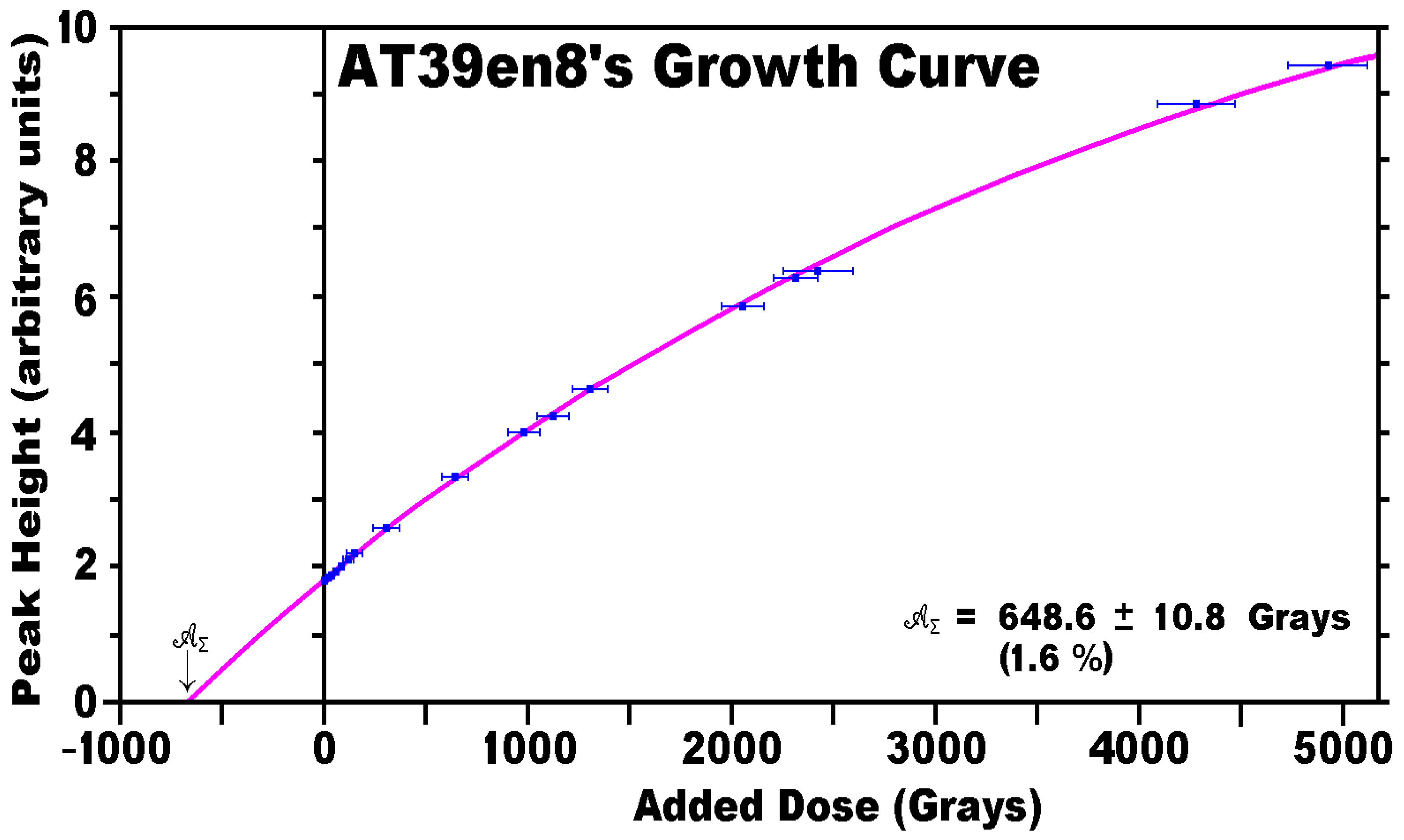
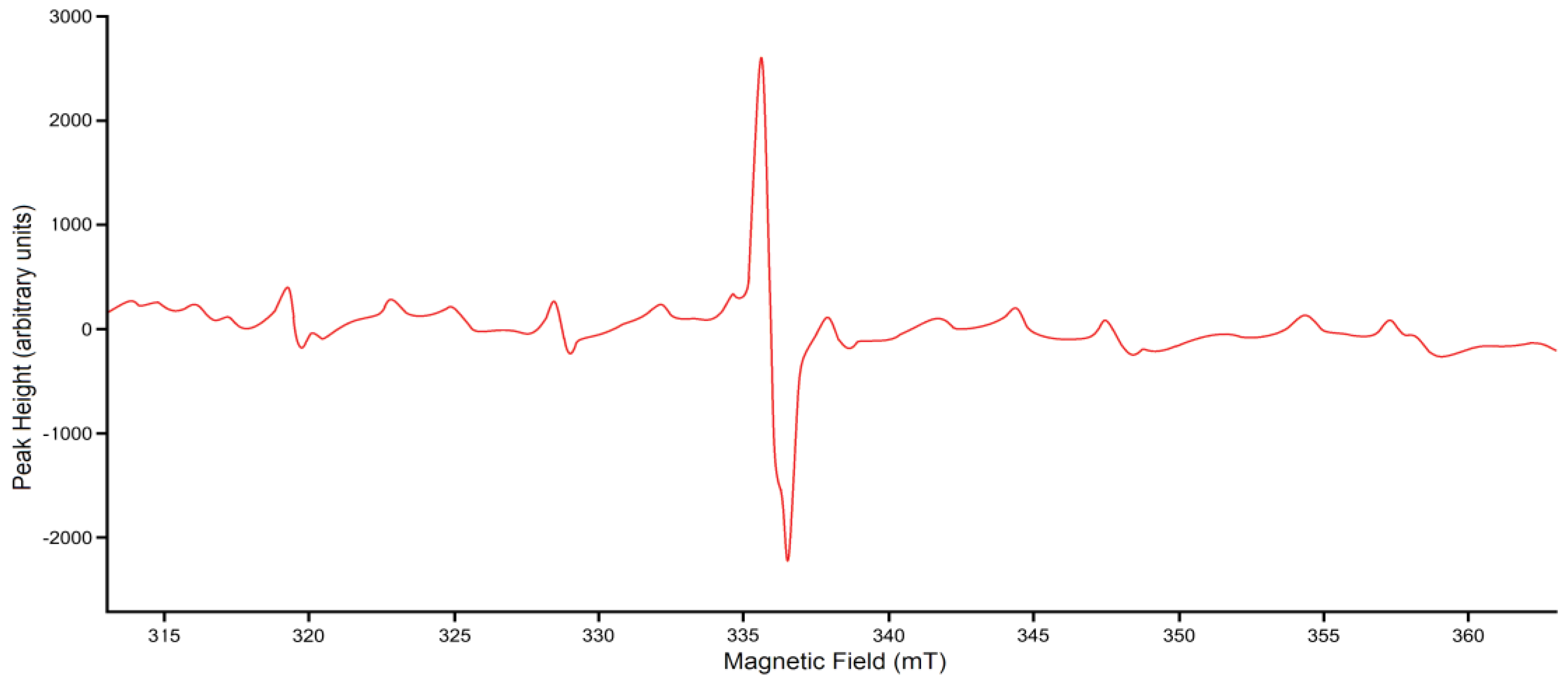
3.4. ESR Data Analyses
3.5. Error Analyses
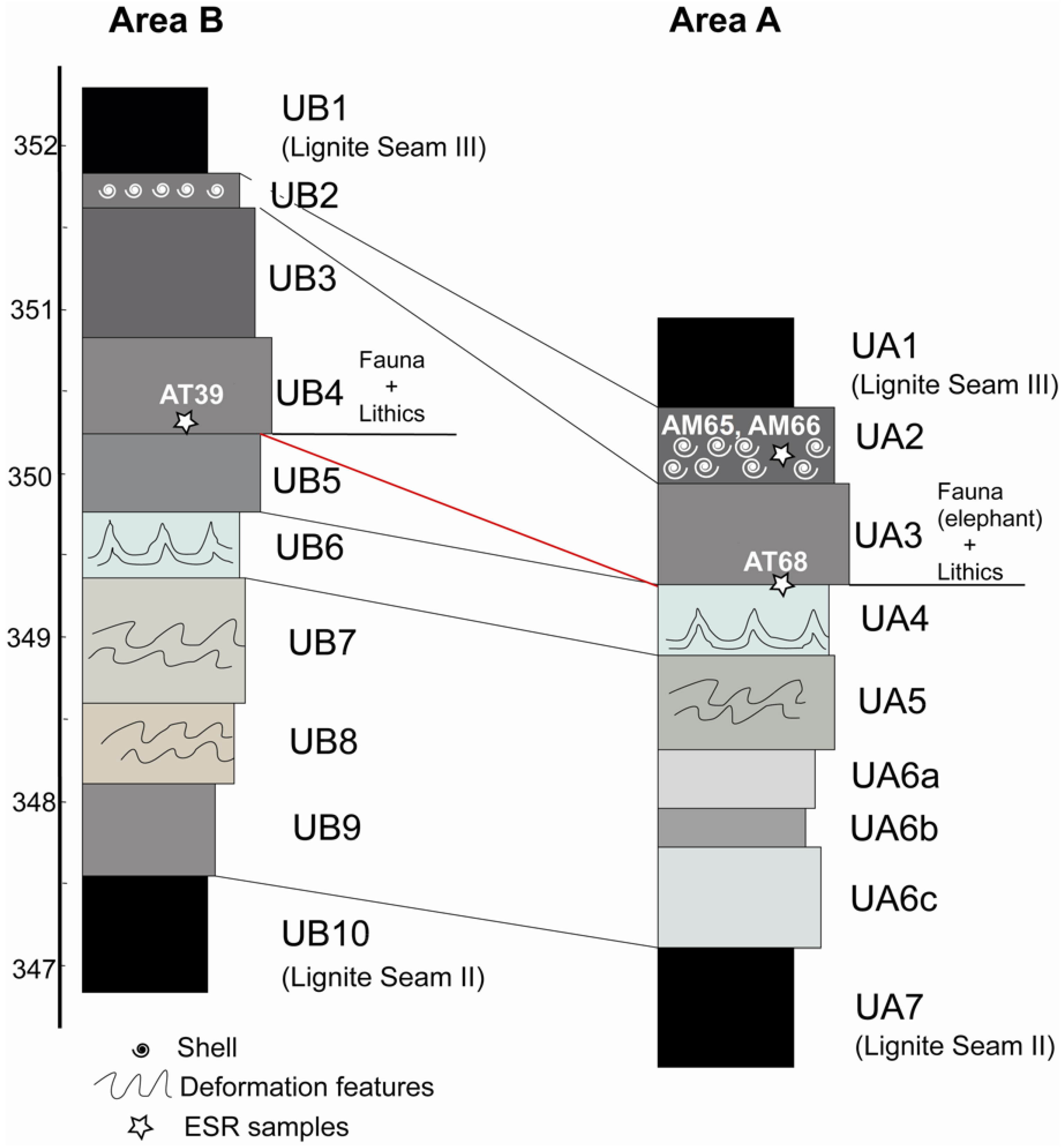
4. Results
4.1. Dating AM66 and AM65
4.2. Dating the Teeth
4.3. Correlations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Definition |
|---|---|
| 𝖠Σ | the total accumulated radiation dose in the dating sample or subsample |
| 𝖠Σ,i | the total accumulated radiation dose in the ith subsample in an isochron analysis |
| 𝖠int | the internally derived accumulated dose component in the dating sample |
| 𝖠ext | the externally derived accumulated dose component in the dating sample |
| DΣ(t) | the total dose rate from all sources for the dating sample |
| Dint(t) | the dose rate from U, its daughters, and other radioisotopes inside the dating subsample |
| Dint,i(t) | the internal dose rate inside the ith subsample in an isochron analysis |
| Dext(t) | the dose rate from all sources outside the dating sample |
| Dsed(t) | the dose rate from sedimentary U, Th, K, and other radioisotopes around the dating sample |
| Dcos(t) | the dose rate from cosmic sources affecting the dating sample |
| t1 | the sample’s age |
| t0 | today |
| [Utooth] | the uranium concentration in the whole tooth |
| [Uen] | the uranium concentration in the enamel for a dated tooth |
| [Uden] | the uranium concentration in the dentine for a dated tooth |
| [Umol] | the uranium concentration in a dated molluscan sample |
| [Used] | the uranium concentration in the sediment around a dating sample |
| [Thsed] | the thorium concentration in the sediment around a dating sample |
| [Ksed] | the potassium concentration in the sediment around a dating sample |
| [Wen] | the water concentration in the enamel in a dated tooth |
| [Wden] | the water concentration in the dentine in a dated tooth |
| [Wcem] | the water concentration in the dental cement in a dated tooth |
| [Wmol] | the water concentration in the in a dated molluscan sample |
| [Wsed] | the water concentration in the sediment around a dating sample |
| [Wsed,pal] | the water concentration in the palustrine sediment around a dating sample |
| [Wsed,all] | the water concentration in the alluvial sediment around a dating sample |
| the time-averaged water concentration in the sediment around a dating sample | |
| ρen | the density of the enamel in a dated tooth |
| ρden | the density of the dentine in a dated tooth |
| ρcem | the density of the dental cementum in a dated tooth |
| ρmol | the density of the carbonate shell in a dated molluscan sample |
| ρsed | the sedimentary density around a dating sample |
| p | the U uptake rate used in calculating Dint(t) |
| EU | the early U uptake model used in calculating Dint(t) with p = −1 |
| LU | the linear (continuous) U uptake model used in calculating Dint(t) with p = 0 |
| RU | any recent U uptake model used in calculating Dint(t), often generally used with p = 10 |
| RUx | the recent U uptake model used in calculating Dint(t) with p = x |
| the external dose rate derived from the bulk sedimentary geochemcial analysis | |
| the sedimentary dose rate from β sources derived from bulk sedimentary geochemical analyses | |
| the sedimentary dose rate from γ sources derived from bulk sedimentary geochemical analyses | |
| the time- and volumetrically averaged internal dose rate | |
| the time-averaged sedimentary dose rate | |
| the time-averaged cosmic dose rate | |
| the time- and volumetrically averaged external dose rate from geochemical and cosmic analyses | |
| the external dose rate derived from the isochron analysis | |
| τ | the mean ESR signal lifetime, a measure of its stability |
| κα | the α efficiency factor |
| (234U/238U)0 | the initial 234U/238U activity ratio |
| MIS | Marine (Oyygen) Isotope Stage |
References
- Harvati, K.; Panagopoulou, E.; Runnels, C. The paleoanthropology of Greece. Evol. Anthropol. 2009, 18, 131–143. [Google Scholar] [CrossRef]
- Tourloukis, V.; Karkanas, P. The Middle Pleistocene archaeological record of Greece and the role of the Aegean in hominin dispersals: New data and interpretations. Quat. Sci. Rev. 2012, 43, 1–15. [Google Scholar] [CrossRef]
- Panagopoulou, E.; Tourloukis, V.; Thompson, N.; Athanassiou, A.; Tsartsidou, G.; Konidaris, G.E.; Giusti, D.; Karkanas, P.; Harvati, K. Marathousa 1: A new Middle Pleistocene archaeological site from Greece. Antiquity Project Gallery 2015, 89, 343. [Google Scholar]
- Harvati, K. Paleoanthropology in Greece: Recent findings and interpretations. In The Paleoanthropology of the Balkans and Anatolia: Human Evolution and Its Context; Harvati, K., Roksandic, M., Eds.; Vertebrate Paleobiology & Paleoanthropology Series; Springer: Dordrecht, The Netherlands, 2016; pp. 3–14. [Google Scholar]
- Tourloukis, V.; Harvati, K. The Palaeolithic record of Greece: A synthesis of the evidence and a research agenda for the future. Quat. Int. 2018, 466, 48–65. [Google Scholar] [CrossRef]
- Vinken, R. Stratigraphie und Tektonik des Beckens von Megalopolis (Peloponnes, Griechenland). Geol. Jahrb. 1965, 83, 97–148. [Google Scholar]
- Van Vugt, N.; de Bruijn, H.; van Kolfschoten, M.; Langereis, C.G.; Okuda, M. Magento- and cyclo- stratigraphy and mammal faunas of Pleistocene lacustrine Megalopolis Basin, Peloponnesos, Greece. Geol. Ultrajectina 2000, 189, 69–92. [Google Scholar]
- Van Vugt, N.; Langereis, C.G.; Hilgen, F.J. Orbital forcing in Pliocene-Pleistocene Mediterranean lacustrine deposits: Dominant expression of eccentricity versus precession. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 172, 193–205. [Google Scholar] [CrossRef]
- Okuda, M.; van Vugt, N.; Nakagawa, T.; Ikeya, M.; Hayashida, A.; Yusada, Y.; Setoguchi, T. Palynological evidence for the astromonical origin of lignite-detritus sequences in the Middle Pleistocene Marathousa Member, Megalopolis, SW Greece. Earth Planet. Sci. Lett. 2002, 201, 143–157. [Google Scholar] [CrossRef]
- Tourloukis, V.; Muttoni, G.; Karkanas, P.; Monesi, E.; Scardia, G.; Panagopoulou, E.; Harvati, K. Magnetostratigraphic and chronostratigraphic constraints on the Marathousa 1 Lower Palaeolithic site and the Middle Pleistocene deposits of the Megalopolis basin, Greece. Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Thompson, N.; Touloukis, V.; Panagopoulou, E.; Harvati, K. In search of Pleistocene remains at the Gates of Europe: Results from the ERC Starting Grant project ‘Paleanthropology at the Gates of Europe’ (PaGE) directed survey of the Megalopolis Basin. Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Tourloukis, V.; Thompson, N.; Panagopoulou, E.; Giusti, D.; Konidaris, G.E.; Karkanas, P.; Harvati, K. Lithic artifacts and bone tools from the Lower Palaeolithic site Marathousa 1, Megalopolis, Greece: Preliminary results. Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Panagopoulou, E.; Tourloukis, V.; Thompson, N.; Konidaris, G.E.; Athanassiou, A.; Giusti, D.; Tsartsidou, G.; Karkanas, P.; Harvati, K. The Lower Palaeolithic site of Marathousa 1, Megalopolis, Greece: Overview of the evidence. Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Karkanas, P.; Tourloukis, V.; Thompson, N.; Giusti, D.; Panagopoulou, E.; Harvati, K. Sedimentology and micromorphology of the Lower Palaeolithic lakeshore site Marathousa 1, Megalopolis, Greece. Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Athanassiou, A.; Tourloukis, V.; Thompson, N.; Giusti, D.; Panagopoulou, E.; Harvati, K. The skeleton of a straight-tasked elephant (Palaeoloxodon antiquus) and other large mammals from the Middle Pleistocene butchering locality Marathousa 1 (Megalopolis Basin, Greece): Preliminary results. Quat. Int. 2018, in press. [Google Scholar]
- Blackwell, B.A.B.; Liang, S.S.; Golovanova, L.V.; Doronichev, V.B.; Skinner, A.R.; Blickstein, J.I.B. ESR at Treugol’naya Cave, northern Caucasus Mt., Russia: Dating Russia’s oldest archaeological site and paleoclimatic change in Oxygen Isotope Stage 11. Appl. Radiat. Isot. 2005, 62, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.G. The Human Career: Human Biological and Cultural Origins, 3rd ed.; University of Chicago Press: Chicago, IL, USA, 2009. [Google Scholar]
- Doukas, C.; van Kolfschoten, T.; Papayanni, K.; Panagopoulou, E.; Harvati, K. The small mammal fauna from the Palaeolithic site Marathousa 1 (Greece). Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Blackwell, B.A.B.; Skinner, A.R.; Blickstein, J.I.B.; Montoya, A.C.; Florentin, J.A.; Baboumian, S.M.; Ahmed, I.J.; Deely, A.E. ESR in the 21st Century: From buried valleys and deserts to the deep ocean and tectonic uplift. Earth Sci. Rev. 2016, 158, 125–159. [Google Scholar] [CrossRef]
- Rink, W.J. Beyond 14C dating: A user’s guide to long-range dating methods in archaeology. Earth Sci. Archaeol. 2001, 27, 975–1005. [Google Scholar]
- Schellmann, G.; Beerten, K.; Radtke, U. Electron spin resonance (ESR) dating on Quaternary materials. Eiszeitalt. Ggw. 2008, 57, 150–178. [Google Scholar]
- Skinner, A.F. Electron spin resonance (ESR) dating: General principles. In Encyclopedia of Scientific Dating Methods; Rink, W.J., Thmpson, J., Eds.; Springer: Heidelberg, Germany, 2015. [Google Scholar]
- Blackwell, B.A.B. Electron spin resonance (ESR) dating in lacustrine environments. In Tracking Environmental Change Using Lake Sediment 1: Basin Analyses, Coring, and Chronological Techniques; Last, W.M., Smol, J.P., Eds.; Kluwer: Dordrect, The Netherlands, 2001; pp. 283–368. [Google Scholar]
- Blackwell, B.A.B. Electron spin resonance (ESR) dating in karst environments. Acta carsol. 2006, 35, 123–147. [Google Scholar] [CrossRef]
- Skinner, A.R.; Shawl, C.E. ESR dating of terrestrial Quaternary shells. Quat. Sci. Rev. 1994, 13, 679–684. [Google Scholar] [CrossRef]
- Blackwell, B.A.B.; Kim, D.M.Q.; Curry, B.B.; Grimly, D.A.; Blickstein, J.I.B.; Skinner, A.R. Shell we date? ESR dating Sangamon Interglacial Episode deposits at Hopwood Farm, IL. Radiat. Prot. Dosim. 2016, 172, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Skinner, A.R.; Blackwell, B.A.B.; Chasteen, D.E.; Shao, J.M.; Min, S.S. Improvements in dating tooth enamel by ESR. Appl. Radiat. Isot. 2000, 52, 1337–1344. [Google Scholar] [CrossRef]
- Skinner, A.R.; Blackwell, B.A.B.; Chasteen, D.E.; Shao, J.M. Q-band ESR studies of fossil tooth enamel. Quat. Sci. Rev. 2001, 20, 1027–1030. [Google Scholar] [CrossRef]
- Sayin, U.; Bakkal, G.; Aydin, H.; Isik, M.; Delikan, A.; Yesilyurt, S.K.; Demir, A.; Biyik, R.; Engin, B.; Orhan, H.; Ozmen, A. ESR dating of mollusc shells from Adkale location in Konya Closed Basin, Turkey. In Proceedings of the Solid State Dosimetry 18th Conference (SSD18), Munich, Germany, 3–8 July 2016. [Google Scholar]
- Blackwell, B.A. Electron spin resonance dating. In Dating Methods for Quaternary Deposit; Rutter, N.W., Catto, N.R., Eds.; Geological Association of Canada, GEOtext: St. John’s, NF, Canada, 1995; Volume 2, pp. 209–251. [Google Scholar]
- Vichaidid, T.; Youngchuay, U.; Limsuwan, P. Dating of aragonite fossil shell by ESR for paramagnetic species assignment of Mae Moh basin. Nucl. Instrum. Methods Phys. Res. Sect. 2007, B62, 323–328. [Google Scholar] [CrossRef]
- Blackwell, B.A.B.; Skinner, A.R.; Smith, J.R.; Hill, C.L.; Churcher, C.S.; Kieniewicz, J.M.; Adelsberger, K.A.; Blickstein, J.I.B.; Florentin, J.A.; Deely, A.E.; Spillar, K.V. ESR analyses for herbivore teeth and molluscs from Kharga, Dakhleh, and Bir Tarfawi Oases: Constraining water availablility and hominin Paleolithic activity on the Western Desert, Egypt. J. Afr. Earth Sci. 2017, 136, 216–238. [Google Scholar] [CrossRef]
- Deely, A.E.; Blackwell, B.A.B.; Mylroie, J.E.; Carew, J.L.; Blickstien, J.I.B.; Skinner, A.R. Testing cosmic dose rate models for ESR: Dating corals and molluscs on San Salvador, Bahamas. Radiat. Meas. 2011, 46, 853–859. [Google Scholar] [CrossRef]
- Blackwell, B.A.B.; Skinner, A.R.; Brassard, P.; Blickstein, J.I.B. U uptake in tooth enamel: Lessons from isochron analyses and laboratory simulation experiments. In Proceedings of the International Symposium on New Prospects in ESR Dosimetry and Dating, Advances in ESR Applications; Whitehead, N.E., Ikeya, M., Eds.; Society of ESR Applied Metrology: Osaka, Japan, 2002; Volume 18, pp. 97–118. [Google Scholar]
- Blackwell, B.A.; Schwarcz, H.P. ESR isochron dating for teeth: A brief demonstration in solving the external dose calculation problem. Appl. Radiat. Isot. 1993, 44, 243–252. [Google Scholar] [CrossRef]
- Dibble, H.L.; Aldeias, V.; Alvarez-Fernández, E.; Blackwell, B.A.; Hallett-Desguez, E.; Jacobs, Z.; Goldberg, P.; Lin, S.C.; Morala, A.; Meyer, M.C.; et al. New excavations at the site of Contrebandiers Cave, Morocco. Paleoanthropology 2012, 2012, 145–201. [Google Scholar]
- Adamiec, G.; Aitken, M.J. Dose rate conversion factors: Update. Anc. TL 1998, 16, 37–50. [Google Scholar]
- Blackwell, B.A.B.; Blickstein, J.I.B. Considering sedimentary U uptake in external dose rate determinations for ESR and luminescent dating. Quat. Int. 2000, 68, 329–343. [Google Scholar] [CrossRef]
- Prescott, J.R.; Hutton, J.T. Cosmic ray contribution to dose rates for luminescence and ESR dating: Large depths and long-term variabtion. Radiat. Meas. 1994, 2–3, 497–500. [Google Scholar] [CrossRef]
- Blackwell, B.A. Problems associated with reworked teeth in electron spin resonance (ESR) dating. Quat. Sci. Rev. 1994, 13, 651–660. [Google Scholar] [CrossRef]
- Giusti, D.; Tourloukis, V.; Konidaris, G.E.; Thompson, N.; Karkanas, P.; Panagopoulou, E.; Harvati, K. Beyond maps: Patterns of formation processes at the Middle Pleistocene open-air site of Marathousa 1, Megalopolis Basin, Greece. Quat. Int. 2018, in press. [Google Scholar] [CrossRef]
- Blackwell, B.A. Laboratory Procedures for ESR Dating of Tooth Enamel; Department of Geology Technical Memo 89.1; McMaster University: Hamilton, ON, Canada, 1989; 234p. [Google Scholar]
- Lee, H.K.; Rink, W.J.; Schwarcz, H.P. Comparison of ESR signal dose-responses in modern and fossil tooth enamel. Radiat. Meas. 1997, 27, 405–411. [Google Scholar] [CrossRef]
- Skinner, A.R.; Blackwell, B.A.B.; Hasan, M.M.; Blickstein, J.I.B. Expanding the range of electron spin resonance dating. In Archaeological Chemistry: Analytical Techniques and Archaeological Interpretation; Glascock, M.D., Speakman, R.J., Popelka-Filcoff, R.S., Eds.; ACS Symposium Series 968; American Chemical Society: Washington, DC, USA, 2007; pp. 1–14. [Google Scholar]
- Duval, M.; Grün, R. Are published ESR dose assessments on tooth enamel reliable? Quat. Geochronol. 2016, 31, 19–27. [Google Scholar] [CrossRef]
- Brennan, B.J.; Rink, W.J.; McGuirl, E.L.; Schwarcz, H.P. β doses in tooth enamel by “one-group” theory and the Rosy ESR dating software. Radiat. Meas. 1997, 27, 307–314. [Google Scholar] [CrossRef]
- Grün, R. Electron spin resonance (ESR) dating. Quat. Int. 1989, 1, 65–109. [Google Scholar] [CrossRef]
- Barabas, M.; Walther, R.; Wieser, A.; Radtke, U.; Grün, R. Second interlaboratory-comparison project on ESR dating. Appl. Radiat. Isot. 1993, 44, 119–129. [Google Scholar] [CrossRef]
- Wieser, A.; Debuyst, R.; Fattibene, P.; Meghzifene, A.; Onori, S.; Brik, A.; Scherbina, O.; Bayankin, S.N.; Ivanov, D.; Pivovarov, S.; et al. The 3rd International Intercomparison on EPR Tooth Dosimetry: Part 2, final analysis. Radiat. Prot. Dosim. 2005, 120, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.F.; Bender, M.L.; Broecker, W.S. Surface of biogenic carbonates and their relationship to ultrastructure and diagenesis. J. Sediment. Petrol. 1973, 43, 471–477. [Google Scholar]
- Ayling, B.F.; Eggins, S.; McCulloch, M.T.; Chappell, J.; Grün, R.; Mortimer, G. Uranium uptake history, open-system behaviour and uranium-series ages of fossil Tradacna gigas from the Huon Peninsula, Papua New Guinea. Geochim. Cosmochim. Acta 2017, 213, 475–501. [Google Scholar] [CrossRef]
- Florentin, J.A.; Blackwell, B.A.B.; Tüysüz, O.; Tarı, U.; Genç, Ş.C.; İmren, C.; Mo, S.; Huang, Y.E.W.; Blickstein, J.I.B.; Skinner, A.R.; et al. ESR dating molluscs from marine terraces near Mağaracık and Samandağ, Hatay Province, Turkey. Radiat. Prot. Dosim. 2014, 159, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, Z.; Li, B.; Karkanas, P.; Tourloukis, V.; Thompson, N.; Panagopoulou, E.; Harvati, K. Dating of the Middle Pleistocene Marathousa 1 (Greece) lacustrine sediment using multiple aliquot pre-dose multi- elevated temperature post-infrared stimulated luminescence (MET-pIRIR). Quat. Int. 2018. in review. [Google Scholar]
- Tzedakis, P.C.; Hooghiemstra, H.; Pälike, H. The last 1.35 million years at Tenaghi Philippon: Revised chronostratigraphy and long-term vegetation trends. Quat. Sci. Rev. 2006, 25, 3416–3420. [Google Scholar] [CrossRef]
- Blackwell, B.A. Problems of Amino Acid Racemization Dating Analyses: Bones and Teeth from the Archeological Sites Lachaise and Montgaudier (Charente, France). Unpub. Ph.D., University of Alberta, Edmonton, AB, Canada, 1987; 650p. [Google Scholar]
- Blackwell, B.; Schwarcz, H.P. Archaeochronology and scale. In The Effects of Scale on Archaeological and Geoscientific Perspectives; Stein, J.K., Linse, A., Eds.; Geological Society of America Special Paper 283; Geological Society of Amer: Boulder, CO, USA, 1993; pp. 39–58. [Google Scholar]
- McDermott, F.; Grün, R.; Stringer, C.; Hawkesworth, C. Mass spectrometric U-series dates for Israeli Neanderthal/early modern hominid sites. Nature 1993, 363, 252–254. [Google Scholar] [CrossRef] [PubMed]
- Grün, R.; McDermott, F. Open system modelling for U-series and ESR dating of teeth. Quat. Sci. Rev. 1994, 13, 121–125. [Google Scholar] [CrossRef]
- Jones, H.L.; Rink, W.J.; Schepartz, L.A.; Miller-Antonio, S.; Huang, W.; Hou, Y.; Wang, W. Coupled electron spin resonance (ESR)/uranium-series dating of mammalian tooth enamel at Panxian Dadong, Guizhou Province, China. J. Archeaol. 2004, 31, 965–977. [Google Scholar] [CrossRef]
- Duval, M.; Falguères, C.; Bajaom, J.J.; Grün, R.; Shao, Q.; Aubert, M.; Dolo, J.M.; Agusto, J.; Martinez Navarro, B.; Palmqvist, P.; et al. On the limits of using combined U-series/ESR method to date fossil teeth from two Early Pleistocene archaeological sites of the Orce area (Guadix-Baza Basin, Spain). Quat. Res. 2012, 77, 482–491. [Google Scholar] [CrossRef]
- Ivanovich, M.; Latham, A.; Ku, T.L. Uranium series disequilibrium applications in geochrology. In Uranium Series Disequilibrium: Application to Environmental Problems, 2nd ed.; Ivanovich, M., Harmon, R.S., Eds.; Clarendon: Oxford, UK, 1992; pp. 62–94. [Google Scholar]
- Schwarcz, H.P.; Blackwell, B.A. Uranium series dating of archaeological sites. In Uranium Series Disequilibrium: Application to Environmental Problems, 2nd ed.; Ivanovich, M., Harmon, R.S., Eds.; Clarendon: Oxford, UK, 1992; pp. 513–552. [Google Scholar]
- Qi, J.K.; Blackwell, B.A.B.; Singh, I.K.; Stepanchuk, V.N.; Blickstein, J.I.B.; Florentine, J.A.; Skinner, A.R. Preliminary results of dating for the Lower Paleolithic sites of Ukraine (Medzhibozh 1 and Medzhibozh A, Khmelnitskii region) by electron spin resonance method. Geophys. J. 2018, 40, 155–177. [Google Scholar]
- Lang, N.; Wolff, E.W. Interglacial and glacial variability from the last 800 ka in marine, ice, and terrestrial archives. Clim. Past 2011, 7, 361–380. [Google Scholar] [CrossRef] [Green Version]








| Number | Location | Sample Type | ||||
|---|---|---|---|---|---|---|
| ESR | ESR | Site | Layer/ | Depth | Species/ | Tooth/ |
| Analysis | Catalogue | Area | Boundary | (cm) | Taxon | Shell Part |
| AM65 | 2014.009a1 | A | UA2 2 | 349.94 | bivalve | outer shell |
| AM66 | 2014.009a3 | A | UA2 2 | 349.94 | bivalve | nacreous layer |
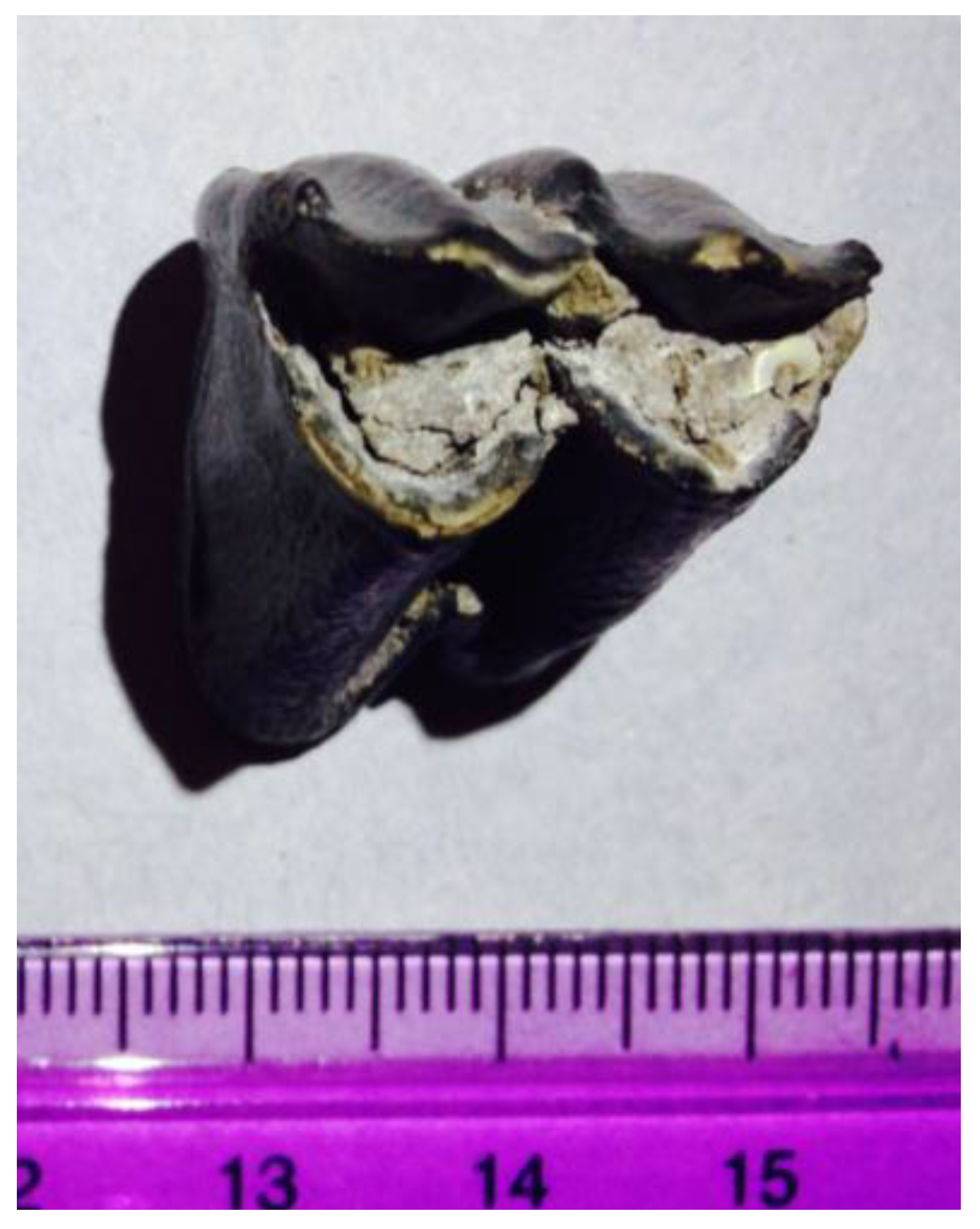
| AT68 | 2016.001 | A | UA3c/UA4 1 | 350.19 | Cervus elaphus | molar |
| AT39 | 2014.008 | B | UB4c/UB5 1 | 350.33 | cervid | molar |
| Sample | U Concentrations | Internal dose rates, Dint(t) | |||||
| Enamel/ | Inner | Outer | Enamel/ | Inner | Outer | ||
| Shell | Dentine | Dentine | Shell | Dentine | Dentine | ||
| (ppm) | (ppm) | (ppm) | (µGy/y) | (µGy/y) | (µGy/y) | ||
| a. AT39, 2014.008, cervid cheek tooth, Area B 1 | |||||||
| AT39en1 | - | 18.41 | - | ||||
| AT39en2 | - | 16.02 | - | ||||
| AT39en3 | - | 11.41 | - | ||||
| AT39en1-3 | 2.08 | 15.28 | 21.60 | 0.230 | 0.134 | 0.190 | |
| AT39en4 | 1.50 | 26.45 | 15.71 | 0.171 | 0.189 | 0.113 | |
| AT39en5 | 1.53 | 14.72 | 17.55 | 0.176 | 0.100 | 0.120 | |
| AT39en6 | - | 14.78 | - | ||||
| AT39en7 | - | 19.18 | - | ||||
| AT39en6+7 | 0.79 | 16.98 | 19.98 | 0.089 | 0.138 | 0.162 | |
| AT39en8 | 1.90 | 32.87 | 17.87 | 0.218 | 0.222 | 0.121 | |
| AT39 mean | 1.56 | 19.23 | 18.54 | 0.177 | 0.156 | 0.141 | |
| ± | 0.50 | 7.08 | 2.28 | 0.055 | 0.048 | 0.033 | |
| b. AT68, 2016.001, cervid molar, Area A 1 | |||||||
| AT68en1-9 | 0.07 | 12.97 | 15.19 | 0.008 | 0.123 | 0.145 | |
| ± | 0.02 | 0.02 | 0.02 | 0.003 | 0.009 | 0.012 | |
| c. 2014.009, bivalves, Area A 2 | |||||||
| AM66 | 0.01 | - | - | 0.002 | - | - | |
| AM65 | 0.01 | - | - | 0.002 | - | - | |
| mean | 0.01 | - | - | 0.002 | - | - | |
| bivalves | ± | 0.01 | - | - | 0.002 | - | - |
| Analytical | ± | 0.01 | 0.01 | 0.01 | 0.002 | 0.009 | 0.009 |
| uncertainties 3 | - | 0.02 | 0.02 | 0.02 | 0.032 | 0.016 | 0.033 |
| Detection | ~ | 0.01 | 0.01 | 0.01 | |||
| limit 3 | - | 0.02 | 0.02 | 0.02 | |||
| Typical water | ~ | 0.02 | 0.05 | 0.05 | |||
| concentrations 3 | - | 0.02 | 0.02 | 0.02 | |||
- 1
- Dint(t) calculated assuming a recent uptake model (RU) with p = 2.
- 2
- Dint(t) calculated assuming a linear uptake model (LU) with p = 0.
- 3
- Uncertainties, typical detection limits, and water concentrations depend on the sample’s mass, mineralogy, and tissue type.
| Sediment | Location | Concentrations | Sedimentary Dose Rates 1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample | Area | [U] | [Th] | [K] | |||||
| Type | Locus | (ppm) | (ppm) | (wt%) | (mGy/y) | (mGy/y) | (mGy/y) | (mGy/y) | |
| 2014.014 | B | 4.45 | 7.44 | 1.10 | 0.303 | 1.031 | 0.206 | 0.719 | |
| bulk sediment | ± | 0.02 | 0.16 | 0.03 | 0.029 | 0.065 | 0.022 | 0.060 | |
| AT39sed1 | B | 3.92 | 7.22 | 1.71 | 0.368 | 1.098 | 0.251 | 0.766 | |
| bulk sediment | ± | 0.02 | 0.53 | 0.06 | 0.035 | 0.073 | 0.028 | 0.066 | |
| AT39sed2 6 | B | 17.44 6 | 4.84 | 0.11 | 0.514 | 2.012 | 0.350 | 1.405 | |
| attached sediment | ± | 0.02 | 0.72 | 0.02 | 0.046 | 0.129 | 0.037 | 0.119 | |
| Mean near AT39 | B | 8.60 | 6.51 | 0.98 | 0.396 | 1.382 | 0.269 | 0.965 | |
| (n = 3) | ± | 7.66 | 1.45 | 0.81 | 0.238 | 0.802 | 0.163 | 0.562 | |
| Mean near AT39 7 | B | 4.19 | 7.34 | 1.41 | 0.336 | 1.065 | 0.229 | 0.744 | |
| (n = 2) | ± | 0.38 | 0.16 | 0.43 | 0.046 | 0.047 | 0.047 | 0.093 | |
| 2014.012 | A | 2.68 | 4.63 | 0.46 | 0.372 | 0.584 | 0.254 | 0.408 | |
| bulk sediment | 25 cm > shells | ± | 0.02 | 0.11 | 0.01 | 0.035 | 0.036 | 0.029 | 0.034 |
| 2014.010 | A | 2.95 | 5.21 | 0.75 | 0.516 | 0.701 | 0.334 | 0.489 | |
| bulk sediment | shell layer | ± | 0.02 | 0.14 | 0.02 | 0.048 | 0.045 | 0.037 | 0.041 |
| 2014.013 | A | 3.27 | 4.92 | 0.47 | 0.417 | 0.659 | 0.284 | 0.460 | |
| bulk sediment | shell layer | ± | 0.02 | 0.12 | 0.01 | 0.040 | 0.042 | 0.032 | 0.038 |
| 2014.011 | A | 4.44 | 5.91 | 1.02 | 0.682 | 0.942 | 0.465 | 0.657 | |
| bulk sediment | 25 cm < shells | ± | 0.02 | 0.13 | 0.03 | 0.066 | 0.060 | 0.053 | 0.054 |
| Mean near AM66 | 3.34 | 5.17 | 0.68 | 0.493 | 0.723 | 0.335 | 0.505 | ||
| (n = 4) | ± | 0.78 | 0.55 | 0.27 | 0.112 | 0.111 | 0.079 | 0.082 | |
| Typical detection limits 8 | ~ | 0.01 | 0.10 | 0.002 | 0.002 | 0.010 | 0.002 | 0.008 | |
| – | 0.02 | 0.20 | 0.004 | 0.004 | 0.020 | 0.003 | 0.016 | ||
| NIST 1633B standard | 8.79 | 25.70 | 1.950 | – | – | – | – | ||
| ± | 0.02 | 0.70 | 0.005 | – | – | – | – | ||
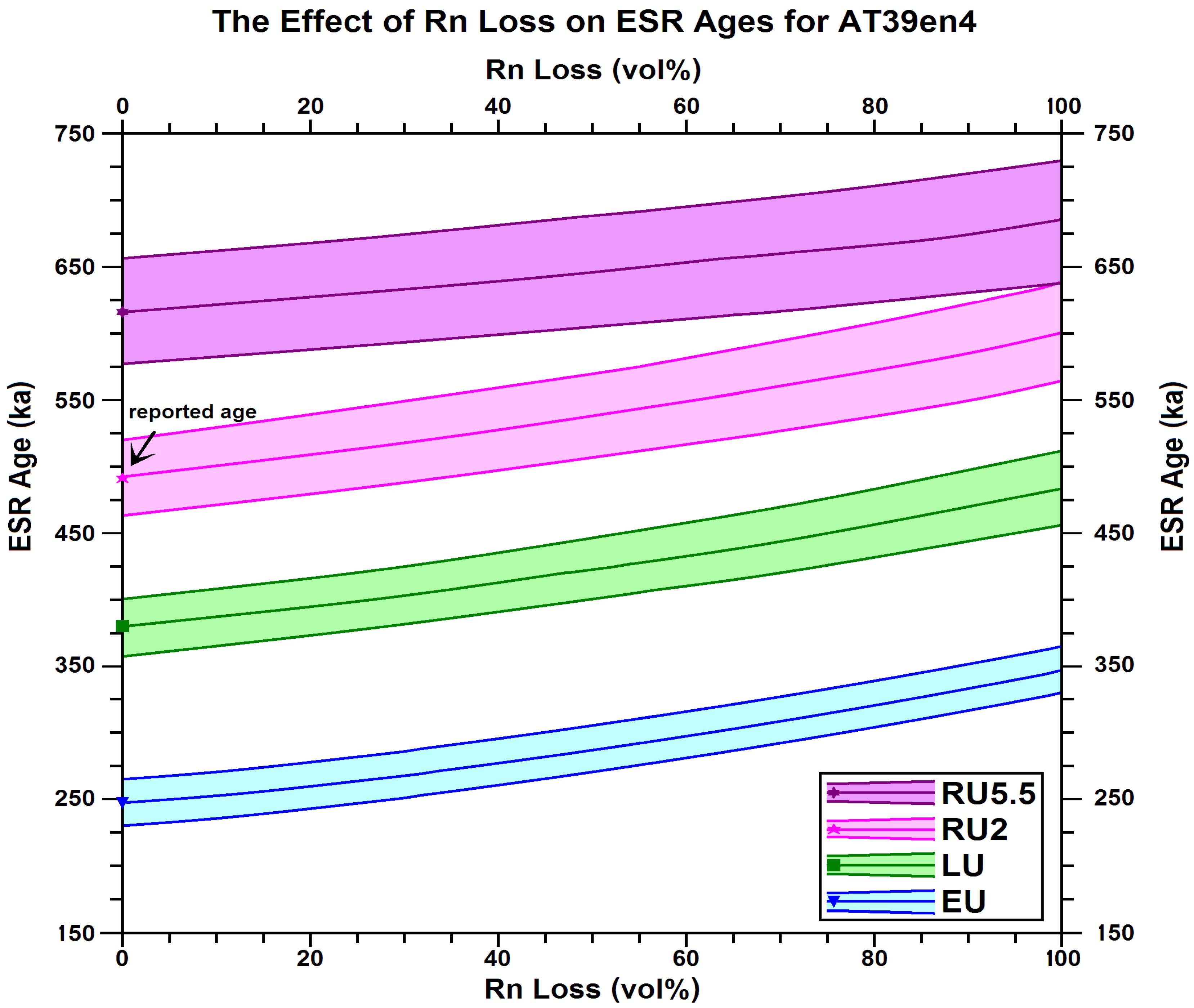
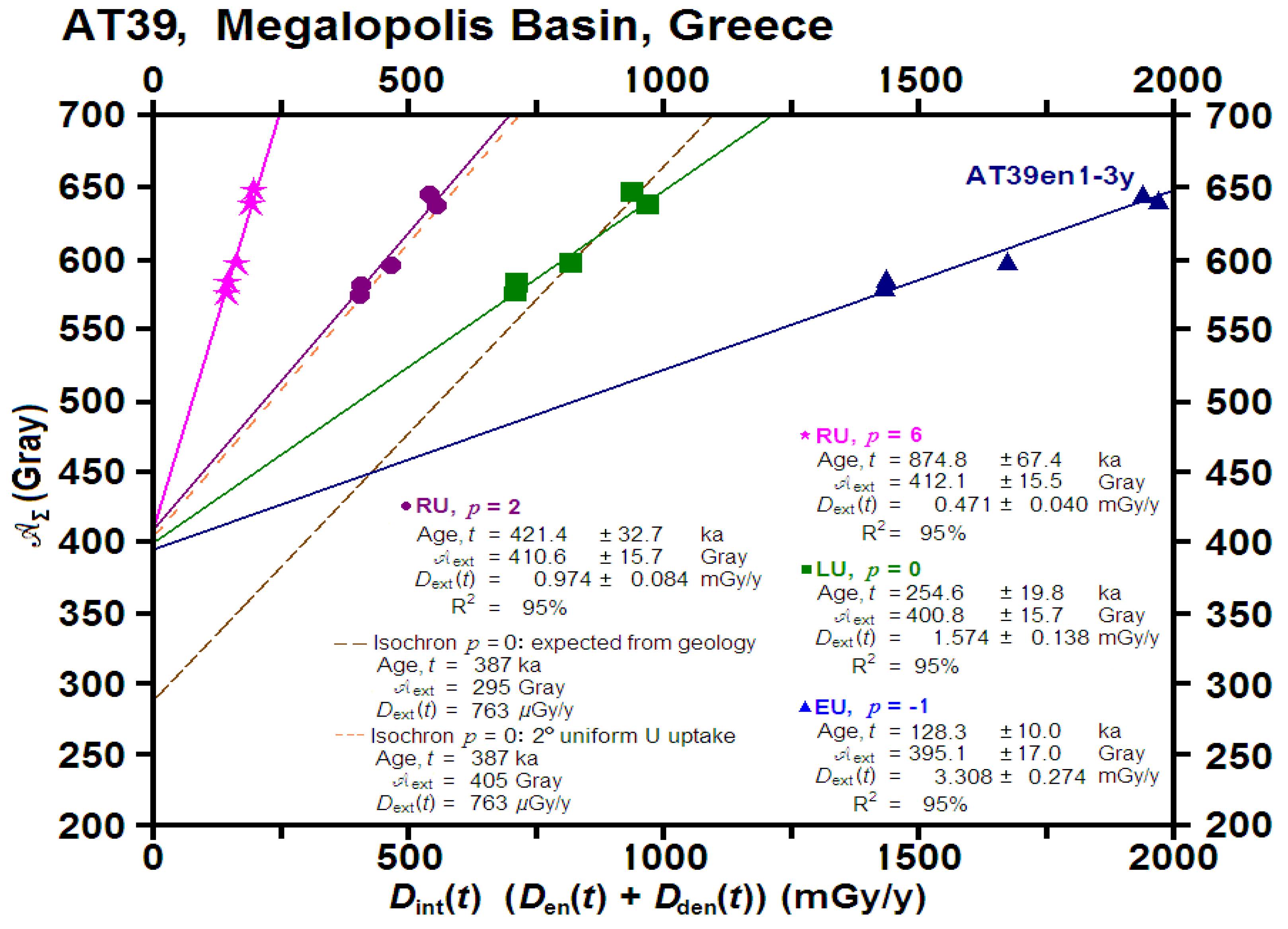
| Accumulated | External Dose Rates | Standard ESR Age 1,2,3,4 | ||||||
| Sample | Dose, 𝖠Σ | EU | LU | RU2 | RU8 | |||
| (Grays) | (μGy/y) | (μGy/y) | (ka) | (ka) | (ka) | (ka) | ||
| a. AT39, 2014.008, cervid tooth, Area B | ||||||||
| AT39en1-3y | 649.0 | 680.0 | 80.0 | 241.6 | 378.6 | 501.0 | 670.3 | |
| ± | 22.5 | 42.0 | 8.0 | 14.9 | 22.0 | 29.7 | 43.5 | |
| AT39en4 | 596.7 | 681.0 | 80.0 | 245.8 | 377.3 | 491.0 | 638.5 | |
| ± | 20.1 | 42.0 | 8.0 | 14.0 | 21.0 | 28.7 | 41.5 | |
| AT39en5 | 582.9 | 665.0 | 85.0 | 268.6 | 404.1 | 514.9 | 650.8 | |
| ± | 17.8 | 41.0 | 9.0 | 15.7 | 22.7 | 30.3 | 42.7 | |
| AT39en6+7 | 576.0 | 666.0 | 84.0 | 267.8 | 401.8 | 512.1 | 641.1 | |
| ± | 16.4 | 41.0 | 9.0 | 13.6 | 20.8 | 28.1 | 41.1 | |
| AT39en8 | 648.6 | 686.0 | 80.0 | 240.5 | 374.3 | 497.4 | 666.2 | |
| ± | 10.8 | 42.0 | 9.0 | 12.5 | 18.2 | 26.1 | 38.6 | |
| AT39 mean | 621.7 | 676.0 | 81.0 | 253.3 | 387.0 | 503.2 | 654.0 | |
| (n = 2) | ± | 7.1 | 19.0 | 4.0 | 6.3 | 9.3 | 12.7 | 18.5 |
| b. AT68, 2016.001, cervid molar, Area A | ||||||||
| AT39en8 | 567.5 | 684.0 | 87.0 | 303.3 | 424.4 | 512.1 | 643.4 | |
| ± | 27.2 | 42.0 | 9.0 | 18.3 | 26.2 | 34.2 | 49.4 | |
| c. Bivalve shells, Area A | ||||||||
| AM66 | 233.8 | 444.0 | 32.0 | 487.9 | 488.8 | 489.9 | 490.8 | |
| 2014.009a3 | ± | 5.9 | 45.0 | 6.0 | 53.2 | 53.6 | 53.8 | 54.0 |
| AM65 | 233.5 | 444.0 | 32.0 | 486.0 | 488.1 | 489.1 | 490.1 | |
| 2014.009a1 | ± | 3.5 | 45.0 | 6.0 | 52.2 | 52.6 | 52.8 | 53.0 |
| mean | 233.6 | 444.0 | 32.0 | 486.9 | 488.4 | 489.5 | 490.4 | |
| (n = 2) | ± | 3.0 | 45.0 | 6.0 | 37.3 | 37.5 | 37.6 | 37.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blackwell, B.A.B.; Sakhrani, N.; Singh, I.K.; Gopalkrishna, K.K.; Tourloukis, V.; Panagopoulou, E.; Karkanas, P.; Blickstein, J.I.B.; Skinner, A.R.; Florentin, J.A.; et al. ESR Dating Ungulate Teeth and Molluscs from the Paleolithic Site Marathousa 1, Megalopolis Basin, Greece. Quaternary 2018, 1, 22. https://doi.org/10.3390/quat1030022
Blackwell BAB, Sakhrani N, Singh IK, Gopalkrishna KK, Tourloukis V, Panagopoulou E, Karkanas P, Blickstein JIB, Skinner AR, Florentin JA, et al. ESR Dating Ungulate Teeth and Molluscs from the Paleolithic Site Marathousa 1, Megalopolis Basin, Greece. Quaternary. 2018; 1(3):22. https://doi.org/10.3390/quat1030022
Chicago/Turabian StyleBlackwell, Bonnie A. B., Neeraj Sakhrani, Impreet K. Singh, Kalyani K. Gopalkrishna, Vangelis Tourloukis, Eleni Panagopoulou, Panagiotis Karkanas, Joel I. B. Blickstein, Anne R. Skinner, Jonathan A. Florentin, and et al. 2018. "ESR Dating Ungulate Teeth and Molluscs from the Paleolithic Site Marathousa 1, Megalopolis Basin, Greece" Quaternary 1, no. 3: 22. https://doi.org/10.3390/quat1030022
APA StyleBlackwell, B. A. B., Sakhrani, N., Singh, I. K., Gopalkrishna, K. K., Tourloukis, V., Panagopoulou, E., Karkanas, P., Blickstein, J. I. B., Skinner, A. R., Florentin, J. A., & Harvati, K. (2018). ESR Dating Ungulate Teeth and Molluscs from the Paleolithic Site Marathousa 1, Megalopolis Basin, Greece. Quaternary, 1(3), 22. https://doi.org/10.3390/quat1030022