
Two Thousand Years of Land-Use and Vegetation Evolution in the Andean Highlands of Northern Chile Inferred from Pollen and Charcoal Analyses

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
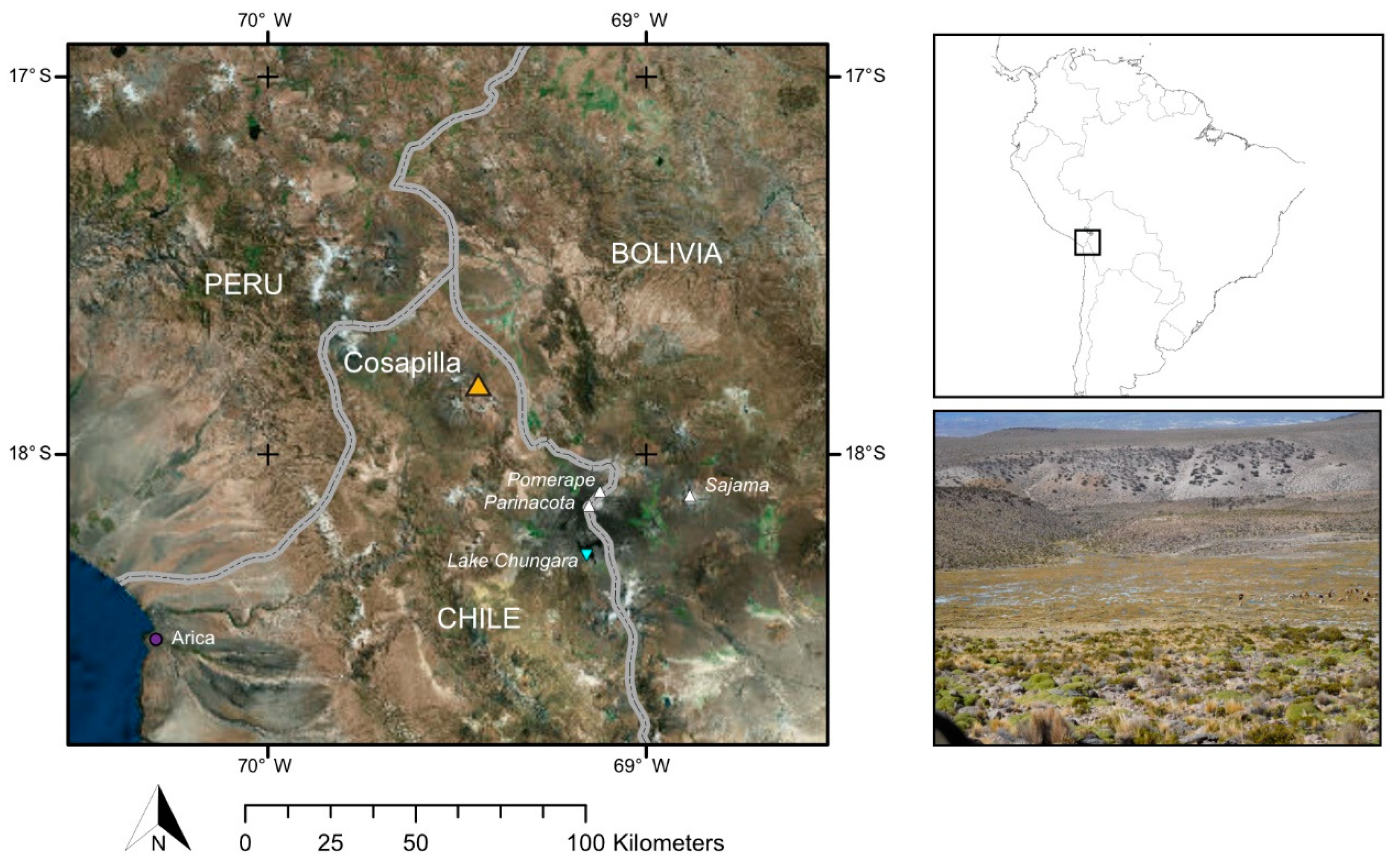
2.1. Study Site
2.2. Sampling and Processing
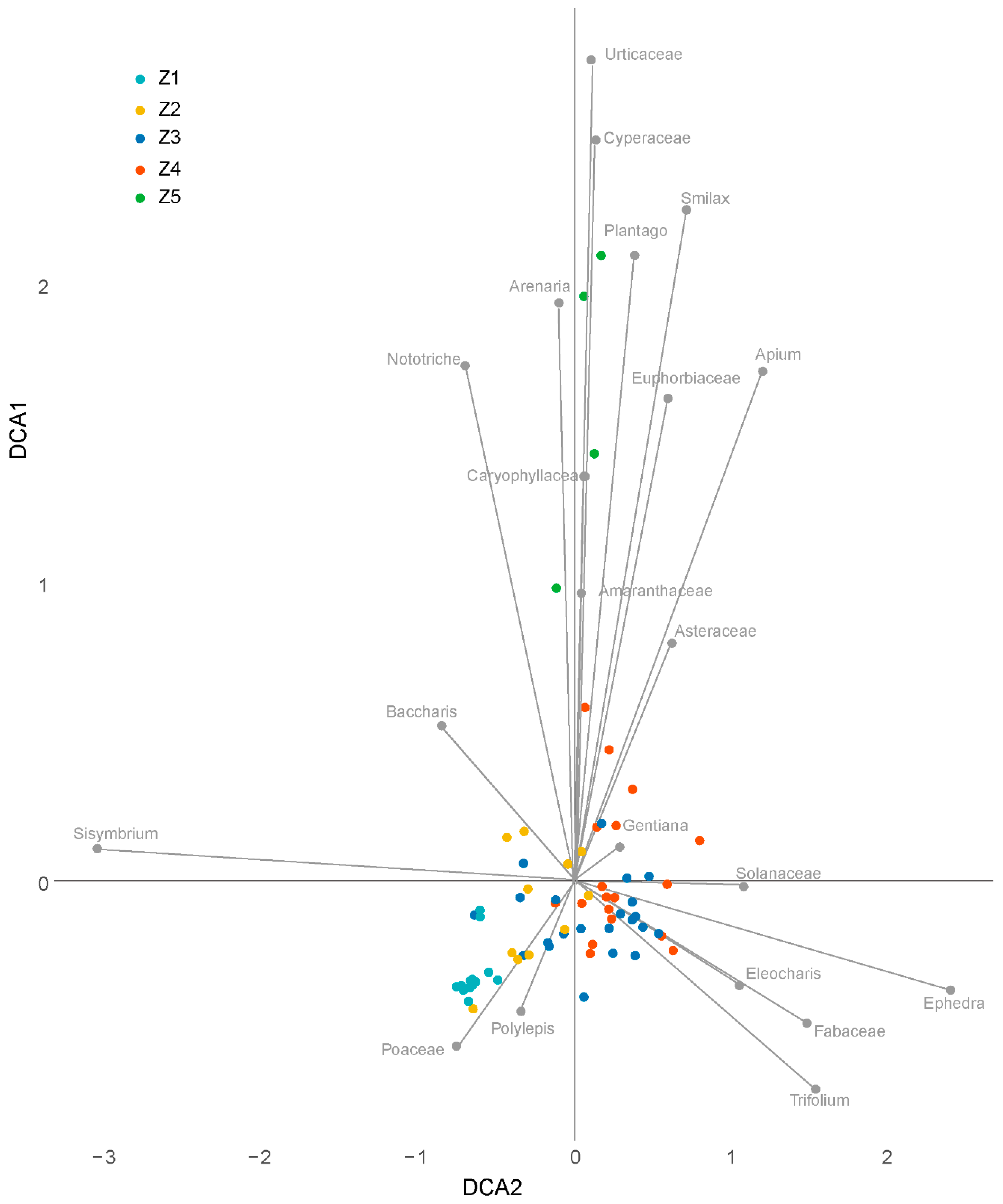
2.3. Statistical Analysis
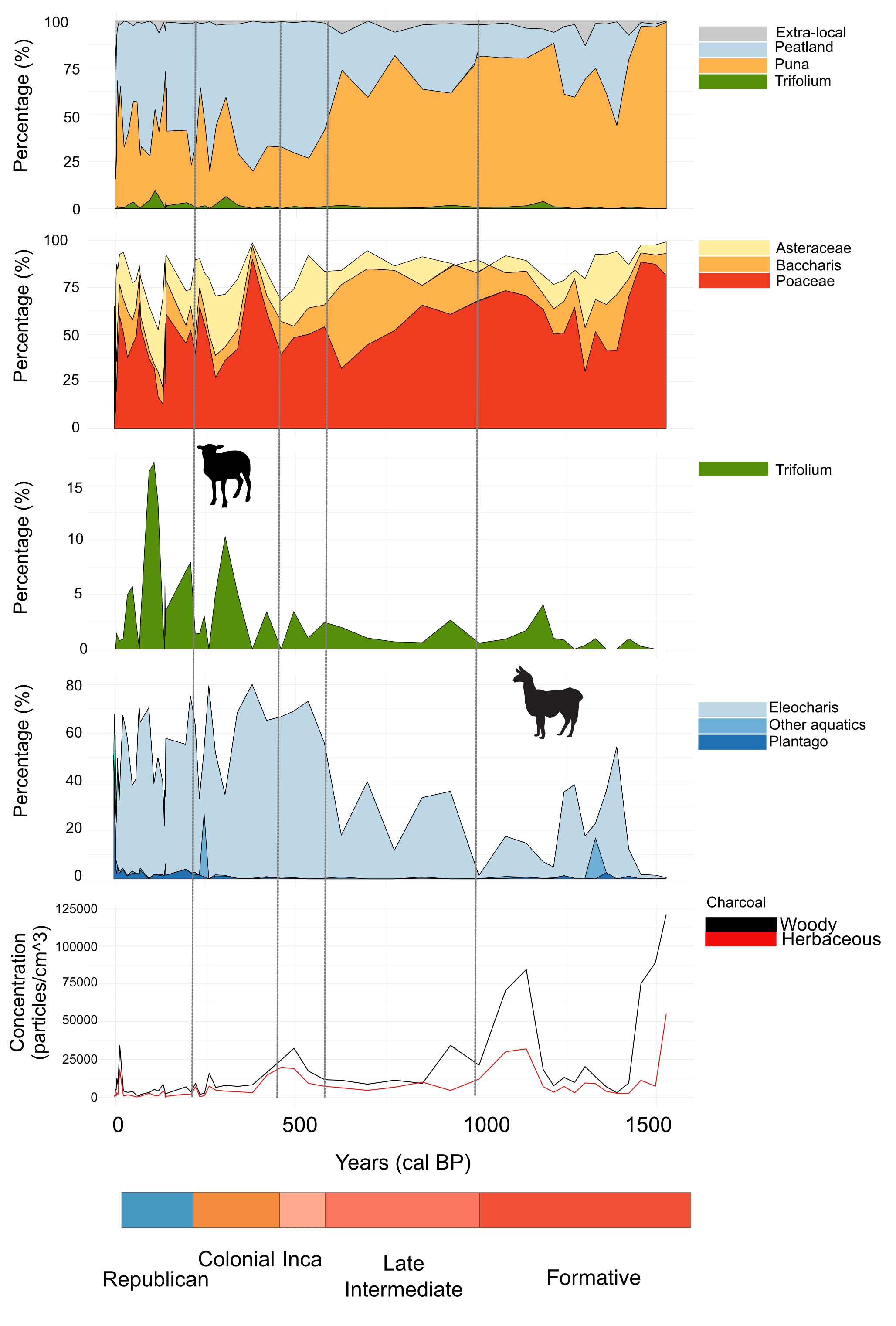
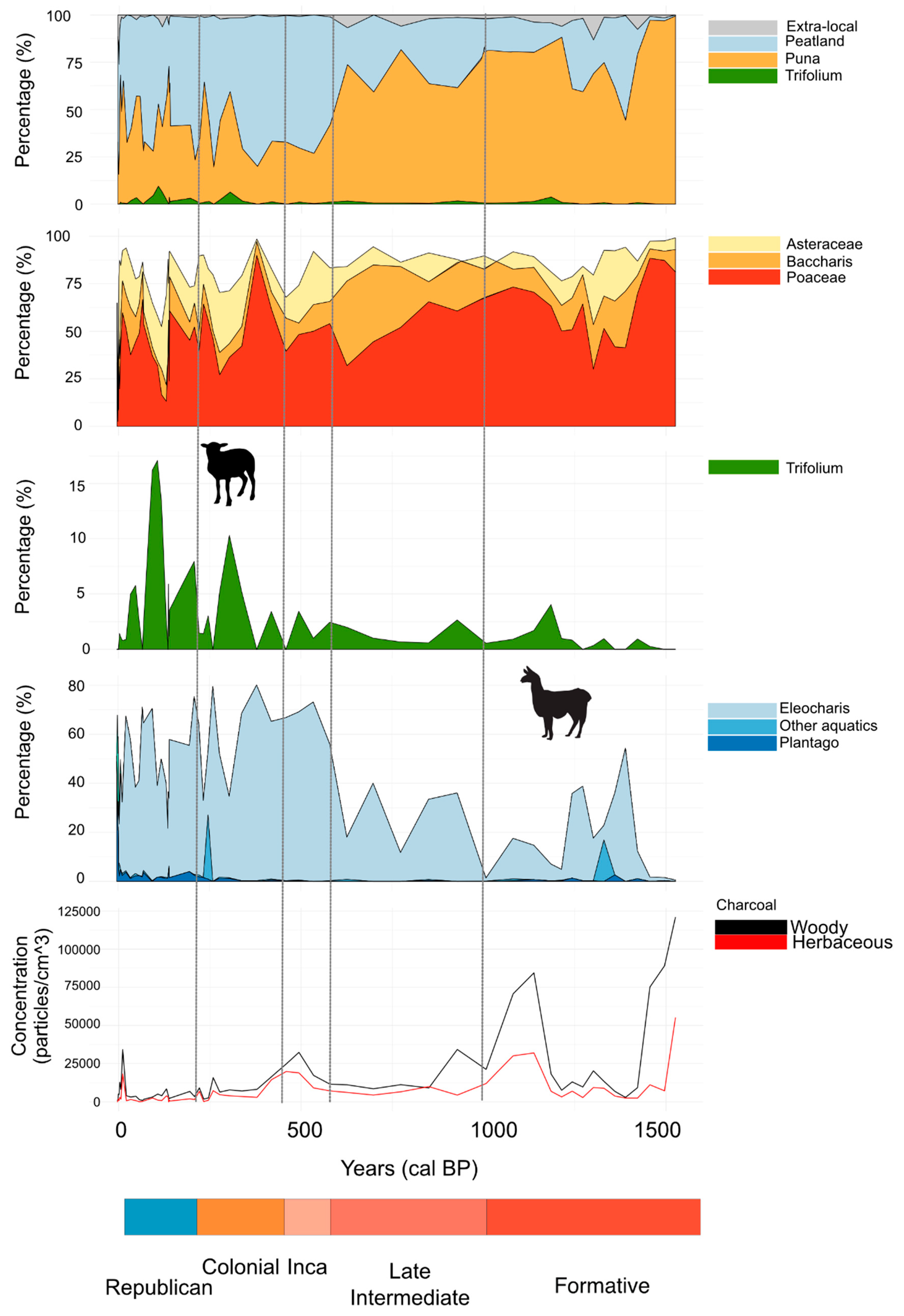
3. Results
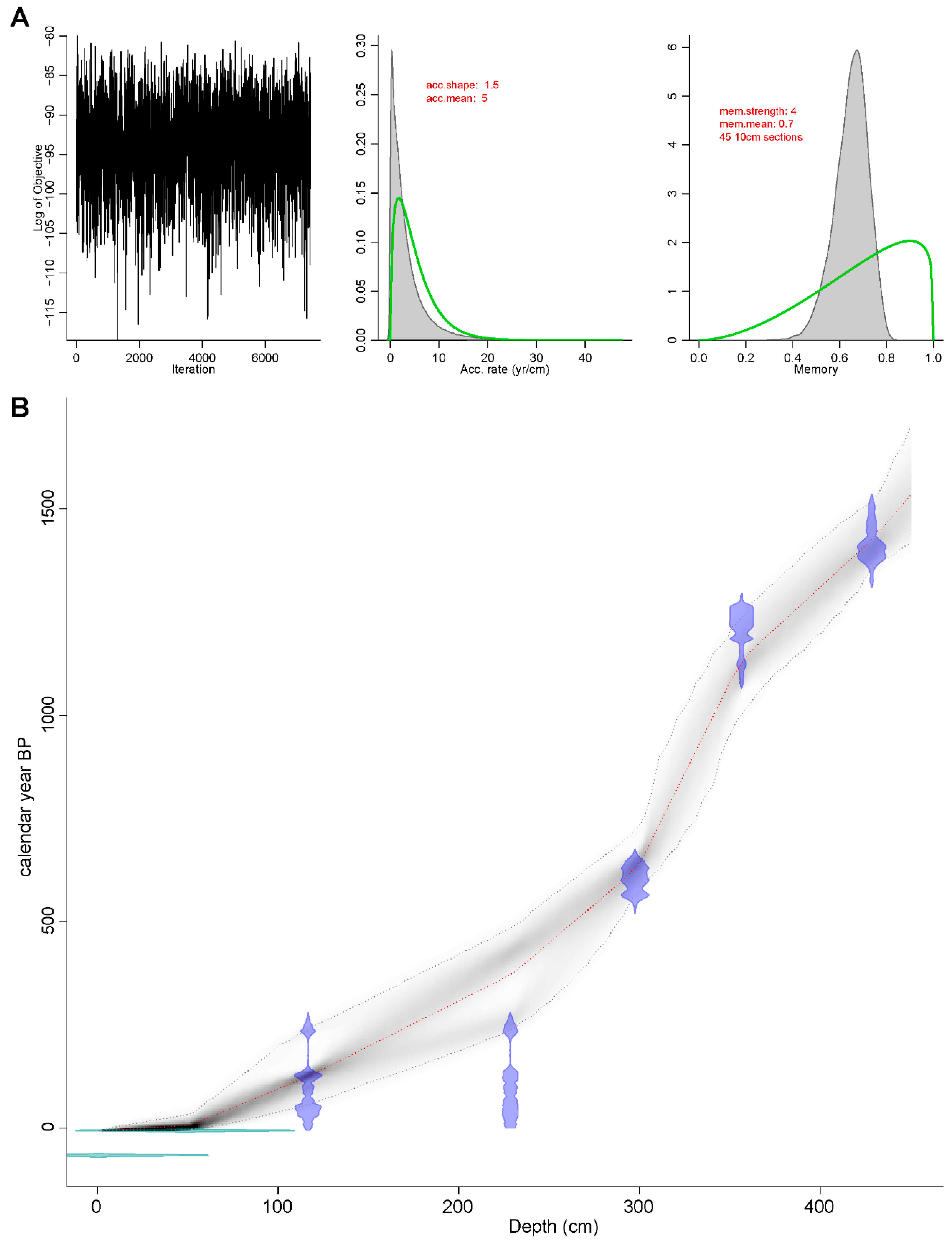
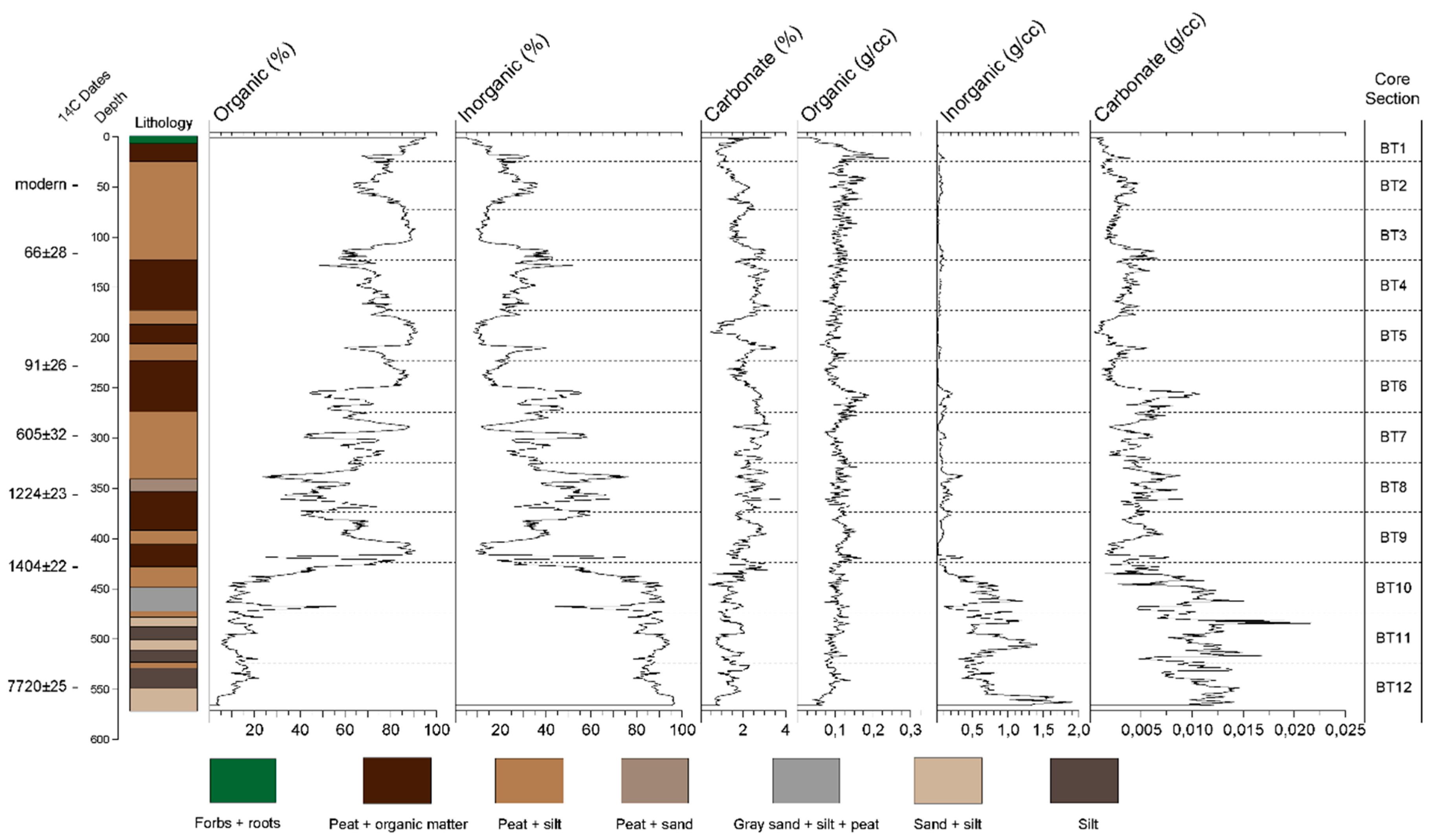
3.1. Chronology and Lithological Description
3.2. Microfossil Analysis
3.2.1. Pollen Zone Z1 (550–435 cm; 7737–1460 calendar year BP)
3.2.2. Pollen Zone Z2 (350–435 cm; 1460–1250 calendar year BP)
3.2.3. Pollen Zone Z3 (150–350 cm; 1250–125 calendar year BP)
3.2.4. Pollen Zone Z4 (30–150 cm; 125–0 calendar year BP)
3.2.5. Pollen Zone Z5 (0–30 cm; 0–4 calendar year BP)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crosby, A.W. The Columbian Exchange: Biological and Cultural Consequences of 1492; Greenwood Publishing Group: Santa Barbara, CA, USA, 2003; ISBN 978-0-275-98073-3. [Google Scholar]
- Denevan, W.M. The pristine myth: The landscape of the Americas in 1492. Ann. Assoc. Am. Geogr. 1992, 82, 369–385. [Google Scholar] [CrossRef]
- Mann, M. The Dark Side of Democracy: Explaining Ethnic Cleansing, 1st ed.; Cambridge University Press: Cambridge, UK, 2004; ISBN 978-0-521-53854-1. [Google Scholar]
- Wheeler, J.C.; Russel, A.J.; Redden, H. Llamas and alpacas: Pre-conquest breeds and post-conquest hybrids. J. Archaeol. Sci. 1995, 22, 833–840. [Google Scholar] [CrossRef]
- Castilla-Beltrán, A.; Hooghiemstra, H.; Hoogland, M.L.P.; Pagán-Jiménez, J.; van Geel, B.; Field, M.H.; Prins, M.; Donders, T.; Herrera Malatesta, E.; Ulloa Hung, J.; et al. Columbus’ footprint in Hispaniola: A paleoenvironmental record of indigenous and colonial impacts on the landscape of the central Cibao Valley, northern Dominican Republic. Anthropocene 2018, 22, 66–80. [Google Scholar] [CrossRef]
- Hooghiemstra, H.; Olijhoek, T.; Hoogland, M.; Prins, M.; van Geel, B.; Donders, T.; Gosling, W.; Hofman, C. Columbus’ environmental impact in the New World: Land use change in the Yaque River valley, Dominican Republic. Holocene 2018, 28, 1818–1835. [Google Scholar] [CrossRef] [PubMed]
- Huber, U.M.; Markgraf, V. European impact on fire regimes and vegetation dynamics at the steppe-forest ecotone of southern Patagonia. Holocene 2003, 13, 567–579. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Villalba, R.; Donnegan, J. Fire history in northern Patagonia: The roles of humans and climatic variation. Ecol. Monogr. 1999, 69, 47–67. [Google Scholar] [CrossRef]
- Rowe, J.H. Culture at the Time of the Spanish Conquest. In Handbook of South American Indians; Smithsonian Institution Press: Washington, DC, USA, 1946; pp. 183–330. [Google Scholar]
- Cook, N.D. Demographic Collapse: Indian Peru, 1520–1620, 1st ed.; Cambridge University Press: Cambridge, UK, 2004; Volume 41. [Google Scholar]
- Santoro, C.M.; Capriles, J.M.; Gayo, E.M.; de Porras, M.E.; Maldonado, A.; Standen, V.G.; Latorre, C.; Castro, V.; Angelo, D.; McRostie, V.; et al. Continuities and discontinuities in the socio-environmental systems of the Atacama Desert during the last 13,000 years. J. Anthropol. Archaeol. 2017, 46, 28–39. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Kolata, A.L. Long-term human response to uncertain environmental conditions in the Andes. Proc. Natl. Acad. Sci. USA 2004, 101, 4325–4330. [Google Scholar] [CrossRef] [Green Version]
- Kennett, D.J.; Marwan, N. Climatic volatility, agricultural uncertainty, and the formation, consolidation and breakdown of preindustrial agrarian states. Philos. Trans. R. Soc. A 2015, 373, 20140458. [Google Scholar] [CrossRef]
- Gade, D.W. Nature and Culture in the Andes; University of Wisconsin Press: Madison, WI, USA, 1999; ISBN 978-0-299-16124-8. [Google Scholar]
- Cooke, C.A.; Balcom, P.H.; Biester, H.; Wolfe, A.P. Over three millennia of mercury pollution in the Peruvian Andes. Proc. Natl. Acad. Sci. USA 2009, 106, 8830–8834. [Google Scholar] [CrossRef] [Green Version]
- Capparelli, A.; Lema, V.; Giovannetti, M.; Raffino, R. The introduction of Old World crops (wheat, barley and peach) in Andean Argentina during the 16th century A.D.: Archaeobotanical and ethnohistorical evidence. Veg. Hist. Archaeobot. 2005, 14, 472–484. [Google Scholar] [CrossRef]
- De France, S.D. Diet and provisioning in the high Andes: a Spanish colonial settlement on the outskirts of Potosí, Bolivia. Int. J. Hist. Archaeol. 2003, 7, 99–125. [Google Scholar] [CrossRef]
- Cade, D.W. Landscape, system, and identity in the post-conquest Andes. Ann. Assoc. Am. Geogr. 1992, 82, 460–477. [Google Scholar] [CrossRef]
- Bush, M.B.; Hanselman, J.A.; Hooghiemstra, H. Andean montane forests and climate change. In Tropical Rainforests Responses to Climate Change; Springer: New York, NY, USA, 2007; pp. 33–54. ISBN 978-3-642-05383-2. [Google Scholar]
- Weng, C.; Bush, M.B.; Curtis, J.H.; Kolata, A.L.; Dillehay, T.D.; Binford, M.W. Deglaciation and Holocene climate change in the western Peruvian Andes. Quat. Res. 2006, 66, 87–96. [Google Scholar] [CrossRef]
- Urrego, D.H.; Silman, M.R.; Correa-Metrio, A.; Bush, M.B. Pollen–vegetation relationships along steep climatic gradients in western Amazonia. J. Veg. Sci. 2011, 22, 795–806. [Google Scholar] [CrossRef]
- Hanselman, J.A.; Gosling, W.D.; Paduano, G.M.; Bush, M.B. Contrasting pollen histories of MIS 5e and the Holocene from Lake Titicaca (Bolivia/Peru). J. Quat. Sci. 2005, 20, 663–670. [Google Scholar] [CrossRef]
- Gosling, W.D.; Hanselman, J.A.; Knox, C.; Valencia, B.G.; Bush, M.B. Long-term drivers of change in Polylepis woodland distribution in the central Andes. J. Veg. Sci. 2009, 20, 1041–1052. [Google Scholar] [CrossRef]
- Capriles, J.M. Mobile communities and pastoralist landscapes during the Formative Period in the Central Altiplano of Bolivia. Lat. Am. Antiq. 2014, 25, 3–26. [Google Scholar] [CrossRef]
- Baker, P.A.; Fritz, S.C. Nature and causes of Quaternary climate variation of tropical South America. Quat. Sci. Rev. 2015, 124, 31–47. [Google Scholar] [CrossRef]
- Hastorf, C.A.; Whitehead, W.T.; Johannessen, S. Late prehistoric wood use in an Andean intermontane valley. Econ. Bot. 2005, 59, 337–355. [Google Scholar] [CrossRef]
- Capriles, J.M. The Economic Organization of Early Camelid Pastoralism in the Andean Highlands of Bolivia; BAR International Series; BAR Publishing: Oxford, UK, 2014; Volume 2597, ISBN 978-1-4073-1229-3. [Google Scholar]
- Vining, B.R.; Steinman, B.A.; Abbott, M.B.; Woods, A. Paleoclimatic and archaeological evidence from Lake Suches for highland Andean refugia during the arid middle-Holocene. Holocene 2018. [Google Scholar] [CrossRef]
- Kessler, M. Present and potential distribution of Polylepis (Rosaceae) forests in Bolivia. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: New York, NY, USA, 1995; pp. 281–294. ISBN 0893273902. [Google Scholar]
- Kessler, M. The “Polylepis problem”: Where do we stand? Ecotropica 2002, 8, 97–110. [Google Scholar]
- Paduano, G.M.; Bush, M.B.; Baker, P.A.; Fritz, S.C.; Seltzer, G.O. A vegetation and fire history of Lake Titicaca since the Last Glacial Maximum. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 194, 259–279. [Google Scholar] [CrossRef] [Green Version]
- Capriles, J.M.; Flores Bedregal, E. The economic, symbolic, and social importance of the “kenua” (Polylepis spp.) during prehispanic times in the Andean highlands of Bolivia. Ecotropica 2002, 8, 225–231. [Google Scholar]
- Baied, C.A.; Wheeler, J.C. Evolution of high Andean puna ecosystems: Environment, climate, and culture change over the last 12,000 years in the Central Andes. Mt. Res. Dev. 1993, 13, 145–156. [Google Scholar] [CrossRef]
- Hanselman, J.A.; Bush, M.B.; Gosling, W.D.; Collins, A.; Knox, C.; Baker, P.A.; Fritz, S.C. A 370,000-year record of vegetation and fire history around Lake Titicaca (Bolivia/Peru). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 305, 201–214. [Google Scholar] [CrossRef]
- Williams, J.J.; Gosling, W.D.; Brooks, S.J.; Coe, A.L.; Xu, S. Vegetation, climate and fire in the eastern Andes (Bolivia) during the last 18,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 312, 115–126. [Google Scholar] [CrossRef]
- Reese, C.A.; Liu, K.B.; Thompson, L.G. An ice-core pollen record showing vegetation response to Late-glacial and Holocene climate changes at Nevado Sajama, Bolivia. Ann. Glaciol. 2013, 54, 183–190. [Google Scholar] [CrossRef]
- Kuentz, A.; Ledru, M.-P.; Thouret, J.-C. Environmental changes in the highlands of the western Andean Cordillera, southern Peru, during the Holocene. Holocene 2011, 22, 1215–1216. [Google Scholar] [CrossRef]
- Rundel, P.W.; Palma, B. Preserving the unique puna ecosystems of the Andean Altiplano. Mt. Res. Dev. 2000, 20, 262–271. [Google Scholar] [CrossRef]
- Beck, S.G.; Domic, A.I.; Garcia, C.; Meneses, R.I.; Yager, K.; Halloy, S. El Parque Nacional Sajama y sus Plantas; Herbario Nacional de Bolivia, Fundacion PUMA: La Paz, Bolivia, 2010; ISBN 978-99905-48-01-3. [Google Scholar]
- Ruthsatz, B. Vegetation and ecology of the high Andean peatlands of Bolivia. Phytocoenologia 2012, 42, 133–179. [Google Scholar] [CrossRef]
- Meneses, R.I.; Domic, A.I.; Beck, S.G.; Yager, K. Bofedales Altoandinos: Oasis en La Puna; Herbario Nacional de Bolivia: La Paz, Bolivia, 2018. [Google Scholar]
- Squeo, F.A.; Warner, B.G.; Aravena, R.; Espinoza, D. Bofedales: High altitude peatlands of the central Andes. Rev. Chil. Hist. Nat. 2006, 79, 245–255. [Google Scholar] [CrossRef]
- Pfister, J.A.; Martin, F.S.; Rosales, L.; Sisson, D.V.; Flores, E.; Bryant, F.C. Grazing behavior of llamas, alpacas and sheep in the Andes of Peru. Appl. Anim. Behav. Sci. 1989, 23, 237–246. [Google Scholar] [CrossRef]
- Bengtsson, L.; Enell, M. Chemical analysis. In Handbook of Holocene Palaeoecology and Palaeohydrology; John Wiley: Chichester, UK, 1986; pp. 423–451. ISBN 1930665806. [Google Scholar]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; John Wiley: Chichester, UK, 1989. [Google Scholar]
- Roubik, D.W.; Moreno, E. Pollen and Spores of Barro Colorado Island; Missouri Botanical Garden: Saint Louis, MO, USA, 1981. [Google Scholar]
- Sandoval, A.P.; Marconi, L.; Ortuño, T. Flora Polínica de Bofedales y Zonas Aledañas del Tuni Condoriri; Herbario Nacional de Bolivia: La Paz, Bolivia, 2011. [Google Scholar]
- Bush, M.B.; Weng, C. Introducing a new (freeware) tool for palynology. J. Biogeogr. 2007, 34, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Collao-Alvarado, K.; Maldonado, A.; González, L.; Sandoval, A.; De Porras, M.E.; Zamora, A.; Arancio, G. Estudio de la relación polen-vegetación actual en el Norte de Chile, en el transecto Pozo Almonte-Salar de Huasco (20°15′S/69°06′O). Gayana Botánica 2018, 72, 0717–6643. [Google Scholar] [CrossRef]
- Walsh, M.K.; Prufer, K.M.; Culleton, B.J.; Kennett, D.J. A late Holocene paleoenvironmental reconstruction from Agua Caliente, southern Belize, linked to regional climate variability and cultural change at the Maya polity of Uxbenká. Quat. Res. 2014, 82, 38–50. [Google Scholar] [CrossRef]
- Hogg, A.G.; Hua, Q.; Blackwell, P.G.; Niu, M.; Buck, C.E.; Guilderson, T.P.; Heaton, T.J.; Palmer, J.G.; Reimer, P.J.; Reimer, R.W.; et al. SHCal13 Southern Hemisphere Calibration, 0–50,000 years cal BP. Radiocarbon 2013, 55, 1889–1903. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, A.J. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- De Porras, M.E.; Maldonado, A.; De Pol-Holz, R.; Betancourt, J.L. Late Quaternary environmental dynamics in the Atacama Desert reconstructed from rodent midden pollen records. J. Quat. Sci. 2017, 32, 665–684. [Google Scholar] [CrossRef]
- Grimm, E.C. Tilia: Version 1.5.12; Illinois State Museum: Springfield, MA, USA, 2011. [Google Scholar]
- Wirrmann, D.; De Oliveira Almeida, L.F. Low Holocene level (7700 to 3650 years ago) of Lake Titicaca (Bolivia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1987, 59, 315–323. [Google Scholar] [CrossRef]
- Tapia, P.M.; Fritz, S.C.; Baker, P.A.; Seltzer, G.O.; Dunbar, R.B. A Late Quaternary diatom record of tropical climatic history from Lake Titicaca (Peru and Bolivia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 194, 139–164. [Google Scholar] [CrossRef] [Green Version]
- Pueyo, J.J.; Sáez, A.; Giralt, S.; Valero-Garcés, B.L.; Moreno, A.; Bao, R.; Schwalb, A.; Herrera, C.; Klosowska, B.; Taberner, C. Carbonate and organic matter sedimentation and isotopic signatures in Lake Chungará, Chilean Altiplano, during the last 12.3 kyr. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 307, 339–355. [Google Scholar] [CrossRef]
- Ledru, M.-P.; Jomelli, V.; Samaniego, P.; Vuille, M.; Hidalgo, S.; Herrera, M.; Ceron, C. The Medieval Climate Anomaly and the Little Ice Age in the eastern Ecuadorian Andes. Clim. Past 2013, 9, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Tchilinguirian, P.; Morales, M.R. Mid-Holocene paleoenvironments in northwestern Argentina: Main patterns and discrepancies. Quat. Int. 2013, 307, 14–23. [Google Scholar] [CrossRef]
- Moreno, A.; Giralt, S.; Valero-Garcés, B.; Sáez, A.; Bao, R.; Prego, R.; Pueyo, J.J.; González-Sampériz, P.; Taberner, C. A 14 kyr record of the tropical Andes: The Lago Chungará sequence (18° S, northern Chilean Altiplano). Quat. Int. 2007, 161, 4–21. [Google Scholar] [CrossRef]
- Grosjean, M.; Núñez, A.L. Lateglacial, early and middle Holocene environments, human occupation, and resource use in the Atacama (Northern Chile). Geoarchaeology 1994, 9, 271–286. [Google Scholar] [CrossRef]
- Grosjean, M.; Geyh, M.A.; Messerli, B.; Schotterer, U. Late-glacial and early Holocene lake sediments, ground-water formation and climate in the Atacama Altiplano 22–24° S. J. Paleolimnol. 1995, 14, 241–252. [Google Scholar] [CrossRef]
- Grosjean, M.; Núñez, L.; Cartajena, I.; Messerli, B. Mid-Holocene climate and culture change in the Atacama desert, Northern Chile. Quat. Res. 1997, 48, 239–246. [Google Scholar] [CrossRef]
- Grosjean, M.; van Leeuwen, J.F.N.; van der Knaap, W.O.; Geyh, M.A.; Ammann, B.; Tanner, W.; Messerli, B.; Núñez, L.A.; Valero-Garcés, B.L.; Veit, H. A 22,000 14C year BP sediment and pollen record of climate change from Laguna Miscanti (23° S), northern Chile. Glob. Planet. Chang. 2001, 28, 35–51. [Google Scholar] [CrossRef]
- Núñez, L.; Loyola, R.; Cartajena, I.; López, P.; Santander, B.; Maldonado, A.; de Souza, P.; Carrasco, C. Miscanti-1: Human occupation during the arid Mid-Holocene event in the high-altitude lakes of the Atacama Desert, South America. Quat. Sci. Rev. 2018, 181, 109–122. [Google Scholar] [CrossRef]
- Grosjean, M.; Valero-Garcés, B.L.; Geyh, M.A.; Messerli, B.; Schotterer, U.; Schreier, H.; Kelts, K. Mid- and late-Holocene limnogeology of Laguna del Negro Francisco, northern Chile, and its palaeoclimatic implications. Holocene 1997, 7, 151–159. [Google Scholar] [CrossRef]
- Grosjean, M.; Geyh, M.A.; Messerli, B.; Schreier, H.; Veit, H. A late-Holocene (<2600 BP) glacial advance in the south-central Andes (29° S), northern Chile. Holocene 1998, 8, 473–479. [Google Scholar] [CrossRef]
- Latorre, C.; Betancourt, J.L.; Rylander, K.A.; Quade, J.; Matthei, O. A vegetation history from the arid prepuna of northern Chile (22–23° S) over the last 13,500 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 194, 223–246. [Google Scholar] [CrossRef]
- Latorre, C.; Betancourt, J.L.; Arroyo, M.T.K. Late Quaternary vegetation and climate history of a perennial river canyon in the Río Salado basin (22° S) of northern Chile. Quat. Res. 2006, 65, 450–466. [Google Scholar] [CrossRef]
- Mujica, M.I.; Latorre, C.; Maldonado, A.; González-Silvestre, L.; Pinto, R.; de Pol-Holz, R.; Santoro, C.M. Late Quaternary climate change, relict populations and present-day refugia in the northern Atacama Desert: A case study from Quebrada La Higuera (18° S). J. Biogeogr. 2015, 42, 76–88. [Google Scholar] [CrossRef]
- Valero-Garcés, B.; Grosjean, M.; Schwalb, A.; Geyh, M.; Messerli, B.; Kelts, K. Limnogeology of Laguna Miscanti: Evidence for mid to late Holocene moisture changes in the Atacama Altiplano (Northern Chile). J. Paleolimnol. 1996, 16, 1–21. [Google Scholar] [CrossRef]
- Schittek, K.; Kock, S.T.; Lücke, A.; Hense, J.; Ohlendorf, C.; Kulemeyer, J.J.; Lupo, L.C.; Schäbitz, F. A high-altitude peatland record of environmental changes in the NW Argentine Andes (24° S) over the last 2100 years. Clim. Past 2016, 12, 1165–1180. [Google Scholar] [CrossRef]
- Thompson, L.G.; Davis, M.E.; Mosley-Thompson, E.; Sowers, T.A.; Henderson, K.A.; Zagorodnov, V.S.; Lin, P.-N.; Mikhalenko, V.N.; Campen, R.K.; Bolzan, J.F.; et al. A 25,000-year tropical climate history from Bolivian ice cores. Science 1998, 282, 1858–1864. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.G.; Mosley-Thompson, E.; Bolzan, J.F.; Koci, B.R. A 1500-year record of tropical precipitation in ice cores from the Quelccaya ice cap, Peru. Science 1985, 229, 971–973. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.G.; Mosley-Thompson, E.; Dansgaard, W.; Grootes, P.M. The Little Ice Age as recorded in the stratigraphy of the tropical Quelccaya ice cap. Science 1986, 234, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.S.; Christie, D.A.; Villalba, R.; Argollo, J.; Pacajes, J.; Silva, J.S.; Alvarez, C.A.; Llancabure, J.C.; Gamboa, C.S. Precipitation changes in the South American Altiplano since 1300 AD reconstructed by tree-rings. Clim. Past 2012, 8, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Reese, C.A.; Thompson, L.G. Ice-core pollen record of climatic changes in the central Andes during the last 400 yr. Quat. Res. 2005, 64, 272–278. [Google Scholar] [CrossRef]
- Baied, C.A. Distribución actual de matorrales altoandinos de Polylepis en los Andes Centro-Sur: Oscilaciones climáticas y el impacto de la actividad humana en el pasado. In En los Tres Reinos: Prácticas de Recolección en el cono sur de América; Universidad Nacional de Tucumán: Tucumán, Argentina, 1999; pp. 129–139. [Google Scholar]
- Rodríguez, F.; Behling, H. Late Quaternary vegetation, climate and fire dynamics, and evidence of early to mid-Holocene Polylepis forests in the Jimbura region of the southernmost Ecuadorian Andes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 247–257. [Google Scholar] [CrossRef]
- Moreno, A.; Santoro, C.M.; Latorre, C. Climate change and human occupation in the northernmost Chilean Altiplano over the last ca. 11 500 cal. a BP. J. Quat. Sci. 2008, 24, 373–382. [Google Scholar] [CrossRef]
- Allen, J.R.M.; Huntley, B. Effects of tephra falls on vegetation: A Late-Quaternary record from southern Italy. J. Ecol. 2018, 106, 2456–2472. [Google Scholar] [CrossRef]
- Santoro, C.M.; Núñez, L. Hunters of the dry Puna and the salt Puna in northern Chile. Andean Past 1987, 1, 57–109. [Google Scholar]
- Osorio, D.; Steele, J.; Sepúlveda, M.; Gayo, E.M.; Capriles, J.M.; Herrera, K.; Ugalde, P.; De Pol-Holz, R.; Latorre, C.; Santoro, C.M. The dry Puna as an ecological megapatch and the peopling of South America: Technology, mobility, and the development of a late Pleistocene/early Holocene Andean hunter-gatherer tradition in northern Chile. Quat. Int. 2017, 461, 41–53. [Google Scholar] [CrossRef]
- Osorio, D.; Jackson, D.; Ugalde, P.C.; Latorre, C.; De Pol-Holz, R.; Santoro, C.M. Hakenasa Cave and its relevance for the peopling of the southern Andean Altiplano. Antiquity 2011, 85, 1194–1208. [Google Scholar] [CrossRef]
- Núñez, L.; Cartajena, I.; Grosjean, M. Archaeological silence and ecorefuges: Arid events in the Puna of Atacama during the middle Holocene. Quat. Int. 2013, 307, 5–13. [Google Scholar] [CrossRef]
- Vining, B.R. Pastoral intensification, social fissioning, and ties to state economies at the formative period–middle horizon transition in the Lake Suches region, southern Peru. In The Archaeology of Andean Pastoralism; University of New Mexico Press: Alburquerque, NM, USA, 2016; ISBN 978-0-8263-5703-8. [Google Scholar]
- Aragón, R.; Carilla, J.; Cristóbal, L. Fire, plant species richness and aerial biomass distribution in mountain grasslands of NW Argentina. In Land Use Change and Mountain Biodiversity; Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 89–99. ISBN 9780849335235. [Google Scholar]
- Carilla, J.; Aragón, R.; Gurvich, D.E. Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands. Acta Oecol. 2011, 37, 337–345. [Google Scholar] [CrossRef]
- Patty Aguilar, L.B. Effects of Grazing and Fire on Herbaceous Species in the Bolivian Altiplano. Ph.D. Thesis, University of Basel, Basel, Switzerland, 2012. [Google Scholar]
- Verzijl, A.; Quispe, S.G. The system nobody sees: Irrigated wetland management and alpaca herding in the Peruvian Andes. Mt. Res. Dev. 2013, 33, 280–293. [Google Scholar] [CrossRef]
- Lane, K.; Grant, J. A question of altitude: Exploring the limits of highland pastoralism in the prehispanic Andes. In The Archaeology of Andean Pastoralism; University of New Mexico Press: Alburquerque, NM, USA, 2016; pp. 139–157. [Google Scholar]
- Castellaro, G.; Squella, F.; León, F.; Raggi, A. Composición botánica de la dieta de alpaca (Lama pacos Linn.) en un pastizal del secano mediterráneo de la zona central de Chile. Chil. J. Agric. Res. 2008, 68, 136–145. [Google Scholar] [CrossRef]
- Borgnia, M.; Vilá, B.L.; Cassini, M.H. Interaction between wild camelids and livestock in an Andean semi-desert. J. Arid Environ. 2008, 72, 2150–2158. [Google Scholar] [CrossRef]
- Machaca, N.C.; Condori, B.; Pardo, A.R.; Anthelme, F.; Meneses, R.I.; Weeda, C.E.; Perotto-Baldivieso, H.L. Effects of grazing pressure on plant species composition and water presence on bofedales in the Andes mountain range of Bolivia. Mires Peat 2018, 21, 1–15. [Google Scholar]
- Villarroel, E.K.; Pacheco Mollinedo, P.L.; Domic, A.I.; Capriles, J.M.; Espinoza, C. Local management of Andean wetlands in Sajama National Park, Bolivia. Mt. Res. Dev. 2014, 34, 356–368. [Google Scholar] [CrossRef]
- López-i-Gelats, F.; Contreras Paco, J.L.; Huilcas Huayra, R.; Siguas Robles, O.D.; Quispe Peña, E.C.; Bartolomé Filella, J. Adaptation strategies of Andean pastoralist households to both climate and non-climate changes. Hum. Ecol. 2015, 43, 267–282. [Google Scholar] [CrossRef]
- Bruno, M.C.; Hastorf, C.A. Gifts from the camelids: Archaeobotanical insights into camelid pastoralism through the study of dung. In The Archaeology of Andean Pastoralism; University of New Mexico Press: Alburquerque, NM, USA, 2016; pp. 55–65. ISBN 978-0-8263-5703-8. [Google Scholar]
- Markemann, A.; Valle Zárate, A. Traditional llama husbandry and breeding management in the Ayopaya region, Bolivia. Trop. Anim. Health Prod. 2009, 42, 79. [Google Scholar] [CrossRef]
- Villagrán, M.C.; Castro, R.V. Etnobotánica y manejo ganadero de las vegas, bofedales y quebradas en el Loa Superior, Andes de Antofagasta, Segunda Región, Chile. Chungara 1997, 29, 275–304. [Google Scholar]
- Chimmer, R.A.; Bonvissuto, G.L.; Cremona, M.V.; Gaitan, J.J.; López, C.R. Ecohydrological conditions of wetlands along a precipitation gradient in Patagonia, Argentina. Ecol. Aust. 2011, 21, 329–337. [Google Scholar]
- Flantua, S.; Hooghiemstra, H.; Vuille, M.; Behling, H.; Carson, J.F.; Gosling, W.D.; Hoyos, I.; Ledru, M.P.; Montoya, E.; Mayle, F.; et al. Climate variability and human impact in South America during the last 2000 years: Synthesis and perspectives from pollen records. Clim. Past 2016, 12, 483–523. [Google Scholar] [CrossRef]
- Loughlin, N.J.D.; Gosling, W.D.; Coe, A.L.; Gulliver, P.; Mothes, P.; Montoya, E. Landscape-scale drivers of glacial ecosystem change in the montane forests of the eastern Andean flank, Ecuador. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 489, 198–208. [Google Scholar] [CrossRef]
- Rozas, V.; Le Quesne, C.; Rojas-Badilla, M.; González, M.E.; González-Reyes, A. Coupled human-climate signals on the fire history of upper Cachapoal Valley, Mediterranean Andes of Chile, since 1201 CE. Glob. Planet. Chang. 2018, 167, 137–147. [Google Scholar] [CrossRef]
- Dobyns, H.F. An outline of Andean epidemic history to 1720. Bull. Hist. Med. 1963, 37, 493–515. [Google Scholar] [PubMed]
- Brading, D.A.; Cross, H.E. Colonial silver mining: Mexico and Peru. Hisp. Am. Hist. Rev. 1972, 52, 545–579. [Google Scholar] [CrossRef]
- Newson, L.A. The demographic collapse of native peoples of the Americas, 1492–1650. Proc. Br. Acad. 1993, 81, 247–288. [Google Scholar]
- Lima, M.; Christie, D.A.; Santoro, M.C.; Latorre, C. Coupled socio-environmental changes triggered indigenous Aymara depopulation of the semiarid Andes of Tarapacá-Chile during the late 19th–20th centuries. PLoS ONE 2016, 11, e0160580. [Google Scholar] [CrossRef]
- Van Kessel, J. Holocausto al Progreso: Los Aymarás de Tarapacá, 1st ed.; IECTA: Iquique, Chile, 1992; ISBN 9070280523. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab Code | Core Depth (cm) | 14C Years BP | Error | Calibrated Age (Calendar Year BP) |
|---|---|---|---|---|
| D-AMS-028556 | 48.5 | modern | --- | |
| D-AMS-028557 | 116.5 | 87 | 28 | 66 ± 28 |
| D-AMS-028558 | 228.5 | 129 | 26 | 91 ± 26 |
| D-AMS-028559 | 297.5 | 665 | 32 | 605 ± 32 |
| D-AMS-028560 | 356.5 | 1316 | 23 | 1224 ± 23 |
| D-AMS-028561 | 428.5 | 1571 | 22 | 1404 ± 22 |
| D-AMS-028562 | 550 | 6912 | 25 | 7702 ± 38 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domic, A.I.; Capriles, J.M.; Escobar-Torrez, K.; Santoro, C.M.; Maldonado, A. Two Thousand Years of Land-Use and Vegetation Evolution in the Andean Highlands of Northern Chile Inferred from Pollen and Charcoal Analyses. Quaternary 2018, 1, 32. https://doi.org/10.3390/quat1030032
Domic AI, Capriles JM, Escobar-Torrez K, Santoro CM, Maldonado A. Two Thousand Years of Land-Use and Vegetation Evolution in the Andean Highlands of Northern Chile Inferred from Pollen and Charcoal Analyses. Quaternary. 2018; 1(3):32. https://doi.org/10.3390/quat1030032
Chicago/Turabian StyleDomic, Alejandra I., José M. Capriles, Katerine Escobar-Torrez, Calogero M. Santoro, and Antonio Maldonado. 2018. "Two Thousand Years of Land-Use and Vegetation Evolution in the Andean Highlands of Northern Chile Inferred from Pollen and Charcoal Analyses" Quaternary 1, no. 3: 32. https://doi.org/10.3390/quat1030032
APA StyleDomic, A. I., Capriles, J. M., Escobar-Torrez, K., Santoro, C. M., & Maldonado, A. (2018). Two Thousand Years of Land-Use and Vegetation Evolution in the Andean Highlands of Northern Chile Inferred from Pollen and Charcoal Analyses. Quaternary, 1(3), 32. https://doi.org/10.3390/quat1030032