Multicore Study of Upper Holocene Mire Development in West-Frisia, Northern Netherlands: Ecological and Archaeological Aspects

 ,
, 
Abstract
:1. Introduction
1.1. The Abundance of Trees in the West-Frisian Landscape during the Bronze Age
1.2. The Role of Climate Change in Driving Human and Vegetation Dynamics in West-Frisia
1.3. Late Medieval Degredation of the West-Frisian Landscape
1.4. Research Questions
2. Materials and Methods
3. Results and Interpretation
3.1. Beets (BTS, Diagram in ESM2)
3.1.1. Zone BTS-1 (50–9 cm Depth)
3.1.2. Zone BTS-2 (9–1 cm Depth)
3.2. Berkhout (BKH, Diagram in ESM2)
3.2.1. Zone BKH-1 (49–44 cm Depth)
3.2.2. Zone BKH-2 (44–40 cm Depth)
3.2.3. Zone BKH-3 (40–4 cm Depth)
| BKH-3A (40–37 cm Depth) | Mineral-rich fen with C. giganteum. |
| BKH-3B (37–31 cm Depth) | Mineral-rich fen with T. nitens, although the raised bog fungi cf. Entophlyctis lobata (HdV-13) and HdV-65 already indicate development towards mineral-poor conditions. |
| BKH-3C (31–21 cm Depth) | Mineral-rich fen with C. giganteum and a maximum of HdV-123. |
| BKH-3D (21–10 cm Depth) | Mineral-rich fen with Tomentypnum nitens. HdV-13, HdV-65, Amphitrema flavum and the presence of Paludella squarrosa already point to local acidification. Paludella squarrrosa occurs in areas with a relatively high input of groundwater [56] but is especially common in mineral-poor fens. |
| BKH-3E (10–4 cm Depth) | Mineral-rich fen with HdV-123 and C. giganteum. Some temporary disturbance (flooding with water, enriched in minerals) is evidenced by the presence of Mougeotia, Spirogyra, Zygnema-type and HdV-128A in the sample at 5 cm depth. |
3.2.4. Zone BKH-4 (4–1 cm Depth)
3.3. Bovenkarspel (BVK, Diagram in ESM2)
3.3.1. Zone BVK-1 (48–41 cm Depth)
3.3.2. Zone BVK-2 (41–35 cm Depth)
3.3.3. Zone BVK-3 (35–25 cm Depth)
3.3.4. Zone BVK-4 (25–1 cm Depth)
3.4. Enkhuizen-Lange Tuinstraat (ELT, Diagram in ESM2)
3.4.1. Zone ELT-1 (85–63 cm Depth)
3.4.2. Zone ELT-2 (63–51 cm Depth)
3.4.3. Zone ELT-3 (51–27 cm Depth)
3.4.4. Zone ELT-4 (27–1 cm Depth)
3.5. Enkhuizen-Paktuinen (EPK, Diagram in ESM2)
3.5.1. Zone EPK-1 (88–69 cm Depth)
3.5.2. Zone EPK-2 (69–59 cm Depth)
3.5.3. Zone EPK-3 (59–30 cm Depth)
3.5.4. Zone EPK-4 (30–23 cm Depth)
3.5.5. Zone EPK-5 (23–1 cm Depth)
3.6. Enkhuizen-Raamstraat (ERS, Diagram in ESM2)
3.6.1. Zone ERS-1 (100–73 cm Depth)
3.6.2. Zone ERS-2 (73–61 cm Depth)
3.6.3. Zone ERS-3 (61–29 cm Depth)
3.6.4. Zone ERS-4 (29–7 cm Depth)
3.6.5. Zone ERS-5 (7–1 cm Depth)
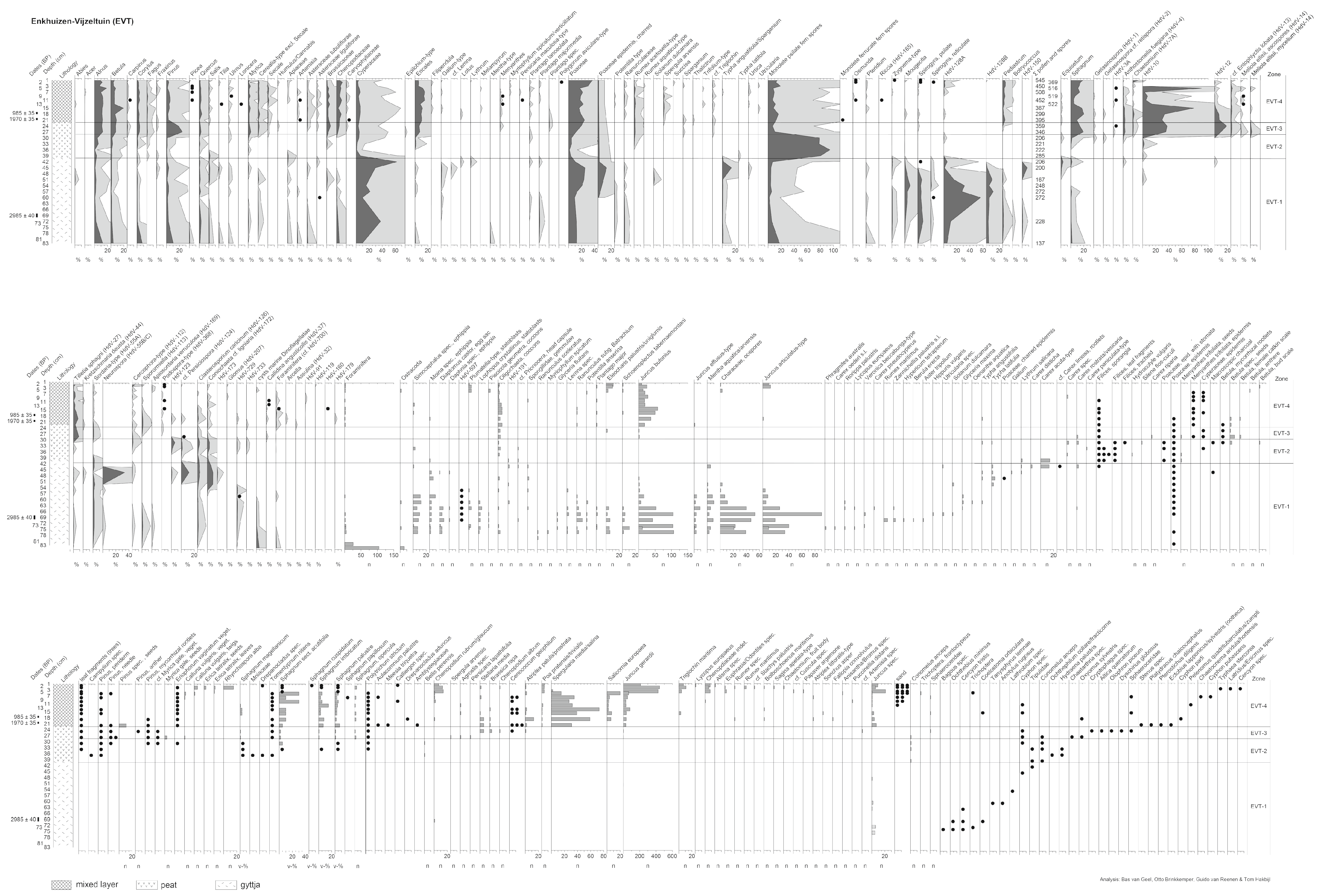
3.7. Enkhuizen-Vijzeltuin (EVT)
3.7.1. Zone EVT-1 (83–40 cm Depth)
3.7.2. Zone EVT-2 (40–28 cm Depth)
3.7.3. Zone EVT-3 (28–22 cm Depth)
3.7.4. Zone EVT-4 (22–1 cm Depth)
3.8. Enkhuizen-Westeinde (EWE, Diagram in ESM3)
3.8.1. Zone EWE-1 (46–26 cm Depth)
3.8.2. Zone EWE-2 (26–20 cm Depth)
3.8.3. Zone EWE-3 (20–13 cm Depth)
3.8.4. Zone EWE-4 (13–3 cm Depth)
3.9. Grootebroek-Zesstedenweg (GBZ, Diagram in ESM3)
3.9.1. Zone GBZ-1 (113–95 cm Depth)
3.9.2. Zone GBZ-2 (95–75 cm Depth)
3.9.3. Zone GBZ-3 (75–49 cm Depth; the Interval between 65 and 49 cm Depth Was not Sampled)
3.9.4. Zone GBZ-4 (49–39 cm Depth)
3.9.5. Zone GBZ-5 (39–1 cm Depth)
3.10. Oostwoud (OWD, Diagram in ESM3)
3.10.1. Zone OWD-1 (86–66 cm Depth)
3.10.2. Zone OWD-2 (66–60 cm Depth)
3.10.3. Zone OWD-3 (60–51 cm Depth)
3.10.4. Zone OWD-4 (51–27 cm Depth)
3.10.5. Zone OWD-5 (27–22 cm Depth)
3.11. Venhuizen-Elbaweg (VEL) and Venhuizen-Spuiter (VSP)
3.12. Venhuizen-Elbaweg (VEL, Diagram in ESM3)
3.12.1. Zone VEL-1 (99–74 cm Depth)
3.12.2. Zone VEL-2 (74–52 cm Depth)
3.12.3. Zone VEL-3 (52–19 cm Depth)
3.12.4. Zone VEL-4 (19–16 cm Depth)
3.12.5. Zone VEL-5 (16–5 cm Depth)
3.13. Venhuizen-Spuiter (VSP, Diagram in ESM3)
3.13.1. Zone VSP-1 (43–30 cm Depth)
3.13.2. Zone VSP-2 (30–21 cm Depth)
3.13.3. Zone VSP-3 (21–1 cm Depth)
4. Discussion
4.1. Forest in West-Frisia during Late Bronze Age? (Research Question 1)
4.2. Rise of Groundwater and Climate Change (Research Question 2)
4.3. Medieval Start of Peat Land Reclamation and Settlements (Research Question 3)
4.4. Human Impact and Increasing Marine Influence in West-Frisia
4.5. Forest and Toponyms
4.6. Relevance for Management of Wetlands (Research Question 4)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Some Remarks about the Ecological Indicator Value of Non-Pollen Palynomorphs (NPP) in the West-Frisian Cores
A.1. Fungi of Mesotrophic and Oligotrophic Conditions
A.1.1. Neurospora the Fire Indicator
A.1.2. Coprophilous Fungi
A.1.3. Zygnemataceae
References
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multi-layered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Birks, H.J.B. What is natural? The need for a long-term perspective in biodiversity conservation. Science 2006, 314, 1261–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, A.L.; Columbo, S.; Hanley, N. Improving the application of long-term ecology in conservation and land management. J. Appl. Ecol. 2013, 51, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Faber, A.H.; Kooijman, A.M.; Brinkkemper, O.; van der Plicht, J.; van Geel, B. Palaeoecological reconstructions of vegetation successions in two contrasting former turbaries in the Netherlands and implications for conservation. Rev. Palaeobot. Palynol. 2016, 233, 77–92. [Google Scholar] [CrossRef]
- Vos, P.C. Origin of the Dutch Coastal Landscape; Bakhuis Publishing: Eelde, The Netherlands, 2015. [Google Scholar]
- Van Amerongen, Y.F. Wild West Frisia: The role of Domestic and Wild Resource Exploitation in Bronze Age Subsistence. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 10 November 2016. [Google Scholar]
- Van Zijverden, W.K. After the Deluge, a Palaeogeographical Reconstruction of Bronze Age West-Frisia (2000-800 BC). Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 15 February 2017. [Google Scholar]
- Roessingh, W. Dynamiek in Beeld; Onderzoek van Westfriese Nederzettingen uit de Bronstijd. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 13 December 2018. (In Dutch). [Google Scholar]
- Bakker, J.A.; Brandt, R.W. Opgravingen te Hoogkarspel III. West-Frieslands Oud Nieuw 1966, 33, 176–224. (In Dutch) [Google Scholar]
- Buurman, J. The Eastern Part of West-Friesland in Later Prehistory. Agricultural and Environmental Aspects. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 31 October 1996. [Google Scholar]
- Pals, J.P.; van Geel, B.; Delfos, A. Paleoecological studies in the Klokkeweel bog near Hoogkarspel (prov. of Noord Holland). Rev. Palaeobot. Palynol. 1980, 30, 371–418. [Google Scholar] [CrossRef]
- Van Amerongen, Y.F. All’s well? Comparing on- and off-site pollen samples and exploring the potential of pollen from man-made contexts. Veg. Hist. Archaeobotany 2019, 29, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Buurman, J. Archaeological Investigations of a Middle and Late Bronze Age Settlement Site at Westwoud (West-Friesland). Ber. Rijksd. Oudheidkund. Bodemonderz. 1999, 43, 99–140. [Google Scholar]
- Van Geel, B.; Buurman, J.; Waterbolk, H.T. Archaeological and palaeoecological indications for an abrupt climate change in The Netherlands and evidence for climatological teleconnections around 2650 BP. J. Quat. Sci. 1996, 11, 451–460. [Google Scholar] [CrossRef]
- Van Geel, B.; van der Plicht, J.; Kilian, M.R.; Klaver, E.R.; Kouwenberg, J.H.M.; Renssen, H.; Reynaud-Farrera, I.; Waterbolk, H.T. The sharp rise of ∆14C ca. 800 cal BC: Possible causes, related climatic teleconnections and the impact on human environments. Radiocarbon 1998, 40, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Van Geel, B.; Heijnis, H.; Charman, D.J.; Thompson, G.; Engels, S. Bog burst in the eastern Netherlands triggered by the 2.8 kyr BP climate event. Holocene 2014, 24, 1465–1477. [Google Scholar] [CrossRef]
- Van Geel, B.; Hallewas, D.P.; Pals, J.P. A Late Holocene deposit under the Westfriese Zeedijk near Enkhuizen (Prov. of N-Holland, The Netherlands): Palaeoecological and archaeological aspects. Rev. Palaeobot. Palynol. 1983, 38, 269–335. [Google Scholar] [CrossRef]
- Besteman, J.C. The pre-urban development of Medemblik: From an early medieval trading centre tot a medieval town. In Medemblik and Monnickendam. Aspects of Medieval Urbanization in Northern Holland; Heidinga, H.A., van Regteren Altena, H.H., Eds.; University of Amsterdam: Amsterdam, The Netherlands, 1989. [Google Scholar]
- Borger, G.J. De Veenhoop. Een Historisch-Geografisch Onderzoek naar het Verdwijnen van het Veendek in een deel van West-Friesland; Buijten& Schipperheijn: Amsterdam, The Netherlands, 1975. (In Dutch) [Google Scholar]
- Nieuwhof, A.; Bakker, M.; Knol, E.; de Langen, G.J.; Nicolay, J.A.W.; Postma, D.; Schepers, M.; Varwijk, T.W.; Vos, P.C. Adapting to the sea: Human habitation in the coastal area of the northern Netherlands before medieval dike building. Ocean Coast. Manag. 2019, 173, 77–89. [Google Scholar] [CrossRef]
- Schrickx, C.P. Huisraad van de bewoners van Berkhout rond 1200. In Archeologisch Onderzoek op het Perceel Westeinde 322 in Berkhout, Gemeente Koggenland; West-Friese Archeologische Rapporten 105; Archeologie West-Friesland: Hoorn, The Netherlands, 2017. (In Dutch) [Google Scholar]
- ter Steege, B. Wonen in de Beetskoog. Archeologisch Onderzoek op het Perceel Beets 47 in Beets, Gemeente Zeevang; West-Friese Archeologische Rapporten 99; Archeologie West-Friesland: Hoorn, The Netherlands, 2016. (In Dutch) [Google Scholar]
- Gerritsen, S.; Schrickx, C.P. Kringgreppels rond De Leeuwenhalm. Archeologisch Onderzoek naar resten uit de Bronstijd, Middeleeuwen en Nieuwe Tijd op het Terrein van het oude Postkantoor (Hoofdstaat 17–19 en 23–29) in Bovenkarspel, gemeente Stede Broec; West-Friese Archeologische Rapporten 60; Archeologie West-Friesland: Hoorn, The Netherlands, 2014. (In Durch) [Google Scholar]
- Schrickx, C.P.; ter Steege, B.C. Smeden, gieters en tappers in een nieuwbouwwijk uit de Gouden Eeuw. In Archeologisch Onderzoek aan de Zuidzijde van de Lange Tuinstraat in Enkhuizen; West-Friese Archeologische Rapporten 123; Archeologie West-Friesland: Hoorn, The Netherlands, 2018. (In Dutch) [Google Scholar]
- Schrickx, C.P.; Duijn, D.M. Nouveau Riche aan de Nieuwe Haven. In Archeologisch Onderzoek naar woon-en Pakhuizen en het Kantoor van de WIC Tussen de Paktuinen en Nieuwe haven in Enkhuizen; West-Friese Archeologische Rapporten 83; Archeologie West-Friesland: Hoorn, The Netherlands, 2016. (In Dutch) [Google Scholar]
- Duijn, D.M. Middeleeuwse Sporen onder een 16e-eeuwse Nieuwbouwwijk: Archeologie West Friesland Rapporten 89; Archeologie West-Friesland: Hoorn, The Netherlands, 2016. (In Dutch) [Google Scholar]
- Schrickx, C.P. Enkhuizen, Vijzelstraat. In Archeologische Kroniek van Noord-Holland over 2011; Provincie Noord-Holland: Haarlem, The Netherlands, 2012; pp. 39–43. (In Dutch) [Google Scholar]
- Gerritsen, S.; Duijn, D.M. Bronstijd Greppels en een Middeleeuwse terp. Inventariserend en Aanvullend Archeologisch Onderzoek aan Westeinde 107, Gemeente Enkhuizen; West-Friese Archeologische Rapporten 66; Archeologie West-Friesland: Hoorn, The Netherlands, 2014. (In Dutch) [Google Scholar]
- Gerritsen, S.; Archeologie West-Friesland, Hoorn, The Netherlands. Personal communication, 2019.
- Gerritsen, S.; Duijn, D.M. Een middeleeuwse terp onder een 17de- eeuwse stolp. Bouwhistorische en archeologisch onderzoek aan de Oostwouder Dorpsstraat 69 in Oostwoud, gemeente Medemblik; West-Friese Archeologische Rapporten 62; Archeologie West-Friesland: Hoorn, The Netherlands, 2014. (In Dutch) [Google Scholar]
- Bartels, M.H. Dwars door de Dijk. Archeologisch en Historisch Onderzoek van de Westfriese Omringdijk Tussen Hoorn en Enkhuizen; Publicaties Stichting Archeologie West-Friesland: Hoorn, The Netherlands, 2016. (In Dutch) [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; Wiley: Chichester, UK, 1989. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis, 2nd ed.; Blackwell: Oxford, UK, 1991; pp. 1–216. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete; Verlag Dr Friedrich Pfeil: München, Germany, 2004. (In German) [Google Scholar]
- Van Geel, B. A palaeoecological study of Holocene peat bog sections in Germany and the Netherlands. Rev. Palaeobot. Palynol. 1978, 25, 1–120. [Google Scholar] [CrossRef]
- Van Geel, B.; Aptroot, A. Fossil ascomycetes in Quaternary deposits. Nova Hedwig. 2006, 82, 313–329. [Google Scholar] [CrossRef]
- Van Geel, B.; Bohncke, S.J.P.; Dee, H. A palaeoecological study of an upper late glacial and holocene sequence from “de borchert”, The Netherlands. Rev. Palaeobot. Palynol. 1981, 31, 367–448. [Google Scholar] [CrossRef]
- Van Geel, B.; Coope, G.R.; van der Hammen, T. Palaeoecology and stratigraphy of the Lateglacial type section at Usselo (The Netherlands). Rev. Palaeobot. Palynol. 1989, 60, 25–129. [Google Scholar] [CrossRef]
- Van Geel, B.; Buurman, J.; Brinkkemper, O.; Schelvis, J.; Aptroot, A.; van Reenen, G.; Hakbijl, T. Environmental reconstruction of a Roman Period settlement site in Uitgeest (The Netherlands), with special reference to coprophilous fungi. J. Archaeol. Sci. 2003, 30, 873–883. [Google Scholar] [CrossRef]
- Grimm, E.C. Tilia and Tiliagraph Version 2.0.41; Illinois State Museum: Springfield, IL, USA, 1991. [Google Scholar]
- Mauquoy, D.; van Geel, B. Mire and peat macros. In Encyclopedia of Quaternary Science; Elias, S.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 2315–2336. [Google Scholar]
- Cappers, R.T.J.; Bekker, R.M.; Jans, J.E.A. Digital seed atlas of the Netherlands. Groningen Archaeological Studies 4; Barkhuis Publishing: Groningen, The Netherlands, 2006. [Google Scholar]
- Körber-Grohne, U. Bestimmungsschlüssel für subfossile Juncus-Samen and Gramineen-Früchte. Probl. Küstenforschung Südl. Nordseegebiet 1964, 7, 1–47. (In German) [Google Scholar]
- Körber-Grohne, U. Bestimmungsschlüssel für subfossile Gramineen-Früchte. Probl. Küstenforschung Südl. Nordseegebiet 1991, 18, 169–234. (In German) [Google Scholar]
- BLWG. Available online: http://www.verspreidingsatlas.nl/mossen (accessed on 12 December 2018).
- Landwehr, J. Nieuwe Atlas Nederlandse Bladmossen; Thieme: Zutphen, The Netherlands, 1984; pp. 1–568. (In Dutch) [Google Scholar]
- Nyholm, E. Illustrated Moss Flora of Fennoscandia II. Musci Fasc. 1–6; Swedish Natural Science Research Council: Stockholm, Sweden, 1954. [Google Scholar]
- Siebel, H.; During, H. Beknopte Mosflora van Nederland en België; KNNV Uitgeverij: Utrecht, The Netherlands, 2006. (In Dutch) [Google Scholar]
- Smith, A.J.E. The Moss Flora of Britain and Ireland, 2nd ed.; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Touw, A.; Rubers, W.V. De Nederlandse Bladmossen; KNNV Uitgeverij: Utrecht, The Netherlands, 1989. (In Dutch) [Google Scholar]
- Hedenäs, L.; Department of Botany, Swedish Museum of Natural History, Stockholm, Sweden. Personal communication, 2019.
- Vorst, O. Catalogus van de Nederlandse kevers (Coleoptera); Monografieën van de Nederlandse Entomologische Vereniging: Amsterdam, The Netherlands, 2010. (In Dutch) [Google Scholar]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Bronk Ramsey, C.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef] [Green Version]
- Bronk Ramsey, C. OxCal Version 4.3 (Online Version); Oxford University: Oxford, UK, 2019. [Google Scholar]
- Hedenäs, L.; Kooijman, A.M. Phylogeny and habitat adaptations within a monophyletic group of wetland moss genera (Amblystegiaceae). Plant Syst. Evol. 1996, 199, 33–52. [Google Scholar] [CrossRef]
- Bohncke, S.J.P.; van Haaster, H.; Wiegers, J. Paludella squarrosa (Hedw.) Brid. in a Late Subboreal Holland Peat sequence. J. Bryol. 1984, 13, 219–226. [Google Scholar] [CrossRef]
- Van Geel, B.; Bos, J.M.; Pals, J.P. Archaeological and palaeoecological aspects of a medieval house terp in a reclaimed raised bog area in North Holland. Ber. Rijksd. Oudheidk. Bodemonderz. 1983, 33, 419–444. [Google Scholar]
- Van der Plicht, J. Radiocarbon, the calibration curve and Scythian chronology. In Impact of the Environment on Human Migration in Eurasia; Scott, E.M., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 45–61. [Google Scholar]
- Shumilovskikh, L.S.; Schlütz, F.; Achterberg, I.; Bauerochse, A.; Leuschner, H.H. Non-Pollen Palynomorphs from mid-Holocene peat of the raised bog Borsteler Moor (Lower Saxony, Germany). Stud. Quat. 2015, 32, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Martin-Puertas, C.; Matthes, K.; Brauer, A.; Muscheler, R.; Hansen, F.; Petrick, C.; Aldahan, A.; Possnert, G.; van Geel, B. Regional atmospheric circulation shifts induced by a grand solar minimum. Nat. Geosci. 2012, 5, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Roep, T.B.; van Regteren Altena, J.F. Palaeotidal levels in tidal sediments (3800-3635 BP); compaction, sea level rise and human occupation (3275-2620 BP) at Bovenkarspel, NW Netherlands. In Tide-Influenced Sedimentary Environments and Facies; de Boer, P.L., van Gelder, A., Nio, J.D., Eds.; D. Reidel: Dordrecht, The Netherlands, 1988; pp. 215–231. [Google Scholar]
- Heidinga, H.A. Indications of severe drought during the 10th century AD from an inland dune area in the central Netherlands. Geol. Mijnbo. Delft 1984, 63, 241–248. [Google Scholar]
- Overbeck, F. Botanisch-Geologische Moorkunde; Karl Wachholtz Verlag: Neumünster, Germany, 1975. (In German) [Google Scholar]
- Gottschalk, M.K.E. Stormvloeden en Rivieroverstromingen in Nederland I, de Periode voor 1400; Van Gorkum: Assen, The Netherlands, 1971. (In Dutch) [Google Scholar]
- Gottschalk, M.K.E. Stormvloeden en Rivieroverstromingen in Nederland II, de periode 1400-1600; Van Gorkum: Assen, The Netherlands, 1975. (In Dutch) [Google Scholar]
- Van Popta, Y.T. No country for Men: Searching for late medieval submerged settlements in the north-eastern Zuiderzee area in the Netherlands. Eur. J. Archaeol. 2019, 22, 567–587. [Google Scholar] [CrossRef] [Green Version]
- Van Popta, Y.T.; Westerdahl, C.L.; Duncan, B.G. Maritime culture in the Netherlands: Accessing the late medieval maritime cultural landscapes of the north-eastern Zuiderzee. Int. J. Naut. Archaeol. 2019, 48, 172–188. [Google Scholar]
- Van Smeerdijk, D.; Pals, J.P.; van Geel, B.; van Bennekom, J. Mens en Landchap in middeleeuws Diemen. In Diemen in Het Land van Amstel; Blok, H., Krook, W., van Reenen, P., Wiggers, R., Eds.; De Bataafse Leeuw: Amsterdam, The Netherlands, 2009; pp. 351–366. (In Dutch) [Google Scholar]
- McCarroll, J.; Chambers, F.M.; Webb, J.C.; Thom, T. Using palaeoecology to advise peatland conservation: An example from West Arkengarthdale, Yorkshire, UK. J. Nat. Conserv. 2016, 30, 90–102. [Google Scholar] [CrossRef]
- Roche, J.R.; Mitchell, F.J.G.; Waldren, S.; Stefanini, B.S. Palaeoecological evidence for survival of Scots pine through the Late Holocene in western Ireland: Implications for ecological management. Forests 2018, 9, 350. [Google Scholar] [CrossRef] [Green Version]
- Kuhry, P.; Nicholson, B.J.; Gignac, L.D.; Vitt, D.H.; Bayley, S.E. Development of Sphagnum-dominated peatlands in boreal continental Canada. Can. J. Bot. 1993, 71, 10–22. [Google Scholar] [CrossRef]
- Lust, N.; Geudens, G.; Oltshoorn, A.F.M. Scots pine in Belgium and The Netherlands. Investig. Agrar. Sist. Recur. For. 2000, 9, 213–231. [Google Scholar]
- Jamrichová, E.; Bobek, P.; Šolcová, A.; Tkáč, P.; Hédl, R.; Valachovič, M. Lowland pine forest in the northwestern Pannonian Basin: Between natural vegetation and modern plantations. Reg. Environ. Chang. 2019, 19, 2395–2409. [Google Scholar] [CrossRef]
- Brinkkemper, O. Rotterdam en omgeving van de IJzertijd tot en met de Late Middeleeuwen; een milieu-reconstructie op basis van onderzoek van plantenresten. In Archeologisch Onderzoek in het Tracé van de Willemsspoortunnel te Rotterdam; Carmiggelt, A., Guiran, A.J., van Trierum, M.C., Eds.; BOOR: Rotterdam, The Netherlands, 1997; pp. 45–71. (In Dutch) [Google Scholar]
- Dennis, R.W.G. British Ascomycetes; J. Cramer.: Vaduz, Liechenstein, 1978; p. 585. [Google Scholar]
- Van Geel, B.; Grenfell, H.R. Spores of Zygnemataceae. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: New York, NY, USA, 1996; Chapter 7A; pp. 173–179. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
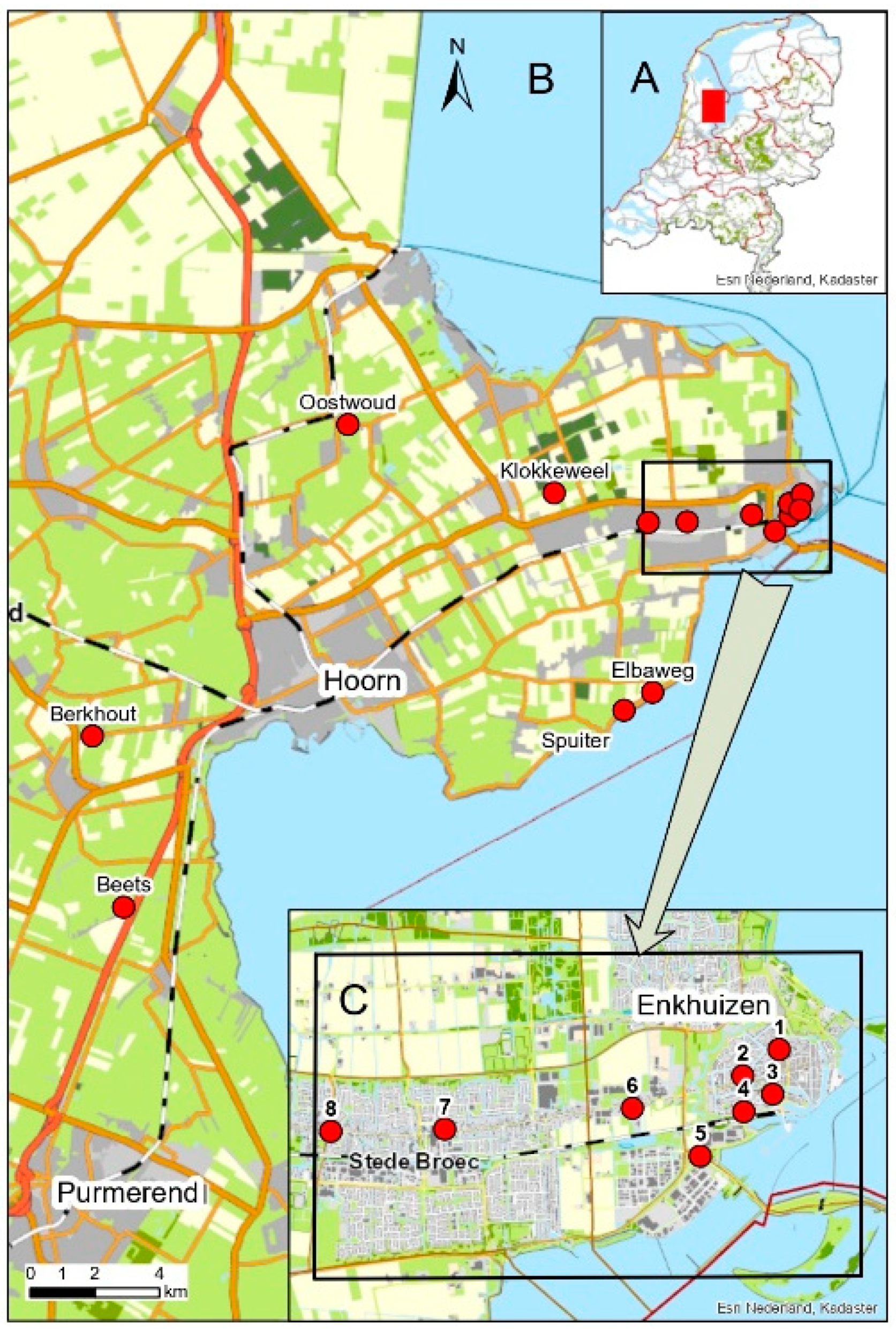
| Project | Archaeological Site Names | Coordinates | Start of Organic Deposit (m-NAP) | Bronze Age Archaeology | Overlying Deposits | Reference | ||
|---|---|---|---|---|---|---|---|---|
| No. | Codes and Short Site Names | X | Y | |||||
| 410 | BKH: Berkhout | Berkhout, Westeinde 322 | 126,411 | 516,703 | 2.88 | Absent | Anthropogenic. 12th or 13th C AD | [21] |
| 414 | BTS: Beets | Beets 47 | 127,375 | 511,361 | No data | Absent | Anthropogenic. Mid 14th C AD | [22] |
| 328 | BVK: Bovenkarspel | Bovenkarspel, Hoofdstraat 17–19 | 144,798 | 523,401 | 1.98 | Middle and Late Bronze Age occupation | Anthropogenic. 12th C AD | [23] |
| 433 | ELT: Lange Tuinstraat | Enkhuizen, Lange Tuinstraat 1 tm 35 | 147,971 | 523,588 | 2.39 | Occupation 300 m from sampling site | Reclamation 12th C AD | [24] |
| 372 | EPK: Paktuinen | Enkhuizen, Paktuinen-Ridderstraat | 148,277 | 523,777 | 3.74 | Occupation 600 m from sampling site | Reclamation 12th C AD. Anthrop. from AD 1590 | [25] |
| 381 | ERS: Raamstraat | Enkhuizen, Raamstraat | 147,960 | 523,973 | 2.72 | Absent | Anthropogenic.12th-13th C AD | [26] |
| 341 | EVT: Vijzeltuin | Enkhuizen, Vijzeltuin-West | 148,345 | 524,256 | 2.59 | Absent | Anthropogenic. 15th C AD | [27] |
| 369 | EWE: Westeinde | Enkhuizen, Westeinde 107 | 146,789 | 523,626 | 2.39 | Middle Bronze Age ditches | Anthropogenic. 12th C AD | [28] |
| 431 | GBZ: Grootebroek | Grotebroek, Zesstedenweg 203 | 143,591 | 523,378 | 2.98 | Absent | Anthropogenic. 12th-13th C AD | [29] |
| 383 | OWD: Oostwoud | Oostwoud, Oostwouderdorpsstraat 69 | 134,302 | 527,441 | 1.66 | Bronze Age ard marks | Anthropogenic. 13th C AD | [30] |
| 325 | VEL: Elbaweg | Zuiderdijk, Venhuizen, Elbaweg | 143,719 | 518,069 | 2.64 | Middle Bronze Age ditches, pits | Anthropogenic. 14th C AD | [31] |
| 325 | VSP: Spuiter | Zuiderdijk, Venhuizen, Spuiter | 142,838 | 517,515 | 2.41 | Absent | Anthropogenic. 14th C AD | [31] |
| Type | Species |
|---|---|
| Atriplex littoralis-type | Atriplex littoralis/prostrata var. salina |
| Carex acuta-type | Carex acuta/elata/nigra/trinervis |
| Carex paniculata-type | Carex appropinquata/diandra/paniculata |
| Juncus articulatus-type | Juncus acutiflorus/articulatus/bulbosus |
| Juncus effusus-type | Juncus conglomeratus/effusus/inflexus |
| Ranunculus repens-type | Ranunculus acris/bulbosus/repens |
| Sagina apetala-type | Sagina apetala/micropetala/procumbens |
| Veronica beccabunga-type | Veronica anagallis-aquatica/beccabunga/catenata |
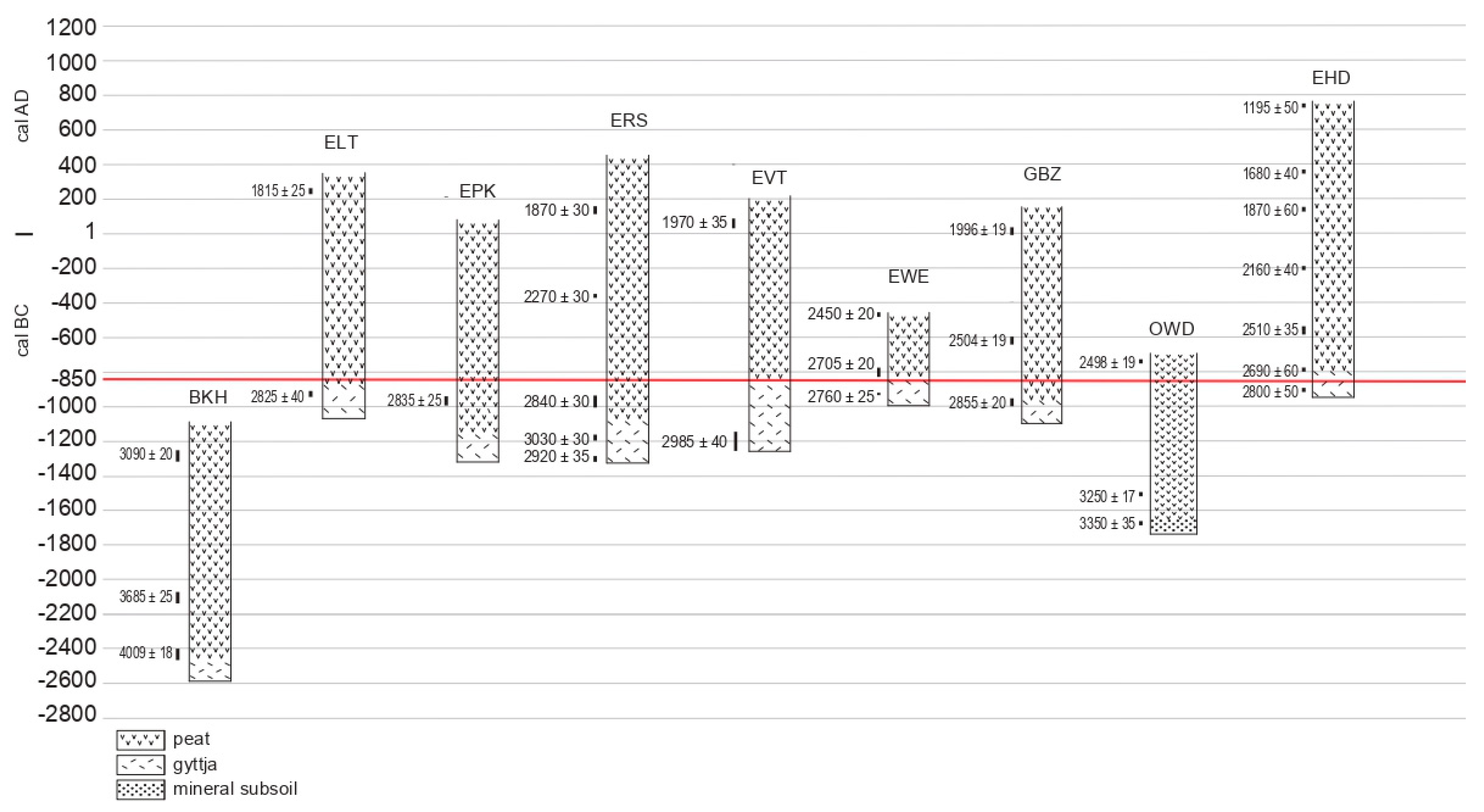
| Sites | Depth (cm) | Lab. Code | 14C BP | Calibrations OxCAL 2019–06-09 (IntCal 13): 68.2% Probability | Material Used for Dating |
|---|---|---|---|---|---|
| BKH: Berkhout | 7 + 8 | GrM-17116 | 3090 ± 20 | cal BC 1406–1381; 1343–1306 | Betula leaf, seed, catkin scale, bud scale. |
| 32 + 33 | GrM-17117 | 3685 ± 25 | cal BC 2133–2082; 2060–2031 | Betula leaf, bud scale; seeds S. flos-cuculi. | |
| 42 + 43 | GrM-17347 | 4009 ± 18 | cal BC 2568–2519; 2498–2489 | Carex seeds. | |
| BTS: Beets | 7 + 8 | GrM-17115 | 1765 ± 20 | cal AD 240–259; 281–324 | R. alba seeds, stems; Sphagnum opercula. |
| ELT: Enkhuizen-Langetuinstr | 29 | GrM-17125 | 1815 ± 25 | cal AD 140–160; 166–196; 208–238 | Betula bud scales, leaves; Musci. |
| 65 | GrM-17839 | 2825 ± 40 | cal BC 1022–919 | Seeds: Alisma, Carex, Mentha, Lythrum. | |
| EPK: Enkhuizen-Paktuinen | 18 + 19 | GrM-17284 | 1950 ± 20 | cal AD 25–75 | Menyanthes seed, Betula leaf. |
| 62 + 63 | GrM-17348 | 2835 ± 25 | cal BC 1018–970; 961–935 | Carex seeds, Iris pseudacorus seed. | |
| ERS: Enkhuizen-Raamstraat | 16 + 17 | Beta-431213 | 1870 ± 30 | cal AD 82–170; 194–210 | leaves: Betula, Oxycoccus palustris, musci. |
| 37 | Beta-431214 | 2270 ± 30 | cal BC 395–357; 282–257; 244–236 | Betula leaf, female catkin scale; fern leaf. | |
| 68 + 69 + 70 | Beta-431215 | 2840 ± 30 | cal BC 1043–971; 960–936 | Seeds: Cyperaceae, Apiaceae, Iris, Lycopus. | |
| 84 + 85 + 86 | Beta-431216 | 3030 ± 30 | cal BC 1374–1354; 1302–1226 | Seeds: Cyperaceae, Apiaceae, Iris, Mentha. | |
| 90 + 91 + 92 | GrA-64000 | 2920 ± 35 | cal BC 1192–1172; 1166–1143; 1132–1051 | Seeds: Mentha, Glyceria, Apiaceae, Alisma, Solanum, Eleocharis. | |
| EVT: Enkhuizen-Vijzeltuin | 18 | GrA-56077 | 985 ± 35 | cal AD 1014–1048; 1087–1123; 1138–1149 | Seeds: Chenopodiaceae. |
| 21 | GrA-56075 | 1970 ± 35 | cal BC 19–13; cal AD 1–72 | Pinus, female cone fragment. | |
| 69 + 70 | GrA-56073 | 2985 ± 40 | cal BC 1267–1155; 1148–1128 | Seeds: Cyperaceae, Mentha. | |
| EWE: Enkhuizen-Westeinde | 13 | GrM-17285 | 2450 ± 20 | cal BC 742–687; 665–645; 550–485 | Seeds: Carex, Menyanthes, Apiaceae. |
| 25 + 26 | GrM-17287 | 2705 ± 20 | cal BC 894–871; 851–818 | Seeds: Menyanthes, Iris, Apiaceae, Solanum, Ranunculaceae, Caryophyllaceae. | |
| 37 | GrM-17124 | 2760 ± 25 | cal BC 927–891; 879–846 | Iris pseudacorus seed. | |
| GBZ: Grootebroek-Zesstedenweg | 48 + 49 | GrM-17206 | 1996 ± 19 | cal BC 37–29; 22–10; cal BC 2–cal AD 25 | Pinus needles, Betula leaf, seed Menyanthes. |
| 69 + 70 | GrM-17207 | 2504 ± 19 | cal BC 767–748; 684–667 | Seeds: Carex, Menyanthes; musci, fern leaf. | |
| cal BC 641–588; 580–559 | |||||
| 96 + 97 | GrM-17209 | 2855 ± 20 | cal BC 1051–979 | Carex seeds. | |
| OWD: Oostwoud | 29 | GrM-17210 | 2498 ± 19 | cal BC 762–748; 685–666 | Seeds: Menyanthes, Carex, S. flos-cuculi. |
| cal BC 642–587; 581–556 | Betula, Caryophyll., Sphagnum, Polytrichum. | ||||
| 61 | GrM-17211 | 3250 ± 17 | cal BC 1595–1589; 1532–1499 | Seeds: Carex, Ranunc., Menyanthes, Lycopus, Apiaceae, Caryophyllaceae. | |
| 68 | GrM-17879 | 3350 ± 35 | cal BC 1690–1611; 1572–1566 | Seeds: Urtica, Hypericum, Mentha, Solanum, Carex, Juncus. | |
| VEL: Venhuizen-Elbaweg | 53–54 | GrA-53539 | 1645 ± 30 | cal AD 353–367; 380–426 | Alkaline fraction of bulk peat. |
| 68–70 | GrA-53538 | 2465 ± 30 | cal BC 751–683; 669–636 | ||
| cal BC 626–614; 592–516 | |||||
| VSP: Venhuizen-Spuiter | 30 | GrA-53540 | 2450 ± 35 | cal BC 747–686; 666–643; | Alkaline fraction of bulk peat. |
| cal BC 554–452; 446–430 | |||||
| Ditch | GrA-53541 | 1555 ± 30 | cal AD 430–494; 510–518; | Leaf fragments, bud scales, seeds. | |
| cal AD 528–545 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Geel, B.; Brinkkemper, O.; van Reenen, G.B.A.; Van der Putten, N.N.L.; Sybenga, J.E.; Soonius, C.; Kooijman, A.M.; Hakbijl, T.; Gosling, W.D. Multicore Study of Upper Holocene Mire Development in West-Frisia, Northern Netherlands: Ecological and Archaeological Aspects. Quaternary 2020, 3, 12. https://doi.org/10.3390/quat3020012
van Geel B, Brinkkemper O, van Reenen GBA, Van der Putten NNL, Sybenga JE, Soonius C, Kooijman AM, Hakbijl T, Gosling WD. Multicore Study of Upper Holocene Mire Development in West-Frisia, Northern Netherlands: Ecological and Archaeological Aspects. Quaternary. 2020; 3(2):12. https://doi.org/10.3390/quat3020012
Chicago/Turabian Stylevan Geel, Bas, Otto Brinkkemper, Guido B.A. van Reenen, Nathalie N.L. Van der Putten, Jasmijn E. Sybenga, Carla Soonius, Annemieke M. Kooijman, Tom Hakbijl, and William D. Gosling. 2020. "Multicore Study of Upper Holocene Mire Development in West-Frisia, Northern Netherlands: Ecological and Archaeological Aspects" Quaternary 3, no. 2: 12. https://doi.org/10.3390/quat3020012
APA Stylevan Geel, B., Brinkkemper, O., van Reenen, G. B. A., Van der Putten, N. N. L., Sybenga, J. E., Soonius, C., Kooijman, A. M., Hakbijl, T., & Gosling, W. D. (2020). Multicore Study of Upper Holocene Mire Development in West-Frisia, Northern Netherlands: Ecological and Archaeological Aspects. Quaternary, 3(2), 12. https://doi.org/10.3390/quat3020012