Slash-and-Burn Practices Decrease Arbuscular Mycorrhizal Fungi Abundance in Soil and the Roots of Didierea madagascariensis in the Dry Tropical Forest of Madagascar


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Soil Chemistry
2.4. Lipid Extraction and Analysis
2.5. Microscopic Analysis of Root Colonization
2.6. Spore Extraction and Analysis
2.7. Statistical Analysis
3. Results
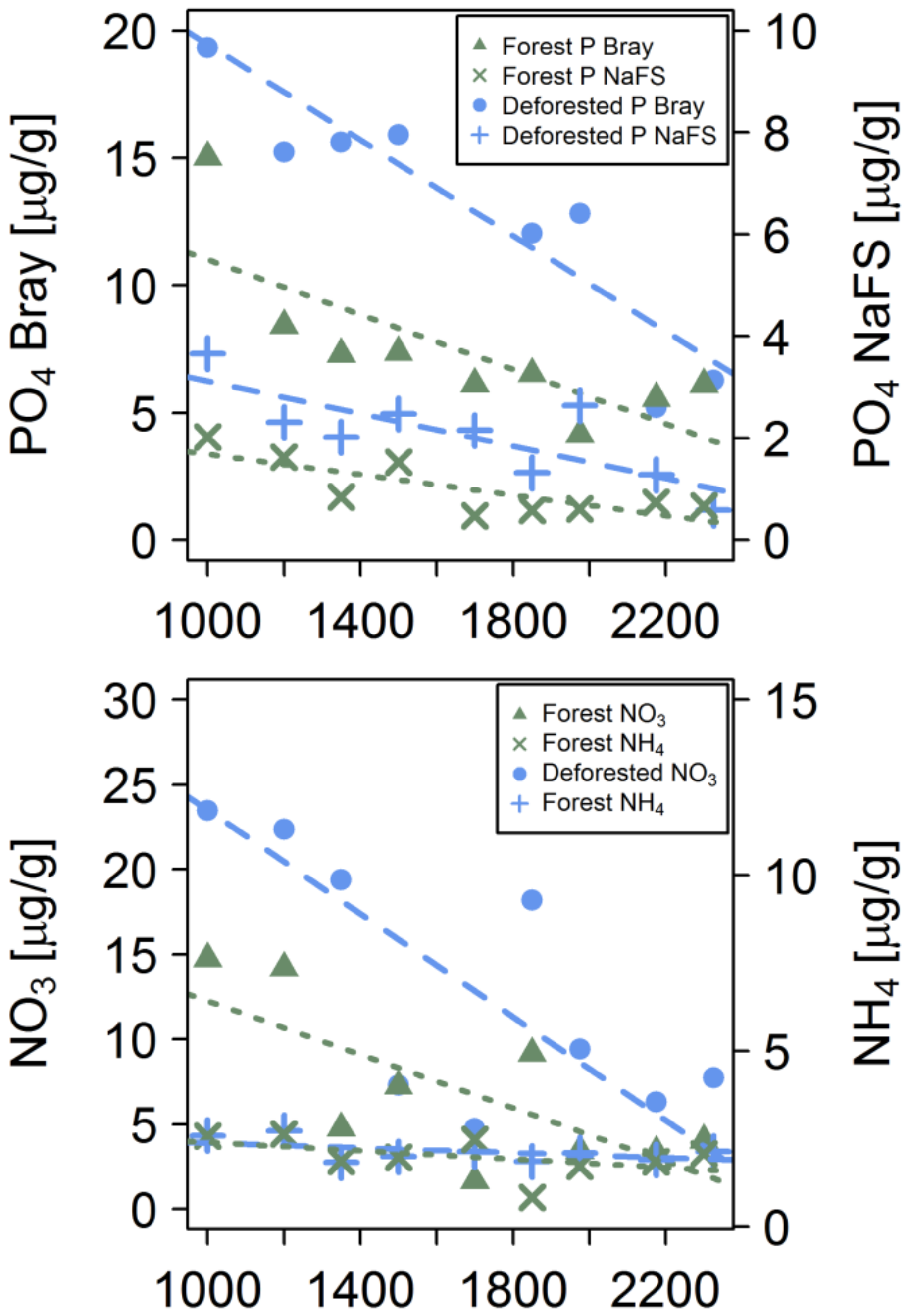
3.1. Soil Chemistry
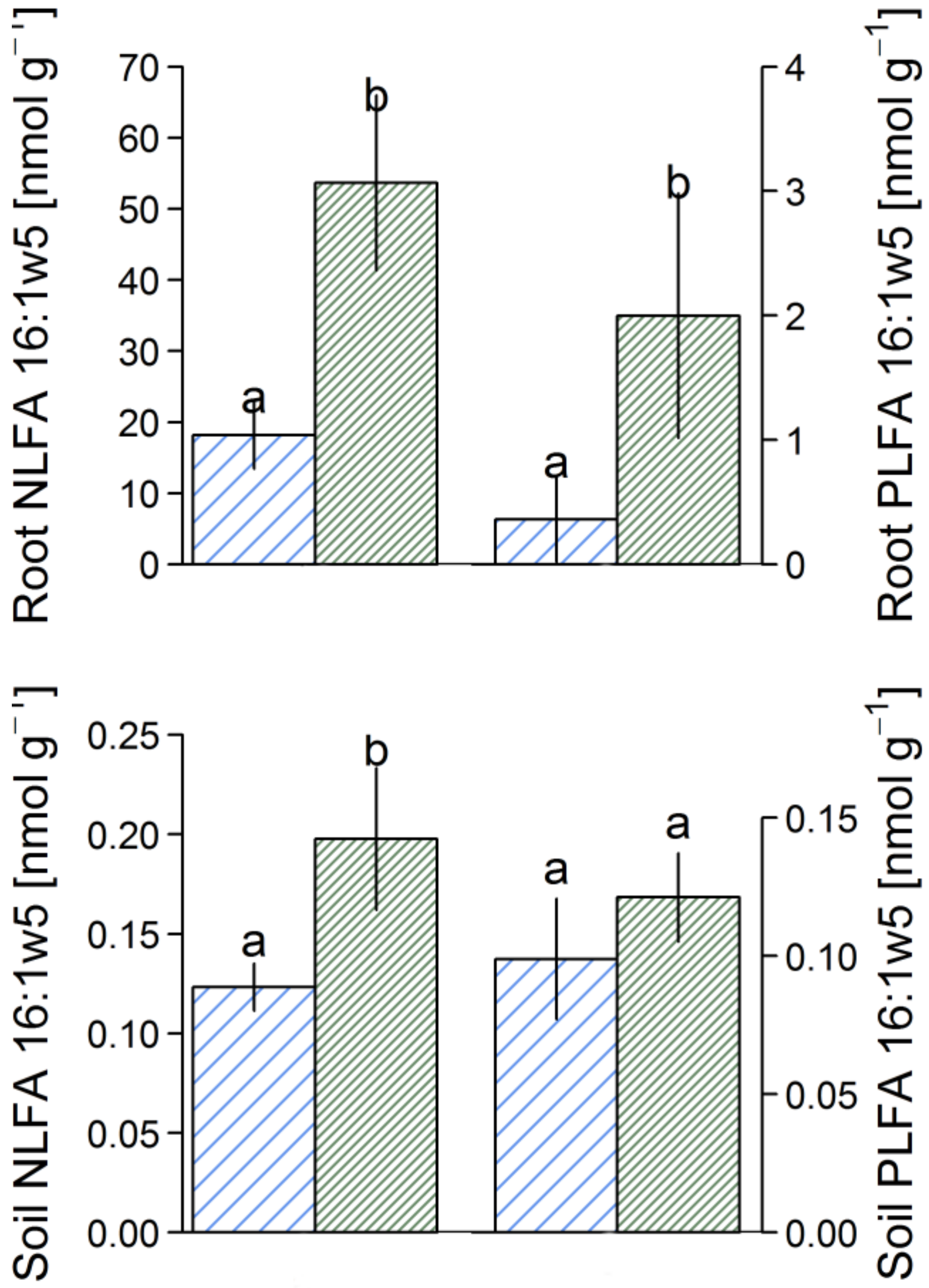
3.2. Fatty Acid Analysis
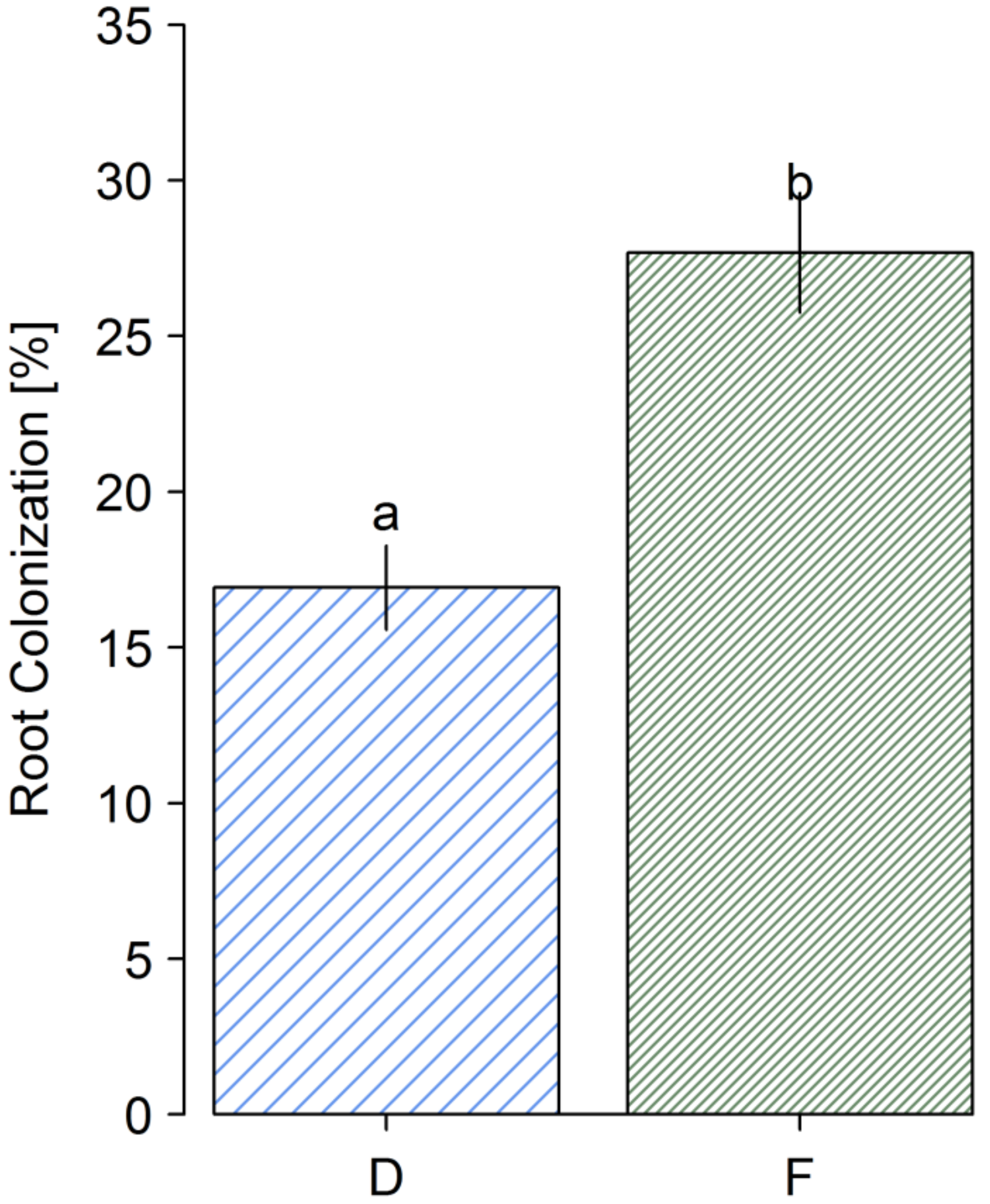
3.3. Microscopic Analysis of Root Colonization
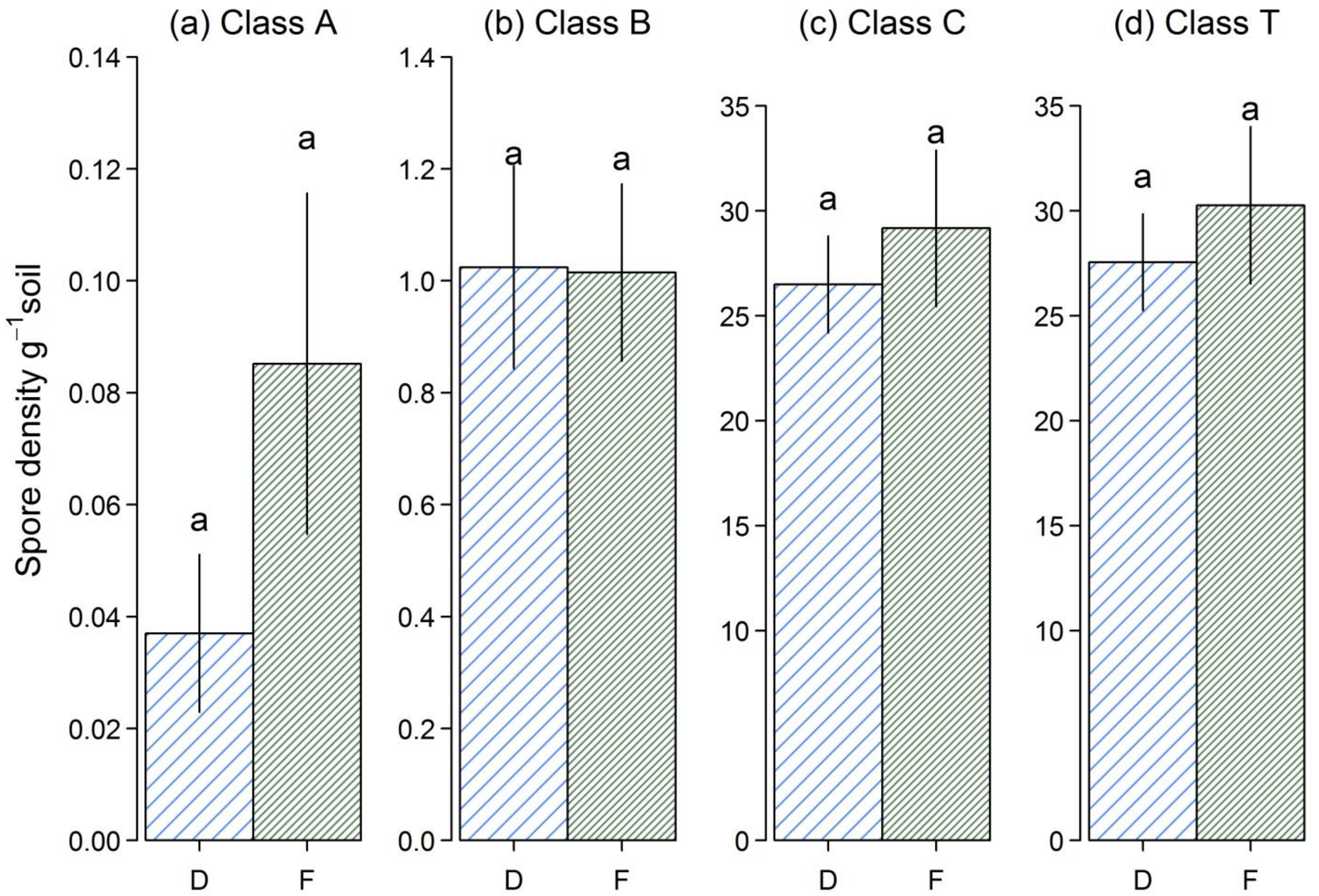
3.4. Soil Spore Density
3.5. Relationships between AM Fungi Abundance and Soil Parameters
4. Discussion
4.1. Soil Chemistry
4.2. AMF Abundance in Soil and Roots of D. Madagascariensis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance (m) | NH4 (µg/g) | NO3 (µg/g) | N (µg/g) | Bray (µg/g) | NaFS (µg/g) | C (mg/g) | C/N Ratio | pH | |
|---|---|---|---|---|---|---|---|---|---|
| Distance | 1 | −0.45 | −0.57 ** | −0.12 | −0.68 ** | −0.58 ** | −0.22 | −0.28 | −0.49 * |
| NH4 | 1 | 0.31 | 0.09 | 0.4 | 0.46 | 0.2 | 0.24 | 0.13 | |
| NO3 | 1 | 0.64 ** | 0.75 ** | 0.62 ** | 0.74 ** | 0.55 * | 0.79 ** | ||
| N | 1 | 0.41 | 0.29 | 0.96 ** | 0.43 | 0.74 ** | |||
| Bray | 1 | 0.86 ** | 0.47 * | 0.37 | 0.81 ** | ||||
| NaFS | 1 | 0.35 | 0.36 | 0.60 ** | |||||
| C | 1 | 0.61 ** | 0.73 ** | ||||||
| Ratio | 1 | 0.31 | |||||||
| pH | 1 |
| Distance (m) | NH4 (µg/g) | NO3 (µg/g) | N (µg/g) | Bray (µg/g) | NaFS (µg/g) | C (mg/g) | Ratio | pH | |
|---|---|---|---|---|---|---|---|---|---|
| Colonization | 0.3 | 0.14 | −0.34 | −0.5 * | −0.37 | −0.34 | −0.45 * | −0.22 | −0.5 * |
| Root PLFA 16:1ω5 | 0.46 * | −0.28 | −0.46 * | −0.15 | −0.4 | −0.17 | −0.18 | −0.09 | −0.46 * |
| Root NLFA 16:1ω5 | 0.03 | −0.15 | −0.31 | −0.23 | −0.26 | −0.23 | −0.21 | −0.02 | −0.33 |
| Soil NLFA 16:1ω5 | −0.16 | 0.08 | 0.11 | 0.33 | −0.06 | −0.11 | 0.42 | 0.24 | 0.13 |
| Soil PLFA 16:1ω5 | −0.21 | 0.36 | 0.19 | 0.36 | 0.14 | 0.13 | 0.47 * | 0.34 | 0.26 |
| Spores | −0.1 | 0.16 | 0.15 | −0.08 | 0.06 | 0.13 | 0.08 | 0.28 | −0.04 |
| Colonization (%) | RootPLFA (nmol g soil−1) | RootNLFA (nmol g soil−1) | SoilNLFA (nmol g soil−1) | SoilPLFA (nmol g soil−1) | Spores (100 g−1) | |
|---|---|---|---|---|---|---|
| Colonization | 1 | 0.18 | 0.56 * | 0.19 | −0.01 | 0.12 |
| Root PLFA 16:1w5 | 1 | 0.54 * | 0.15 | 0.08 | 0.11 | |
| Root NLFA 16:1ω5 | 1 | 0.46 | 0.22 | 0.2 | ||
| Soil NLFA 16:1ω5 | 1 | 0.82 ** | 0.42 | |||
| Soil PLFA 16:1ω5 | 1 | 0.22 | ||||
| Spores | 1 |
References
- Harper, G.J.; Steininger, M.K.; Tucker, C.J.; Juhn, D.; Hawkins, F. Fifty years of deforestation and forest fragmentation in Madagascar. Environ. Conserv. 2007, 34, 325–333. [Google Scholar] [CrossRef]
- Goodman, S.M.; Benstead, J.P. The Natural History of Madagascar; Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Seddon, N.; Tobias, J.; Yount, J.W.; Ramanampamonjy, J.R.; Butchart, S.; Randrianizahana, H. Conservation issues and priorities in the Mikea Forest of south-western Madagascar. Oryx 2000, 34, 287–304. [Google Scholar] [CrossRef]
- Callmander, M.W.; Phillipson, P.B.; Schatz, G.E.; Andriambololonera, S.; Rabarimanarivo, M.; Rakotonirina, N.; Raharimampionona, J.; Chatelain, C.; Gautier, L.; Lowry, P. The endemic and non-endemic vascular flora of Madagascar updated. Plant Ecol. Evol. 2011, 144, 121–125. [Google Scholar] [CrossRef]
- Wilmé, L.; Goodman, S.M.; Ganzhorn, J.U. Biogeographic Evolution of Madagascar’s Microendemic Biota. Science 2006, 312, 1063–1065. [Google Scholar] [CrossRef] [PubMed]
- Raherison, S.M.; Grouzis, M. Plant biomass, nutrient concentration and nutrient storage in a tropical dry forest in south-west Madagascar. Plant Ecol. 2005, 180, 33–45. [Google Scholar] [CrossRef]
- Casse, T.; Milhoj, A.; Ranaivoson, S.; Randriamanarivo, J.R. Causes of deforestation in southwestern Madagascar: What do we know? Forest Policy Econ. 2004, 6, 33–48. [Google Scholar] [CrossRef]
- Randriambanona, H.; Razanaka, S.; Randriamalala, J.; Grouzis, M. Secondary succession and root biomass changes in Madagascar dry deciduous forest (Mikea Forest). Rev. Ecol.-Terre Vie 2018, 73, 283–292. [Google Scholar]
- Giardina, C.P.; Sanford, R.L.; Dockersmith, I.C.; Jaramillo, V.J. The effects of slash burning on ecosystem nutrients during the land preparation phase of shifting cultivation. Plant Soil 2000, 220, 247–260. [Google Scholar] [CrossRef]
- Beliveau, A.; Davidson, R.; Lucotte, M.; Lopes, L.O.D.; Paquet, S.; Vasseur, C. Early effects of slash-and-burn cultivation on soil physicochemical properties of small-scale farms in the Tapajos region, Brazilian Amazon. J. Agric. Sci. 2015, 153, 205–221. [Google Scholar] [CrossRef]
- Ribeiro Filho, A.; Adams, C.; Manfredini, S.; Aguilar, R.; Neves, W.A. Dynamics of soil chemical properties in shifting cultivation systems in the tropics: A meta-analysis. Soil Use Manag. 2015, 31, 474–482. [Google Scholar] [CrossRef]
- Don, A.; Schumacher, J.; Freibaurer, A. Impact of tropical land-use change on soil organic carbon stocks—A meta-analysis. Glob. Change Biol. 2011, 17, 1658–1670. [Google Scholar] [CrossRef]
- Mukul, S.A.; Herbohn, J. The impacts of shifting cultivation on secondary forests dynamics in tropics: A synthesis of the key findings and spatio temporal distribution of research. Environ. Sci. Policy 2016, 55, 167–177. [Google Scholar] [CrossRef]
- Bellgard, S.E.; Whelan, R.J.; Muston, R.M. The impact of wildfire on vesicular arbuscular mycorrhizal fungi and their potential to influence the re-establishment of post-fire plant communities. Mycorrhiza 1994, 4, 139–146. [Google Scholar] [CrossRef]
- Rodrigues, J.L.M.; Pellizari, V.; Mueller, R.; Baek, K.; Jesuse, E.C.; Paula, F.; Mirzaa, B.; Hamaoui, G.; Mui Tsai, S.; Feiglf, B.; et al. Conversion of the Amazon rainforest to agriculture results in biotic homogenization of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef] [PubMed]
- García de León, D.; Neuenkamp, L.; Moora, M.; Öpik, M.; Davison, J.; Peña-Venegas, C.P.; Zobel, M. Arbuscular mycorrhizal fungal communities in tropical rain forest are resilient to slash-and-burn agriculture. J. Trop. Ecol. 2018, 34, 186–199. [Google Scholar] [CrossRef]
- Styger, E. Degrading uplands in the rainforest region of Madagascar: Fallow biomass, nutrient stocks, and soil nutrient availability. Agroforest. Syst. 2009, 77, 107–122. [Google Scholar] [CrossRef]
- Leprun, J.C.; Grouzis, M.; Randriambanona, H. Post-cropping change and dynamics in soil and vegetation properties after forest clearing: Example of the semi-arid Mikea Region (south-western Madagascar). Geoscience 2009, 341, 526–537. [Google Scholar] [CrossRef]
- Gay-des-Combes, J.M.; Robroek, B.J.M.; Herve, D.; Guillaume, T.; Pistocchi, C.; Mills, R.T.E.; Buttler, A. Slash-and-burn agriculture and tropical cyclone activity in Madagascar: Implication for soil fertility dynamics and corn performance. Agric. Ecosyst. Environ. 2017, 239, 207–218. [Google Scholar] [CrossRef]
- Runyan, C.W.; D’Odorico, P.; Lawrence, D. Physical and biological feedbacks of deforestation. Rev. Geophys. 2012, 50, RG4006. [Google Scholar] [CrossRef]
- Styger, E.; Rakotondramasy, H.; Pfeffer, M.J.; Fernandes, E.C.M.; Bates, D.M. Influence of slash-and-burn farming practices on fallow succession and land degradation in the rainforest region of Madagascar. Agr. Ecosyst. Environ. 2007, 119, 257–269. [Google Scholar] [CrossRef]
- Raharimalala, O.; Buttler, A.; Schlaepfer, R.; Gobat, J.M. Quantifying biomass of secondary forest after slash and burn cultivation in central Menabe, Madagascar. J. Trop. For. Sci. 2012, 24, 474–489. [Google Scholar]
- Newman, E.I.; Reddell, P. The distribution of mycorrhizas among families of vascular plants. New Phytol. 1987, 16, 744–751. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Johnson, N.C. Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytol. 2010, 185, 631–647. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, M.G.A.; Boller, T.; Wiemken, A.; Sanders, I.R. Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure. Ecology 1998, 79, 2081–2091. [Google Scholar] [CrossRef]
- Rillig, M.C. Large contribution of arbuscular mycorrhizal fungi to soil carbon pools in tropical forest soils. Plant Soil 2001, 223, 167–177. [Google Scholar] [CrossRef]
- Johnson, N.C.; Wedin, D.A. Soil, carbon, nutrients and mycorrhizae during conversion of dry tropical forest to grassland. Ecol. Appl. 1997, 7, 171–182. [Google Scholar] [CrossRef]
- Muchane, M.N.; Muchane, M.; Mugoya, C.; Masiga, C.W. Effect of land use system on Arbuscular mycorrhiza fungi in Maasai Mara ecosystem, Kenya. Afr. J. Microbiol. Res. 2012, 6, 3904–3916. [Google Scholar]
- Wubet, T.; Kottke, I.; Teketay, D.; Oberwinkler, F. Mycorrhizal status of indigenous trees in dry Afromontane forests of Ethiopia. For. Ecol. Manag. 2003, 179, 387–399. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Jairus, T.; Bechem, E.; Chinoya, S.; Mpumba, R.; Leal, M.; Randrianjohany, E.; Razafimandimbison, S.; Sadam, A.; et al. Spatial structure and the effects of host and soil environments on communities of ectomycorrhizal fungi in wooded savannas and rain forests of Continental Africa and Madagascar. Mol. Ecol. 2011, 20, 3071–3080. [Google Scholar] [CrossRef] [PubMed]
- Ducousso, M.; Ramanankierana, H.; Duponnois, R.; Rabévohitra, R.; Randrihasipara, L.; Vincelette, M.; Dreyfus, B.; Prin, Y. Mycorrhizal status of native trees and shrubs from eastern Madagascar litoral Forests with special emphasis on one new ectomycorrhizal endemic family, the Asteropeiaceae. New Phytol. 2008, 178, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Yokoya, K.; Zettler, L.W.; Kendon, J.P.; Bidartondo, M.I.; Stice, A.L.; Skarha, S.; Sarasan, V. Preliminary findings on identification of mycorrhizal fungi from diverse orchids in the Central Highlands of Madagascar. Mycorrhiza 2015, 25, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanankierana, H. Personal Communication; Laboratoire de Microbiologie de l’Environnement: Antananarivo, Madagascar, 2018. [Google Scholar]
- Olsson, P.A. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal funghi in soil. FEMS Microbiol. Ecol. 1999, 29, 303–313. [Google Scholar] [CrossRef]
- Salomon, J.N. Le Sud-ouest de Madagascar, Etude de Geographie Physique; Presses Universitaires Bordeaux: Bordeaux, France, 1987; p. 998. [Google Scholar]
- Cabanis, Y.; Chabouis, L.; Chabouis, J. Végétaux et Groupements Végétaux de Madagascar et des Mascareignes; Bureau Pour le Développement de la Production Agricole: Antananarivo, Madagascar, 1987; pp. 1969–1970. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of Total Organic and Available Forms of Phosphorus in Soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Van Aarle, I.M.; Olsson, P.A. Fungal lipid accumulation and development of mycelial structures by two arbuscular mycorrhizal fungi. Appl. Environ. Microbiol. 2003, 69, 6762–6767. [Google Scholar] [CrossRef] [PubMed]
- Blight, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assesssment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A.A. New Method which gives an Objective Measure of Colonization of Roots by Vesicular-Arbuscular Mycorrhizal Fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Mckenney, M. Improved Method for Quantifying Endomycorrhizal Fungi Spores from Soil. Mycologia 1987, 79, 779–782. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 27 August 2018).
- Bradshaw, C.J.A.; Sodhi, N.S.; Brook, B.W. Tropical turmoil: A biodiversity tragedy in progress. Front. Ecol. Environ. 2008, 2, 79–87. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Till, H.M.; Shea, R.W. Biochemistry of deforestation and biomass burning. ACS Symp. Ser. 1992, 483, 428–4556. [Google Scholar]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cairney, J.W.G.; Bastias, B.A. Influences of fire on forest soil fungal communities. Can. J. For. Res. 2007, 37, 207–215. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizas and terrestial ecosystem processes. Ecol. Lett. 2004, 7, 740–754. [Google Scholar] [CrossRef]
- Waeber, P.O.; Wilme, L.; Ramamonjisoa, B.; Garcia, C.; Rakotomalala, D.; Rabemananjara, Z.H.; Sorg, J.P. Dry forests in Madagascar: Neglected and under pressure. Int. For. Rev. 2015, 17, 127–148. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. Nature and Properties of Soils; Prentice Hall-Science: Upper Saddle River, NJ, USA, 2002; p. 960. [Google Scholar]
- Kauffman, J. Biomass and Nutrient Dynamics Associated with Slash Fires in Neotropical Dry Forests. Ecology 1993, 74, 140–151. [Google Scholar] [CrossRef]
- Wang, Q.; Shong, M.; Wang, S. A meta-analysis on the response of microbial biomass, dissolved organic matter, respiration, and N mineralization in mineral soil to fire in forest ecosystems. For. Ecol. Manag. 2012, 271, 91–97. [Google Scholar] [CrossRef]
- De Marco, A.; Gentile, A.E.; Arena, C.; De Santo, A.V. Organic matter, nutrient content and biological activity in burned and unburned soils of a Meditarrenean Maqui area of southern Italy. Int. J. Wildl. Fire 2005, 14, 365–377. [Google Scholar] [CrossRef]
- Garcia-Méndez, G.; Maass, J.M.; Matson, P.A.; Vitousek, P.M. Nitrogen transformations and nitrous oxide flux in a tropical deciduous forest in México. Oecologia 1991, 88, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.I.; Noack, A.G. Black carbon in soils and sediments: Analysis, distribution, implications, and current challenge. Glob. Biogeochem. Cycles 2000, 14, 777–793. [Google Scholar] [CrossRef]
- Ketterings, Q.M.; Birgham, J.M. Soil colour as an indicators of slash-and-burn severity and soil fertility in Sumatra, Indonesia. Soil Sci. Soc. Am. J. 2000, 64, 1826–1833. [Google Scholar] [CrossRef]
- Ellingson, L.J.; Kauffman, J.B.; Cummings, D.L.; Sanford, R.L.; Jaramillo, V.J. Soil N dynamics associated with deforestation, biomass burning and pasture conversion in a Mexican tropical dry forest. For. Ecol. Manag. 2000, 137, 41–51. [Google Scholar] [CrossRef]
- Giardina, C.P.; Sanford, R.L.; Dockersmith, I.C. Changes in soil phosphorus and nitrogen during slash-and-burn clearing of a dry tropical forest. Soil Sci. Soc. Am. J. 2000, 64, 399–405. [Google Scholar] [CrossRef]
- Hammer, E.; Nasr, H.; Wallander, H. Effects of different organic materials and mineral nutrients on arbuscular mycorrhizal fungal growth in a Mediterranean saline dryland. Soil Biol. Biochem. 2011, 43, 2332–2337. [Google Scholar] [CrossRef]
- Olsson, P.A.; Jakobsen, I.; Bååth, E.; Söderström, B. The use of phospholipid and neutral lipid fatty acids to estimate biomass of arbuscular mycorrhizal fungi in soil. Mycol. Res. 1995, 99, 623–629. [Google Scholar] [CrossRef]
- D’Ascoli, R.; Rutigliano, F.A.; De Pascale, R.A.; Gentile, A.; Virzo De Santo, A. Functional diversity of microbial community in Mediterranean maquis soils as affected by fires. Int. J. Wildl. Fire 2005, 14, 355–363. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; De Marco, A.; D’Ascoli, A.R.; Castaldi, S.; Gentile, A.; Virzo De Santo, A. Impact of fire on fungal abundance and microbial efficiency on C assimilation and mineralization in a Mediterranean maquis soil. Biol. Fert. Soils 2007, 44, 377–381. [Google Scholar] [CrossRef]
- Ginzburg, O.; Steinberger, Y. Effects of forest wildfire on soil microbial-community activity and chemical components on a temporal-seasonal scale. Plant Soil. 2012, 360, 243–257. [Google Scholar] [CrossRef]
- Leckie, S.E.; Prescott, C.E.; Grayston, S.J.; Neufeld, J.D.; Mohn, W.W. Characterization of humus microbial communities in adjacent forest types that differ in nitrogen availability. Microbial. Ecol. 2004, 48, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, L.D.; Liu, R.J. Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil 2004, 261, 257–263. [Google Scholar] [CrossRef]
- Sene, G.; Samba-Mbaye, R.; Thiao, M.; Khasa, D.; Kane, A.; Manga, A.; Samba-Mbaye, M.; Sylla, S.N. The abundance and diversity of legume-nodulation rhizobia and arbuscular mycorrhizal fungal communities in soil samples from deforested and man-made forests systems in a semiarid Sahel region in Senegal. Eur. J. Soil Biol. 2012, 52, 30–40. [Google Scholar] [CrossRef]
- Korb, J.E.; Johnson, N.C.; Covington, W.W. Arbuscular mycorrhizal propagule densities respond rapidly to ponderosa pine restoration treatments. J. Appl. Ecol. 2000, 40, 101–111. [Google Scholar] [CrossRef]
- Rosendahl, S. Communities, populations and individuals of arbuscular mycorrhizal fungi. New Phytol. 2008, 178, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattinson, G.S.; Hammill, K.A.; Sutton, B.G.; McGee, P.A. Simulated fire reduces the density of arbuscular mycorrhizal fungi at the soil surface. Mycol. Res. 1999, 103, 491–496. [Google Scholar] [CrossRef]
- Azul, A.M.; Ramos, V.; Sales, F. Early effects of fire on herbaceous vegetation and mycorrhizal symbiosis in high altitude grasslands of Natural Park of Estrela Mountain (PNSE). Symbiosis 2010, 52, 113–123. [Google Scholar] [CrossRef]





| Parameter | Forest | Burned |
|---|---|---|
| pH | 7.02 ± 0.21 a | 7.25 ± 0.19 b |
| Phosphate-Bray (µg/g) | 7.40 ± 3.1 a | 13.1 ± 4.67 b |
| Phosphate-NaFS (µg/g) | 1.01 ± 0.56 a | 2.05 ± 0.9 b |
| NH4 (µg/g) | 2.14 ± 0.32 a | 2.00 ± 0.55 a |
| NO2, NO3 (µg/g) | 6.96 ± 4.8 a | 13.2 ± 7.51 b |
| Total N (µg/g) | 692 ± 233 a | 831 ± 224 b |
| Total C (mg/g) | 6.63 ± 2.28 a | 8.42 ± 2.99 b |
| C:N Ratio | 9.60 ± 0.90 a | 9.95 ± 0.92 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barraclough, A.D.; Olsson, P.A. Slash-and-Burn Practices Decrease Arbuscular Mycorrhizal Fungi Abundance in Soil and the Roots of Didierea madagascariensis in the Dry Tropical Forest of Madagascar. Fire 2018, 1, 37. https://doi.org/10.3390/fire1030037
Barraclough AD, Olsson PA. Slash-and-Burn Practices Decrease Arbuscular Mycorrhizal Fungi Abundance in Soil and the Roots of Didierea madagascariensis in the Dry Tropical Forest of Madagascar. Fire. 2018; 1(3):37. https://doi.org/10.3390/fire1030037
Chicago/Turabian StyleBarraclough, Alícia Donnellan, and Pål Axel Olsson. 2018. "Slash-and-Burn Practices Decrease Arbuscular Mycorrhizal Fungi Abundance in Soil and the Roots of Didierea madagascariensis in the Dry Tropical Forest of Madagascar" Fire 1, no. 3: 37. https://doi.org/10.3390/fire1030037
APA StyleBarraclough, A. D., & Olsson, P. A. (2018). Slash-and-Burn Practices Decrease Arbuscular Mycorrhizal Fungi Abundance in Soil and the Roots of Didierea madagascariensis in the Dry Tropical Forest of Madagascar. Fire, 1(3), 37. https://doi.org/10.3390/fire1030037