


Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
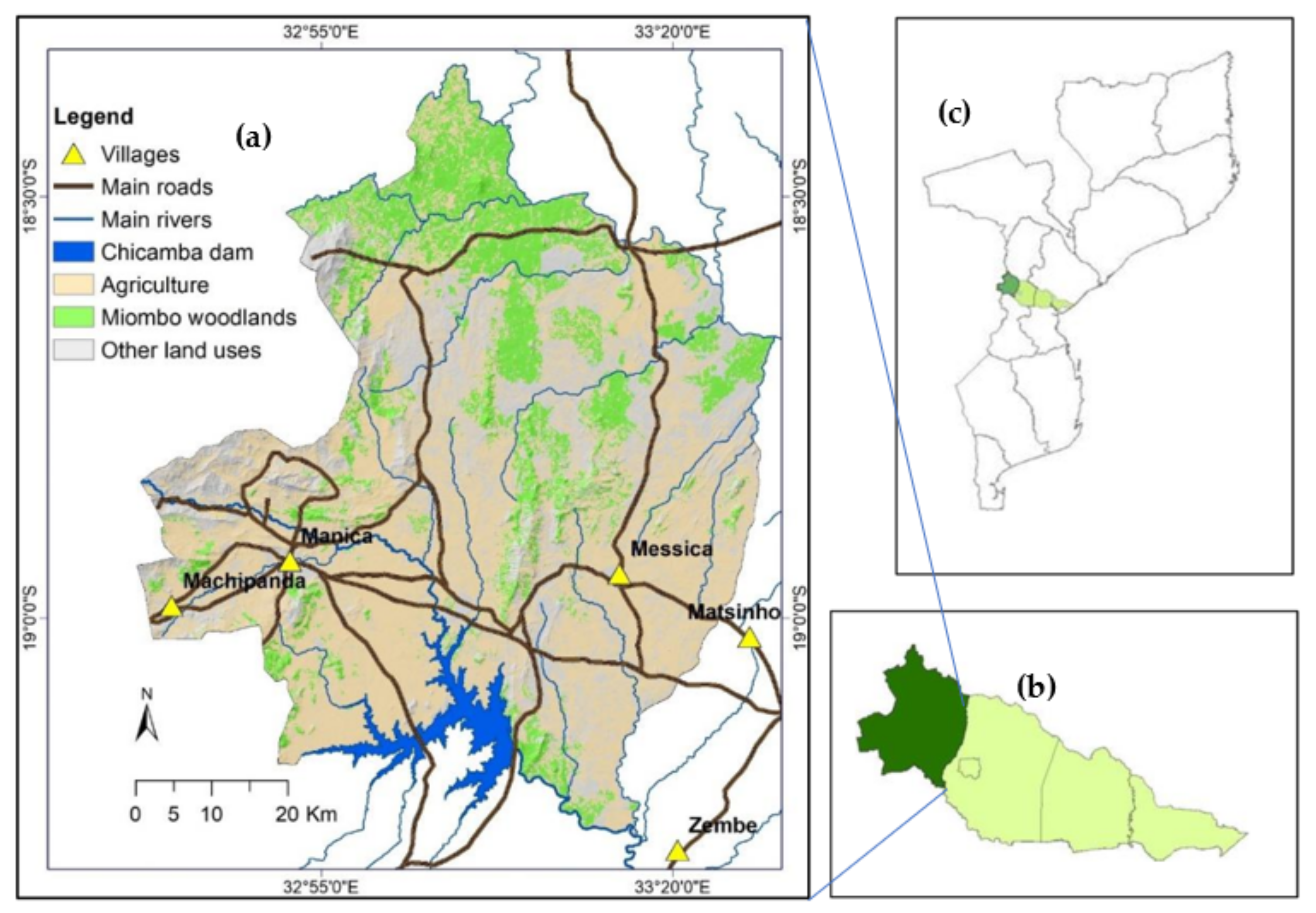
2.1. Study Area Description

2.2. Data Acquisition and Fire Mapping
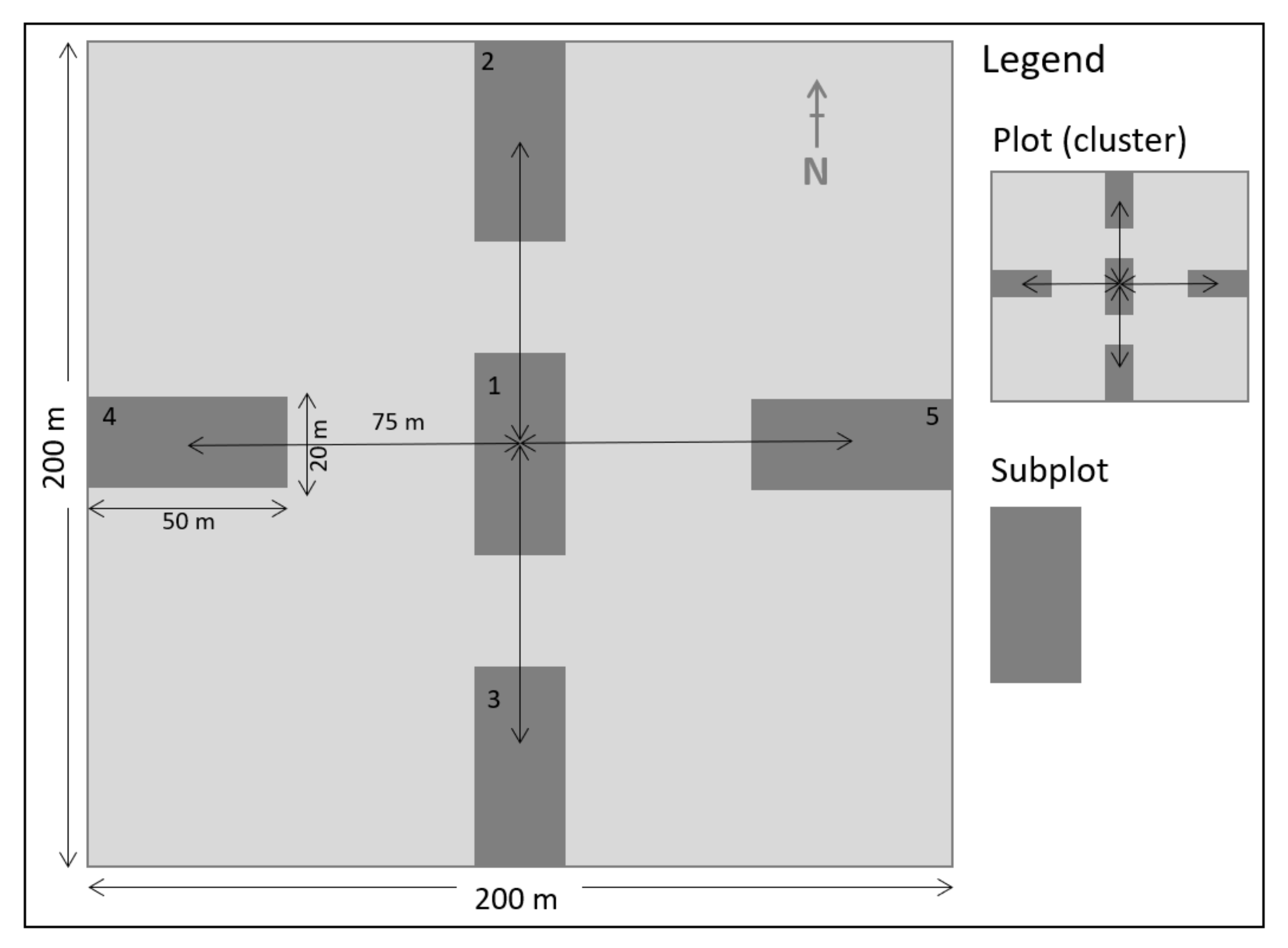
2.3. Field Data Collection
2.4. Data Analysis
3. Results
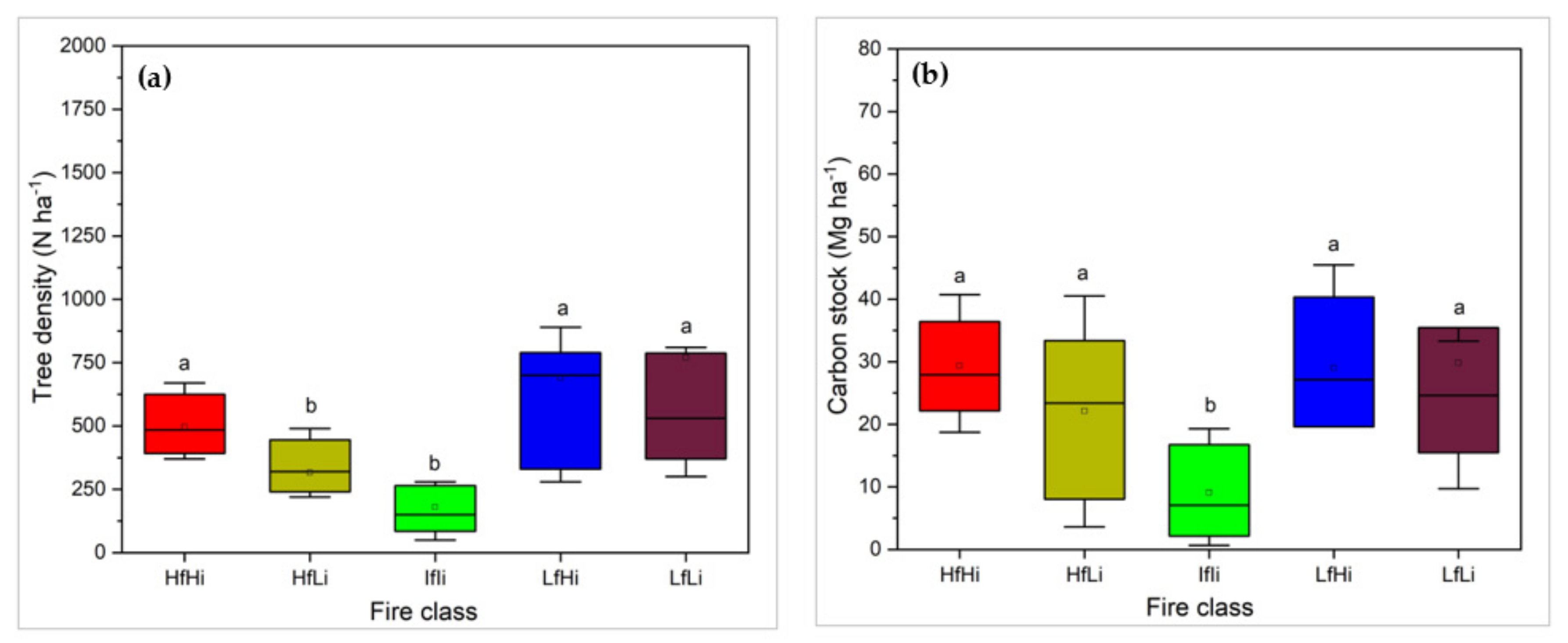
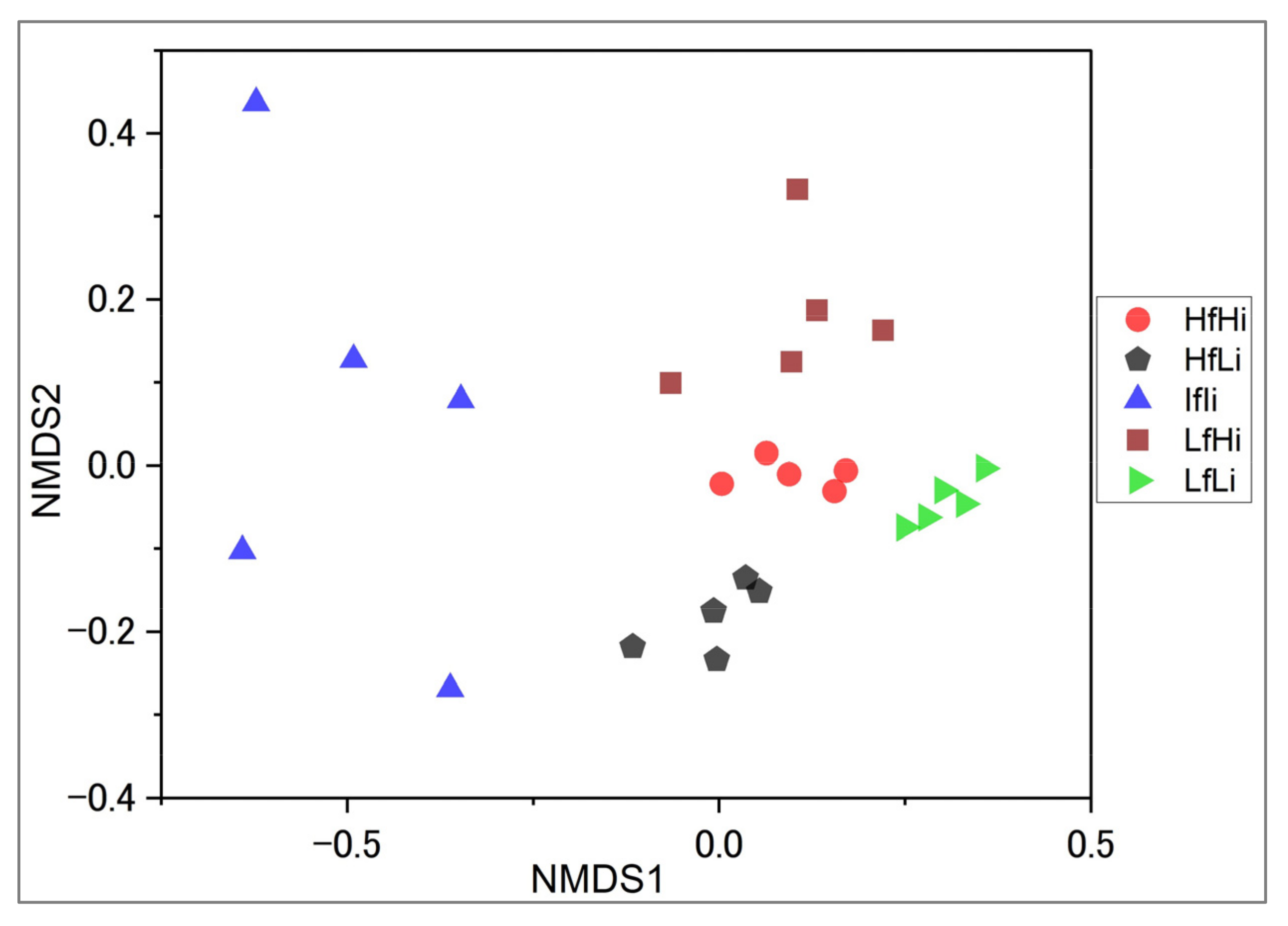
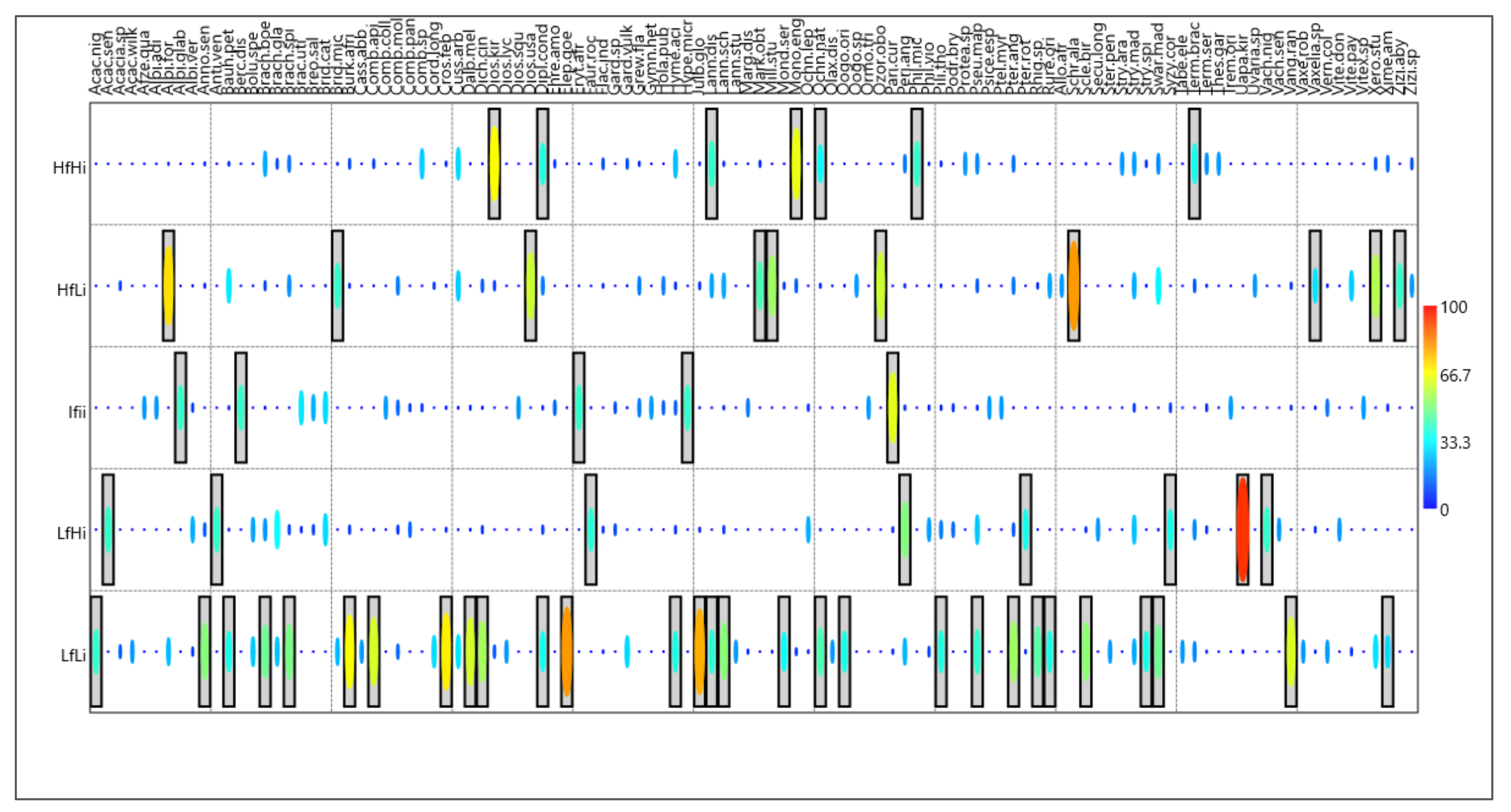
3.1. Tree Species Composition and Structure across Fire Regimes
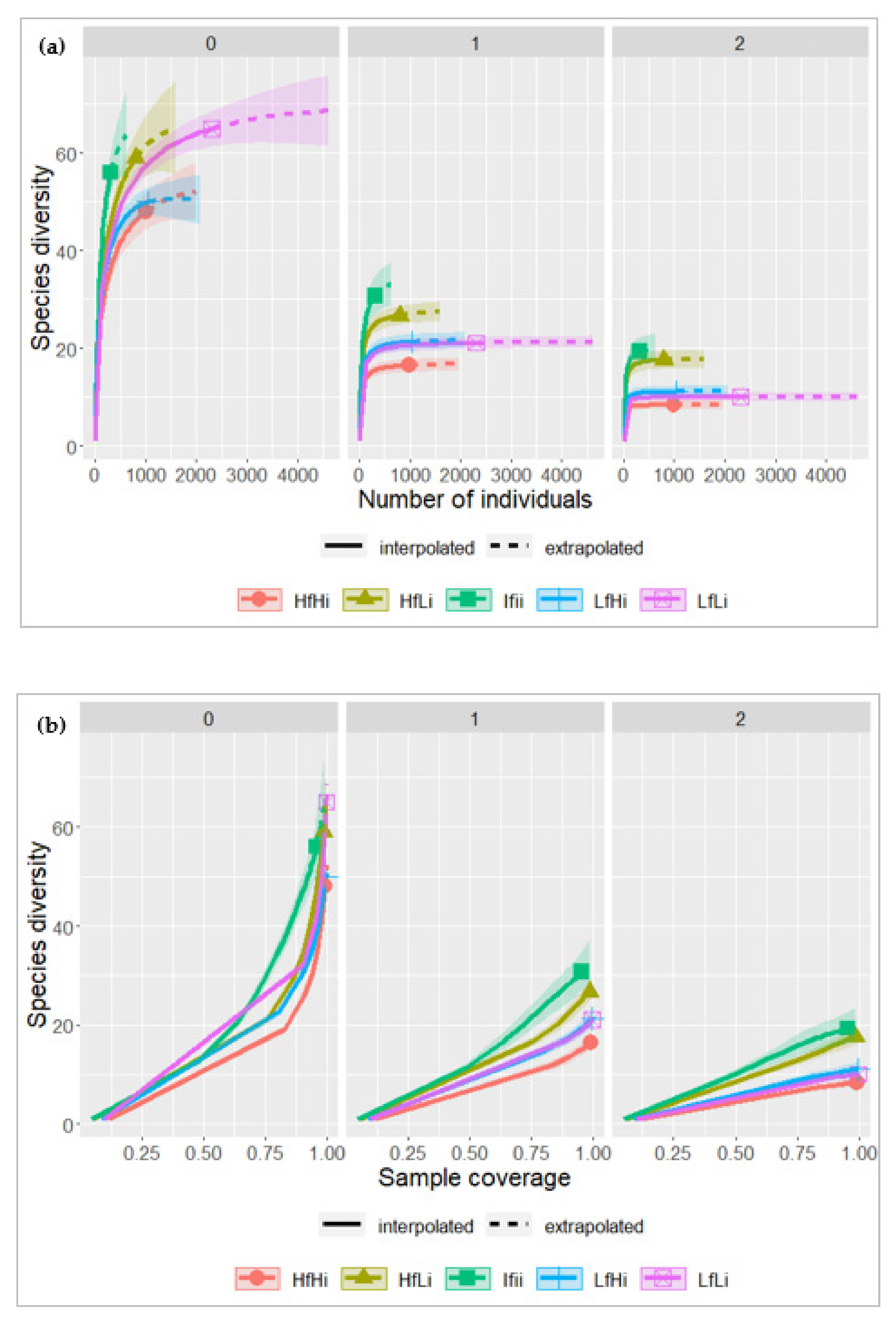
3.2. Tree Species Diversity
3.3. Tree Species Importance Value across Fire Regimes
4. Discussion
4.1. Effects of Fire Regime on Woody Structure and Composition
4.2. Tree Species Diversity
4.3. Tree Species Importance Value across Fire Regimes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bond, W.J.; Keeley, J.E. Fire as a Global “Herbivore”: The Ecology and Evolution of Flammable Ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, C.E.R.; Anderson, T.M.; Sankaran, M.; Higgins, S.I.; Archibald, S.; Hoffmann, W.A.; Hanan, N.P.; Williams, R.J.; Fensham, R.J.; Felfili, J.; et al. Savanna Vegetation-Fire-Climate Relationships Differ Among Continents. Science 2014, 343, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Sharples, J.J. Review of formal methodologies for wind—slope correction of wildfire rate of spread. Int. J. Wildland Fire 2008, 17, 179–193. [Google Scholar] [CrossRef]
- Lentile, L.B.; Holden, Z.A.; Smith, A.M.S.; Falkowski, M.J.; Hudak, A.T.; Morgan, P.; Lewis, S.A.; Gessler, P.E.; Benson, N.C. Remote sensing techniques to assess active fire characteristics and post-fire effects. Int. J. Wildland Fire 2006, 15, 319–345. [Google Scholar] [CrossRef]
- Miettinen, J.; Liew, S.C. Connection between fire and land cover change in Southeast Asia: A remote sensing case study in Riau, Sumatra. Int. J. Remote Sens. 2005, 26, 1109–1126. [Google Scholar] [CrossRef]
- Whitlock, C.; Higuera, P.E.; Mcwethy, D.B.; Briles, C.E. Paleoecological Perspectives on Fire Ecology: Revisiting the Fire-Regime Concept. Open Ecol. J. 2010, 3, 6–23. [Google Scholar] [CrossRef] [Green Version]
- Krebs, P.; Pezzatti, G.B.; Mazzoleni, S.; Talbot, L.M.; Conedera, M. Fire regime: History and definition of a key concept in disturbance ecology. Theory Biosci. 2010, 129, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Keeley, J.E. A Burning Story: The Role of Fire in the History of Life. BioScience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of Fire Regime and Habitat on Tree Dynamics in North Florida Longleaf Pine Savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar] [CrossRef]
- Gandiwa, E. Effects of Repeated Burning on Woody Vegetation Structure and Composition in a Semi-Arid Southern Africa Savanna. Int. J. Environ. Sci. 2011, 2, 458–471. [Google Scholar]
- Frost, P. The Ecology of Miombo Woodlands. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B., Ed.; Centre for International Forestry Research (CIFOR): Bogor, Indonesia, 1996; pp. 11–58. [Google Scholar]
- Eva, H.; Lambin, E. Fires and Land-Cover Change in the Tropics: A Remote Sensing Analysis at the Landscape Scale. J. Biogeogr. 2000, 27, 765–776. Available online: http://www.Jstor.Org/Stable/2656223 (accessed on 27 November 2019). [CrossRef]
- Chuvieco, E.; Mouillot, F.; van der Werf, G.R.; San Miguel, J.; Tanase, M.; Koutsias, N.; García, M.; Yebra, M.; Padilla, M.; Gitas, I.; et al. Historical background and current developments for mapping burned area from satellite Earth observation. Remote Sens. Environ. 2019, 225, 45–64. [Google Scholar] [CrossRef]
- Viedma, O.; Moreno, J.M.; Rieiro, I. Interactions between land use/land cover change, forest fires and landscape structure in Sierra de Gredos (Central Spain). Environ. Conserv. 2006, 33, 212–222. [Google Scholar] [CrossRef]
- Flasse, S.P.; Trigg, S.N.; Ceccato, P.N.; Perryman, A.H.; Hudak, A.T.; Thompson, M.W.; Brockett, B.H.; Drame, M.; Ntabeni, T.; Frost, T.E.; et al. Remote Sensing of Vegetation Fires and Its Contribution to a Fire Management Information System. In Wildland Fire Management Handbook For Sub-Sahara Africa; Goldammer, d.R., Ed.; Global Fire Monitoring Center (GFMC): Mbombela, South Africa, 2004; ISBN 191983365X. [Google Scholar]
- Archibald, S.; Scholes, R.; Roy, D.P.; Roberts, G.; Boschetti, L. Southern African fire regimes as revealed by remote sensing. Int. J. Wildland Fire 2010, 19, 861–878. [Google Scholar] [CrossRef] [Green Version]
- Dziba, L.; Ramoelo, A.; Ryan, R.; Harrison, S.; Pritchard, R.; Tripathi, H.; Sitas, N.; Selomane, O.; Engelbrecht, F.; Pereira, L.; et al. Biogeography and Ecology of Miombo Woodlands. In MW in a CES the R and S of P and Woodlands; Ribeiro, N.S., Katerere, Y., Chirwa, P.W., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Moura, I.; Maquia, I.; Rija, A.A.; Ribeiro, N.; Ribeiro-Barros, A.I. Biodiversity Studies in Key Species from the African Mopane and Miombo Woodlands. In Genetic Diversity; Bitz, L., Ed.; IntechOpen: London, UK, 2017; pp. 91–109. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, N.S.; Matos, C.N.; Moura, I.R.; Washington-Allen, R.A.; Ribeiro, A.I. Monitoring vegetation dynamics and carbon stock density in miombo woodlands. Carbon Balance Manag. 2013, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.; Frost, P.; Byron, N. Miombo Woodlands and Their Use: Overview and Key Issues. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B.M., Ed.; Center for International Forestry Research: Bogor, Indonesia, 1996; pp. 1–10. ISBN 9798764072. [Google Scholar]
- Saito, M.; Luyssaert, S.; Poulter, B.; Williams, M.; Ciais, P.; Bellassen, V.; Ryan, C.M.; Yue, C.; Cadule, P.; Peylin, P. Fire regimes and variability in aboveground woody biomass in miombo woodland. J. Geophys. Res. Biogeosci. 2014, 119, 1014–1029. [Google Scholar] [CrossRef] [Green Version]
- Dewees, P.A.; Campbell, B.M.; Katerere, Y.; Sitoe, A.; Cunningham, A.B.; Angelsen, A.; Wunder, S. Managing the Miombo Woodlands of Southern Africa: Policies, Incentives and Options for the Rural Poor. J. Nat. Resour. Policy Res. 2010, 2, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Chidumayo, E.N.; Timberlake, J.; Sawadogo, L. Distribution and Characteristics of African Dry Forests and Woodlands. In The Dry Forests and Woodlands of Africa: Managing for Products and Services; Chidumayo, E.N., Gumbo, D.J., Eds.; Earthscan: London, UK, 2010; pp. 11–41. [Google Scholar]
- Syampungani, S.; Geldenhuys, C.J.; Chirwa, P.W. Regeneration dynamics of miombo woodland in response to different anthropogenic disturbances: Forest characterisation for sustainable management. Agrofor. Syst. 2015, 90, 563–576. [Google Scholar] [CrossRef]
- Chidumayo, E. Development of Brachystegia-Julbernardia woodland after clear-felling in central Zambia: Evidence for high resilience. Appl. Veg. Sci. 2004, 7, 237–242. [Google Scholar] [CrossRef]
- Tarimo, B.; Dick, Ø.B.; Gobakken, T.; Totland, Ø. Spatial distribution of temporal dynamics in anthropogenic fires in miombo savanna woodlands of Tanzania. Carbon Balance Manag. 2015, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, E.E.; Moss, P.; Haseler, M.; Maron, M. The influence of a variable fire regime on woodland structure and composition. Int. J. Wildland Fire 2015, 24, 59–69. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Bogenrieder, A.; Stahr, K.; Bösch, R.; van der Heijden, M.; Sieverding, E. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol. Biochem. 2010, 42, 724–738. [Google Scholar] [CrossRef]
- Makumbe, P.; Chikorowondo, G.; Dzamara, P.C.; Ndaimani, H.; Gandiwa, E. Effects of Fire Frequency on Woody Plant Composition and Functional Traits in a Wet Savanna Ecosystem. Int. J. Ecol. 2020, 2020, 1672306. [Google Scholar] [CrossRef]
- Steel, Z.L.; Safford, H.D.; Viers, J.H. The fire frequency-severity relationship and the legacy of fire suppression in California forests. Ecosphere 2015, 6, 1–23. [Google Scholar] [CrossRef]
- Trapnell, C.G. Ecological Results of Woodland and Burning Experiments in Northern Rhodisia. J. Ecol. 1959, 47, 129. [Google Scholar] [CrossRef]
- Bond, W.J.; Van Wilgen, B.W. Fire and Plants; Population and Community Biology Series 14; Chapman & Hall: London, UK, 1996; ISBN 9789401057721. [Google Scholar]
- Miller, J.E.D.; Safford, H.D. Are plant community responses to wildfire contingent upon historical disturbance regimes? Glob. Ecol. Biogeogr. 2020, 29, 1621–1633. [Google Scholar] [CrossRef]
- Jinga, P.; Ashley, M.V. Climate change threatens some miombo tree species of sub-Saharan Africa. Flora 2019, 257, 151421. [Google Scholar] [CrossRef]
- Jinga, P.; Palagi, J. Dry and wet miombo woodlands of south-central Africa respond differently to climate change. Environ. Monit. Assess. 2020, 192, 372. [Google Scholar] [CrossRef]
- Ribeiro, N.; Ruecker, G.; Govender, N.; Macandza, V.; Pais, A.; Machava, D.; Chauque, A.; Lisboa, S.N.; Bandeira, R. The influence of fire frequency on the structure and botanical composition of savanna ecosystems. Ecol. Evol. 2019, 9, 8253–8264. [Google Scholar] [CrossRef]
- Foster, C.N.; Barton, P.S.; MacGregor, C.I.; Catford, J.A.; Blanchard, W.; Lindenmayer, D.B. Effects of fire regime on plant species richness and composition differ among forest, woodland and heath vegetation. Appl. Veg. Sci. 2018, 21, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.D.; van Wilgen, B.W.; Burns, C.E.; Govender, N.; Potgieter, A.L.F.; Andelman, S.; Biggs, H.C.; Botha, J.; Trollope, W.S.W. Long-term effects of fire frequency and season on herbaceous vegetation in savannas of the Kruger National Park, South Africa. J. Plant Ecol. 2013, 6, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Governo de Moçambique. Inventário Florestal Nacional: Relatório Final; Ministério da Terra, Ambiente e Desenvolvimento Rural (MITADER): Maputo, Moçambique, 2018; 118p. [Google Scholar]
- Governo de Moçambique. Plano de Acção, Prevenção e Controlo às Queimadas Descontroladas (2008-2018); MICOA: Maputo, Moçambique, 2007; 53p. [Google Scholar]
- Ministério da Administração Estatal (Ed.) Perfil do Distrito de Muanza, Província de Sofala; MAE, Edição: Maputo, Moçambique, 2005; 60p. [Google Scholar]
- Instituto Nacional de Estatística (INE). IV Recenseamento Geral da População e Habitação; INE: Maputo, Moçambique, 2017. [Google Scholar]
- Marzoli, A. Inventário Florestal Nacional—Avaliação Integrada das Florestas de Moçambique (AIFM); DNTF, MINAG: Maputo, Mozambique, 2007. [Google Scholar]
- Giglio, L.; Boschetti, L.; Roy, D.P.; Humber, M.L.; Justice, C.O. The Collection 6 MODIS burned area mapping algorithm and product. Remote Sens. Environ. 2018, 217, 72–85. [Google Scholar] [CrossRef]
- Giglio, L.; Schroeder, W.; Hall, J.V.; Justice, C.O. MODIS Collection 6 Active Fire Product User ’s Guide Revision B. Dep. Geogr. Sci. Univ. Md. 2020, 9. [Google Scholar]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Rogan, J.; Franklin, J. Mapping Wildfire Burn Severity in Southern California Forests and Shrublands Using Enhanced Thematic Mapper Imagery. Geocarto Int. 2001, 16, 91–106. [Google Scholar] [CrossRef]
- Gandiwa, E.; Kativu, S. Influence of Fire Frequency on Colophospermum Mopane and Combretum Apiculatum Woodland Structure and Composition in Northern Gonarezhou National Park, Zimbabwe. Koedoe 2009, 51, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Achilleos, G.A. The Inverse Distance Weighted interpolation method and error propagation mechanism—Creating a DEM from an analogue topographical map. J. Spat. Sci. 2011, 2, 283–304. [Google Scholar] [CrossRef]
- Devkota, J.U. Statistical analysis of active fire remote sensing data: Examples from South Asia. Environ. Monit. Assess. 2021, 193, 608. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Estimating Species Richness. In Biological Diversity: Frontiers in Measurement and Assessment; Oxford University Press: Oxford, UK, 2011; pp. 39–54. [Google Scholar]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Wiser, S.K.; Brotons, L. Using species combinations in indicator value analyses. Methods Ecol. Evol. 2012, 3, 973–982. [Google Scholar] [CrossRef]
- Curtis, J.T.; Mcintosh, R.P. The Interrelations of Certain Analytic and Synthetic Phytosociological Characters. Ecology 1950, 31, 434–455. Available online: http://Www.Jstor.Org/Stable/1931497 (accessed on 16 November 2021). [CrossRef]
- Guedes, B.S.; Sitoe, A.A.; Olsson, B.A. Allometric models for managing lowland miombo woodlands of the Beira corridor in Mozambique. Glob. Ecol. Conserv. 2018, 13, e00374. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Cauldwell, A.E.; Zieger, U. A reassessment of the fire-tolerance of some miombo woody species in the Central Province, Zambia. Afr. J. Ecol. 2000, 38, 138–146. [Google Scholar] [CrossRef]
- Peters, C.M. Sustainable Harvest of Non-Timber Plant Resources in Tropical Moist Forest: An Ecological Primer; Biodiversity Support Program c/o World Wildlife Fund: Washington, DC, USA, 1994; 45p. [Google Scholar]
- Balch, J.K.; Massad, T.J.; Brando, P.; Nepstad, D.C.; Curran, L.M. Effects of high-frequency understorey fires on woody plant regeneration in southeastern Amazonian forests. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120157. [Google Scholar] [CrossRef] [Green Version]
- Ryan, C.M.; Williams, M. How does fire intensity and frequency affect miombo woodland tree populations and biomass? Ecol. Appl. 2011, 21, 48–60. [Google Scholar] [CrossRef]
- Jew, E.K.K.; Dougill, A.J.; Sallu, S.M.; Connell, J.O.; Benton, T.G. Forest Ecology and Management Miombo Woodland under Threat: Consequences for Tree Diversity and Carbon Storage. For. Ecol. Manag. 2016, 361, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Olsson, L.; Barbosa, H.; Bhadwal, S.; Cowie, A.; Delusca, K.; Flores-Renteria, D.; Hermans, K.; Jobbagy, E.; Kurz, W.; Li, D.; et al. Land Degradation. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo, E., Buendia, V., Masson, D., Eds.; IPCC: Geneva, Switzerland, 2019; pp. 345–436. [Google Scholar]
- Thornley, J.H.M.; Cannell, M.G.R. Long-term effects of fire frequency on carbon storage and productivity of boreal forests: A modeling study. Tree Physiol. 2004, 24, 765–773. [Google Scholar] [CrossRef]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J.; Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in Miombo woodlands of Zambia. For. Ecol. Manag. 2013, 304, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, N.S.; Shugart, H.H.; Washington-Allen, R. The effects of fire and elephants on species composition and structure of the Niassa Reserve, northern Mozambique. For. Ecol. Manag. 2008, 255, 1626–1636. [Google Scholar] [CrossRef]
- Chirwa, P.W.; Akinnifesi, F.K. Ecology and Biology of Uapaca Kirkiana, Strychnos Cocculoides and Scelerocarya Birrea in Southern Africa. In Indigenous Fruit in the tropics: Domestication, Utilisation and Commercialisation; Akinnifesi, F.K., Leakey, R.R.B., Ajayi, O.C., Sileshi, G., Tchoundjeu, Z., Matakala, P., Kwesiga, F.R., Eds.; CABI: Egham, UK, 2008; p. 322. [Google Scholar]
- Chawafambira, A.; Sedibe, M.M.; Mpofu, A.; Achilonu, M. Uapaca kirkiana, an indigenous fruit tree in sub-Saharan Africa: A comprehensive review. Cogent Food Agric. 2020, 6, 1766735. [Google Scholar] [CrossRef]
- Richter, C.; Rejmánek, M.; Miller, J.E.D.; Welch, K.R.; Weeks, J.; Safford, H. The species diversity × fire severity relationship is hump-shaped in semiarid yellow pine and mixed conifer forests. Ecosphere 2019, 10, e02882. [Google Scholar] [CrossRef] [Green Version]
- Neeraja, U.V.; Rajendrakumar, S.; Saneesh, C.S.; Dyda, V.; Knight, T.M. Fire alters diversity, composition, and structure of dry tropical forests in the Eastern Ghats. Ecol. Evol. 2021, 11, 6593–6603. [Google Scholar] [CrossRef]
- Kadmon, R.; Benjamini, Y. Effects of Productivity and Disturbance on Species Richness: A Neutral Model. Am. Nat. 2006, 167, 939–946. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs High Diversity of Trees and Corals Is Maintained. Science 1978, 199, 1302–1310. Available online: http://www.jstor.org/stable/1745369 (accessed on 1 October 2022). [CrossRef] [Green Version]
- Tulloch, A.I.T.; Pichancourt, J.-B.; Gosper, C.R.; Sanders, A.; Chadès, I. Fire management strategies to maintain species population processes in a fragmented landscape of fire-interval extremes. Ecol. Appl. 2016, 26, 2175–2189. [Google Scholar] [CrossRef] [Green Version]
- Sheil, D.; Burslem, D.F. Disturbing hypotheses in tropical forests. Trends Ecol. Evol. 2003, 18, 18–26. [Google Scholar] [CrossRef]
- Bongers, F.; Poorter, L.; Hawthorne, W.D.; Sheil, D. The intermediate disturbance hypothesis applies to tropical forests, but disturbance contributes little to tree diversity. Ecol. Lett. 2009, 12, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, H.M.; Mallik, A.U. Predicting Plant Diversity Response to Disturbance: Applicability of the Intermediate Disturbance Hypothesis and Mass Ratio Hypothesis. Crit. Rev. Plant Sci. 2013, 32, 383–395. [Google Scholar] [CrossRef]
- Guitet, S.; Sabatier, D.; Brunaux, O.; Couteron, P.; Denis, T.; Freycon, V.; Gonzalez, S.; Hérault, B.; Jaouen, G.; Molino, J.-F.; et al. Disturbance Regimes Drive The Diversity of Regional Floristic Pools Across Guianan Rainforest Landscapes. Sci. Rep. 2018, 8, 3872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNicol, I.M.; Ryan, C.M.; Williams, M. How resilient are African woodlands to disturbance from shifting cultivation? Ecol. Appl. 2015, 25, 2320–2336. [Google Scholar] [CrossRef]
- Hofiço, A.S.; Costa, E.A.; Fleig, F.D.; José, S.; Nanvonamuquitxo, A. Regulation of the Diametric Structure of the Miombo Woodland Using the De Liocourt Method in Mozambique Regulação Da Estrutura Diamétrica de Uma Floresta de Miombo Usando o Método De Liocourt Em Moçambique. Nativa 2018, 6, 407–414. [Google Scholar] [CrossRef]
- Maquia, I.; Ribeiro, N.S.; Silva, V.; Bessa, F.; Goulao, L.F.; Ribeiro, A.I. Genetic diversity of Brachystegia boehmii Taub. and Burkea africana Hook. f. across a fire gradient in Niassa National Reserve, northern Mozambique. Biochem. Syst. Ecol. 2013, 48, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Malimbwi, R.E.; Zahabu, E.; Monela, G.C.; Misana, S.; Jambiya, G.C.; Mchome, B. Charcoal Potential of Miombo Woodlands at Kitulangalo, Tanzania. J. Trop. For. Sci. 2005, 17, 197–210. Available online: http://www.jstor.org/stable/23616567 (accessed on 8 September 2022).
- CEAGRE & Winrock-International. Agentes e Causas de Desmatamento e Degradação Florestal em Moçambique; CEAGRE-UEM and Winrock International: Maputo, Moçambique, 2016; pp. 1–36. [Google Scholar]
- Gonçalves, F.M.P.; Revermann, R.; Gomes, A.L.; Aidar, M.P.M.; Finckh, M.; Juergens, N. Tree Species Diversity and Composition of Miombo Woodlands in South-Central Angola: A Chronosequence of Forest Recovery after Shifting Cultivation. Int. J. For. Res. 2017, 2017, 6202093. [Google Scholar] [CrossRef] [Green Version]
- Stahle, D.; Mushove, P.; Cleaveland, M.; Roig, F.; Haynes, G. Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. For. Ecol. Manag. 1999, 124, 217–229. [Google Scholar] [CrossRef]
- Geldenhuys, C.J. The effect of different regimes of annual burning on two woodland communities in kavango/die invloed van verskillende stelsels van jaarlikse brand op twee boomveldgemeenskappe in kavango. S. Afr. For. J. 1977, 103, 32–42. [Google Scholar] [CrossRef]
- Banda, T.; Schwartz, M.W.; Caro, T. Effects of fire on germination of Pterocarpus angolensis. For. Ecol. Manag. 2006, 233, 116–120. [Google Scholar] [CrossRef]
- Mojeremane, W.; Lumbile, A.U. A Review of Pterocarpus angolensis DC. (Mukwa) an Important and Threatened Timber Species of the Miombo Woodlands. Res. J. For. 2016, 10, 8–14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stratum | Code | Description |
|---|---|---|
| High-frequency vs. High-intensity | HfHi | Sites usually occur in isolated mature forest fragments with limited human access and high seasonal fuel accumulation, consisting mainly of abundant grasses. |
| High-frequency vs. Low-intensity | HfLi | Combination of mature woodlands and multiple land uses, characterized by intense human activity such as charcoal production, agriculture, and considerable human presence and settlements. |
| Intermediate-frequency vs. Intermediate-intensity | IfIi | Transition areas between mature forest and fallow land. The presence of specific human interference (extraction of poles, firewood and wood) distinguishes this stratum from others. The local communities occasionally use these areas for charcoal and crop production. |
| Low-frequency vs. Low-intensity | LfLi | Abandoned agricultural areas (regrowth stands), with early recovery stages of miombo woodlands (10–15-year-old). |
| Low-frequency vs. High-intensity | LfHi | Similar to LfLi sites but with the natural succession process in a more advanced stage (20–25-year-old regrowth stands) and high seasonal fuel accumulation, consisting mainly of abundant grasses. |
| Species | Family | IVI | ||||
|---|---|---|---|---|---|---|
| HfHi | HfLi | IfIi | LfHi | LfLi | ||
| Brachystegia boehmii | Fabaceae | 74.7 | 47.0 | 31.8 | 60.7 | 75.8 |
| Diplorhynchus condylocarpon | Apocynaceae | 26.5 | 15.8 | 11.3 | 10.8 | 14.2 |
| Pseudolachnostylis maprouneifolia | Phyllanthaceae | 22.2 | 19.8 | 18.2 | 28.0 | 21.5 |
| Pterocarpus angolensis | Fabaceae | 15.7 | 9.8 | 7.2 | 11.0 | 14.0 |
| Swartzia madagascariensis | Fabaceae | 13.6 | 20.3 | - | - | 15.2 |
| Pericopsis angolensis | Fabaceae | 12.6 | - | 14.3 | 20.3 | 9.4 |
| Strychnos madagascariensis | Strychnaceae | 12.3 | 12.1 | 16.2 | 13.2 | - |
| Terminalia brachystemma | Combretaceae | 11.0 | - | - | 9.3 | - |
| Lannea discolor | Anacardiaceae | 10.7 | - | - | - | - |
| Hymenocardia acida | Phyllanthaceae | 9.4 | - | 17.2 | - | 7.1 |
| Brachystegia spiciformis | Fabaceae | - | 8.5 | - | - | 9.3 |
| Albizia forbesii | Fabaceae | - | 15.8 | - | - | - |
| Xeroderris stuhlmannii | Fabaceae | - | 14.4 | - | - | - |
| Bauhinia petersiana | Fabaceae | - | 9.8 | - | - | - |
| Parinari curatellifolia | Chrysobalanaceae | - | - | 34.4 | - | - |
| Albizia glaberrima | Fabaceae | - | - | 15.4 | - | - |
| Julbernardia globiflora | Fabaceae | - | - | 7.2 | - | 20.3 |
| Uapaca kirkiana | Phyllanthaceae | - | - | - | 14.2 | - |
| Piliostigma thonningii | Fabaceae | - | - | - | 8.6 | - |
| Pterocarpus rotundifolius | Fabaceae | - | - | - | 8.0 | - |
| Crossopteryx febrifuga | Rubiaceae | - | - | - | - | 7.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buramuge, V.A.; Ribeiro, N.S.; Olsson, L.; Bandeira, R.R.; Lisboa, S.N. Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique. Fire 2023, 6, 26. https://doi.org/10.3390/fire6010026
Buramuge VA, Ribeiro NS, Olsson L, Bandeira RR, Lisboa SN. Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique. Fire. 2023; 6(1):26. https://doi.org/10.3390/fire6010026
Chicago/Turabian StyleBuramuge, Victorino Américo, Natasha Sofia Ribeiro, Lennart Olsson, Romana Rombe Bandeira, and Sá Nogueira Lisboa. 2023. "Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique" Fire 6, no. 1: 26. https://doi.org/10.3390/fire6010026
APA StyleBuramuge, V. A., Ribeiro, N. S., Olsson, L., Bandeira, R. R., & Lisboa, S. N. (2023). Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique. Fire, 6(1), 26. https://doi.org/10.3390/fire6010026