
Influence of Fires on Desert Plant Communities at the Chernye Zemli (SW Russia)

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
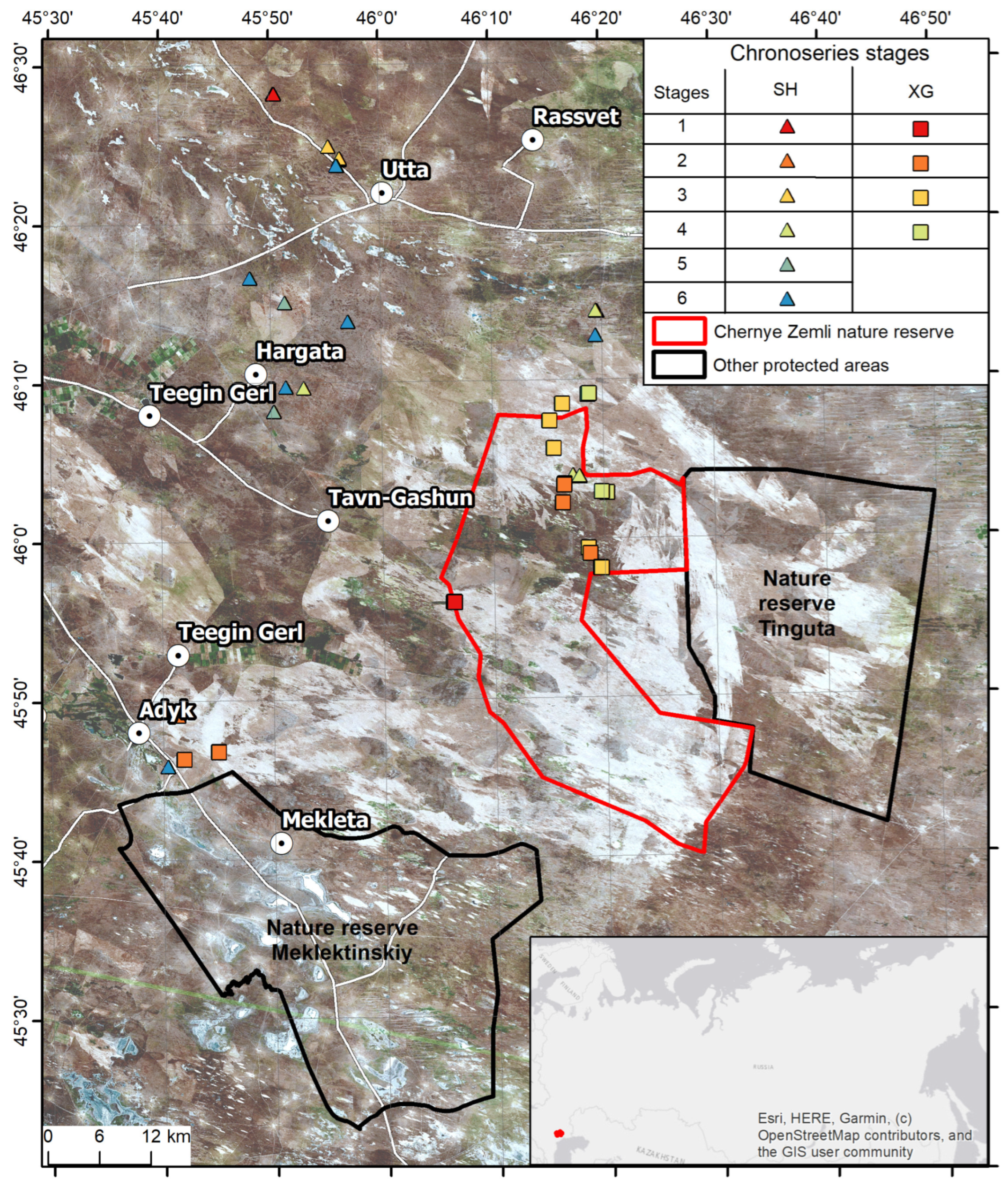
2.1. Characteristics of the Study Area
2.2. Selection of Key Sites for Field Work
2.3. Field Work
2.4. Data Processing
3. Results
3.1. Comparative Analysis of Plant Communities in the “Chernye Zemli” Reserve and Beyond
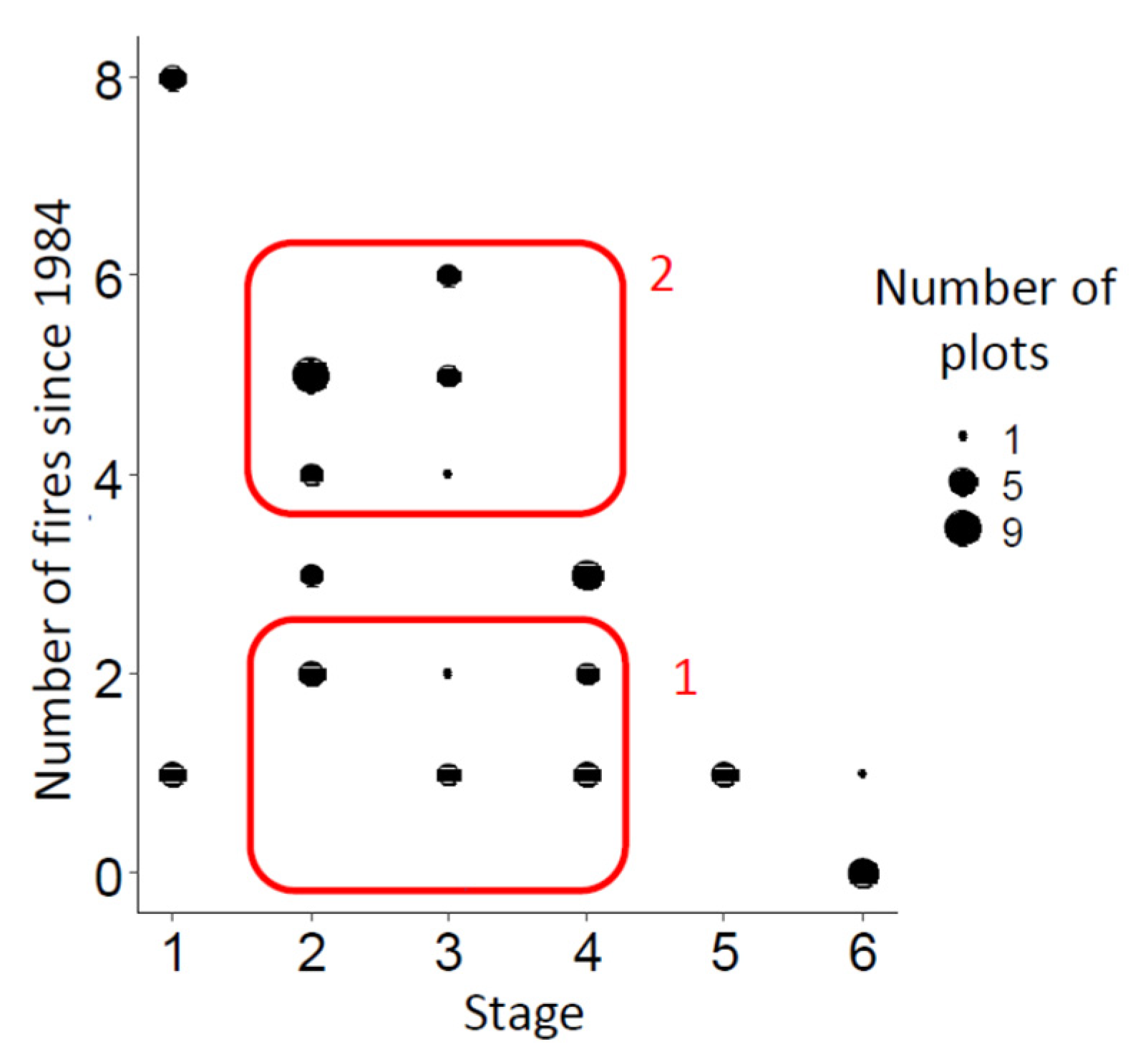
3.2. Comparative Analysis of Plant Communities of the “Chernye Zemli” at Different Stages of Pyrogenic Succession
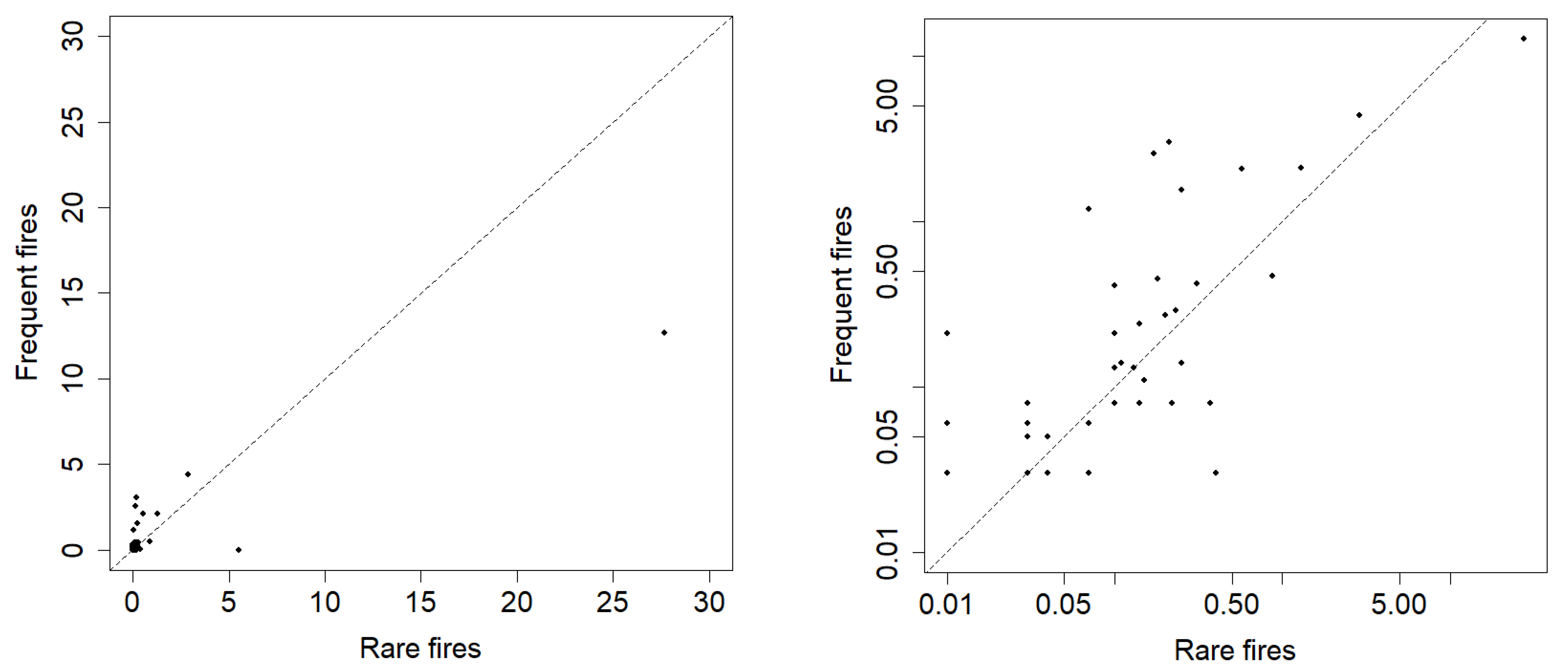
3.3. Assessment of the Impact of Fire Frequency on Plant Communities of the “Chernye Zemli” Nature Reserve
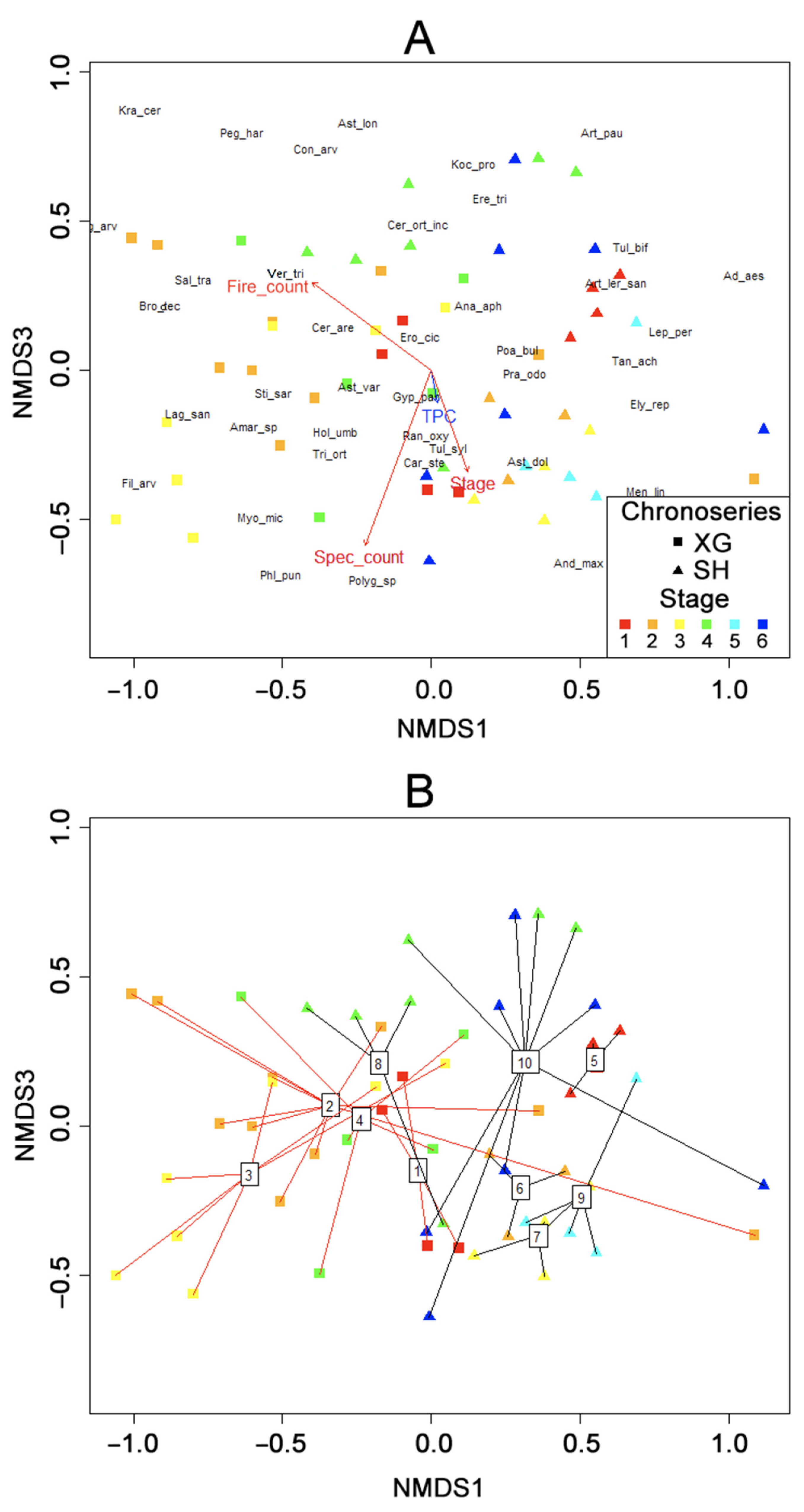
3.4. Non-Metrical Multidimensional Scaling
4. Discussion
- (a)
- For some species, conditions are not optimal both before and after the fire, so their projective cover is low both in the pre-fire and post-fire stages;
- (b)
- During a fire, individuals of species with higher prevalence are more likely to be found in loci less affected by open fire and high temperatures, and thus survive;
- (c)
- After a fire, seeds invade from adjacent territories, and the supply of diaspores is higher for species with a higher number of individuals (vicinism according to [48]);
- (d)
- For the introduction of some minor species, an important role can be played by the primary plant cover, which can provide partial protection of seedlings from negative environmental factors due to shading or moisture retention in the accumulated dead litter in the spring [16]; therefore, such plants cannot appear in the early stages of pyrogenic succession.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubinin, M.Y.; Lushchekina, A.A.; Radelof, F.K. Assessment of the current dynamics of fires in arid ecosystems based on space survey materials (on the example of Black lands). Arid Ecosyst. 2010, 16, 5–16. [Google Scholar]
- Karpachevskiy, A.; Lednev, S.; Semenkov, I.; Sharapova, A.; Nagiyev, S.; Koroleva, T. Delineation of burned arid landscapes using Landsat 8 OLI data: A case study of Karaganda region in Kazakhstan. Arid L Res. Manag. 2021, 35, 292–310. [Google Scholar] [CrossRef]
- Shinkarenko, S.S.; Ivanov, N.M.; Berdengalieva, A.N. Spatio-temporal dynamics of burnt areas in federal Protected Areas in the south-east of European Russia. Nat. Conserv. Res. 2021, 6, 23–44. [Google Scholar] [CrossRef]
- Pavleychik, V.M. Actual problems of investigation and monitoring of steppe fires (at the example of the zavolzhie and south Ural regions). Probl. Ecol. Monit. Ecosyst. Model. 2017, XXVIII, 86–99. [Google Scholar] [CrossRef]
- Freitag, M.; Kamp, J.; Dara, A.; Kuemmerle, T.; Sidorova, T.V.; Stirnemann, I.A.; Velbert, F.; Hölzel, N. Post-Soviet shifts in grazing and fire regimes changed the functional plant community composition on the Eurasian steppe. Glob. Chang. Biol. 2021, 27, 388–401. [Google Scholar] [CrossRef]
- Dara, A.; Baumann, M.; Hölzel, N.; Hostert, P.; Kamp, J.; Müller, D.; Ullrich, B.; Kuemmerle, T. Post-Soviet Land-Use Change Affected Fire Regimes on the Eurasian Steppes. Ecosystems 2020, 23, 943–956. [Google Scholar] [CrossRef]
- Levina, F.Y. To the biology of the renewal of wormwood and other xerophytic semi-shrubs. Bot. Zhurnal 1953, 6, 905–908. [Google Scholar]
- Bespalova, Z.P. To the biology of semishrubs-ediphicators of phytocenoesis of Nogaiskand dry steppes of the Central Kazakhstan. Bot. Zhurnal 1960, 45, 1462–1475. [Google Scholar]
- Smelyanskij, I.E.; Bujvolov, Y.A.; Bazhenov, Y.A.; Bakirova, R.T.; Borovik, L.P.; Borodin, A.P.; Bykova, E.P.; Vlasov, A.A.; Gavrilenko, V.S.; Goroshko, O.A.; et al. Steppe Fires and Fire Situation Management in Steppe Protected Areas: Ecological and Environmental Aspects. Analytical Review; Smelyanskij, I.E., Ed.; BCC Press: Moskva, Russia, 2015; ISBN 978-5-93699-076-2. [Google Scholar]
- Fedorova, N.L. Structure and Dynamics of Natural Ecosystems and Their Components in the State Natural Biosphere Reserve “Chernye Zemli”. Ph.D. Thesis, Kalmyk State University, Saratov, Russia, 2012. 20p. Available online: https://new-disser.ru/_avtoreferats/01005516680.pdf (accessed on 1 January 2024).
- Lazareva, V.G.; Bananova, V.A.; Van Zung, N. Dynamics of Modern Vegetation for Pasture Use in the Northwestern Pre-Caspian Region. Arid Ecosyst. 2020, 10, 276–283. [Google Scholar] [CrossRef]
- Miroshnichenko, Y.M. Steppes of Eurasia and new boundaries between steppes and deserts. Phytodiversity East. Eur. 2006, 1, 215–231. [Google Scholar]
- Badmaev, V.S.; Ubushaev, B.S. State of vegetation on the territory of the “Chernye Zemli” reserve. Arid Ecosyst. 2004, 10, 22–25. [Google Scholar]
- Vaskina, N.A.; Ayusheva, Y.C.; Khalginova, B.V.; Dzhapova, R.R. Grass plants associations in the zone of Black Soils in Kalmykia. Izv. Orenbg. State Agrar. Univ. 2013, 3, 35–37. [Google Scholar]
- Fedorova, N.L.; Ulanova, S.S. The current state of landscapes of natural geosystems of the Black Land Reserve (based on field research). In Proceedings of the Steppes of Northern Eurasia: Proceedings of the IX International Symposium; Chibilev, A.A., Ed.; Orenburg State University: Orenburg, Russia, 2021; pp. 824–829. [Google Scholar]
- Klink, G.V.; Semenkov, I.N.; Nukhimovskaya, Y.D.; Gasanova, Z.U.; Stepanova, N.Y.; Konyushkova, M.V. Temporal change in plant communities and its relationship to soil salinity and microtopography on the Caspian Sea coast. Sci. Rep. 2022, 12, 18082. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Agro-Climatic Resources of the Kalmyk ASSR; Narodetskaya, S.S.; Belousova, Z.N.; Dzhidzhikov, V.N. (Eds.) Gidrometeoizdat: Saint Petersburg, Russia, 1974. [Google Scholar]
- Chemidov, M.M. Dynamics of Natural Self-Recovery of Grass Vegetation on the Black Lands of the Republic of Kalmykia; Natural Institute of Astrakhan State University: Astrakhan, Russia, 2009; 23p. [Google Scholar]
- Ubushaev, B.S.; Skidanov, D.S.; Badmaev, V.S. Diversity of hayfiels and pastures in Kalmykia republic. Izv. Samara Sci. Cent. Russ. Acad. Sci. 2011, 13, 922–925. [Google Scholar]
- Dzhapova, R.R.; Gavinova, A.N.; Menkebairova, B.V. The role of vegetation of automorphic solonetzic soils in vegetable complexes of Caspian lowland in the territory of Kalmykia. Mod. Probl. Sci. Educ. 2016, 4, 24984. [Google Scholar]
- Safronova, I. “Chernye Zemli” Reserve in the System of Botanical and Geographical Demarcation. Polevye Issled. 2020, 7, 40–47. [Google Scholar] [CrossRef]
- Key, C.H.; Benson, N.C. Measuring and remote sensing of burn severity [poster abstract]. Jt. Fire Sci. Conf. Work. Proc. 2000, 284–285. Available online: www.researchgate.net/publication/241687936_Measuring_and_remote_sensing_of_burn_severity_the_CBI_and_NBR (accessed on 1 January 2024).
- Karpachevskiy, A.; Mikhailukova, P.; Sharapova, A.; Semenkov, I.; Koroleva, T. Using SAR-data and optical data to delineate burned territories in Kalmykia, Chernye Zemli reserve. IOP Conf. Ser. Earth Environ. Sci. 2022, 1061, 012013. [Google Scholar] [CrossRef]
- POWO Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 1 January 2024).
- Pickett, S.T.A. Space-for-Time Substitution as an Alternative to Long-Term Studies. In Long-Term Studies in Ecology; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 January 2024).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Baker, W.L. Fire and restoration of sagebrush ecosystems. Wildl. Soc. Bull. 2006, 34, 177–185. [Google Scholar] [CrossRef]
- Crist, M.R.; Belger, R.; Davies, K.W.; Davis, D.M.; Meldrum, J.R.; Shinneman, D.J.; Remington, T.E.; Welty, J.; Mayer, K.E. Trends, Impacts, and Cost of Catastrophic and Frequent Wildfires in the Sagebrush Biome. Rangel. Ecol. Manag. 2023, 89, 3–19. [Google Scholar] [CrossRef]
- Bukowski, B.E.; Baker, W.L. Historical fire regimes, reconstructed from land-survey data, led to complexity and fluctuation in sagebrush landscapes. Ecol. Appl. 2013, 23, 546–564. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, B.; Luna, B.; Pérez, B.; Urbieta, I.R.; Moreno, J.M. Burning season effects on the short-term post-fire vegetation dynamics of a Mediterranean heathland. Appl. Veg. Sci. 2014, 17, 86–96. [Google Scholar] [CrossRef]
- Hansson, M.; Fogelfors, H. Management of a semi-natural grassland; results from a 15-year-old experiment in southern Sweden. J. Veg. Sci. 2000, 11, 31–38. [Google Scholar] [CrossRef]
- Wahlman, H.; Milberg, P. Management of semi-natural grassland vegetation: Evaluation of a long-term experiment in southern Sweden. Ann. Bot. Fenn. 2002, 39, 159–166. [Google Scholar]
- Antonsen, H.; Olsson, P.A. Relative importance of burning, mowing and species translocation in the restoration of a former boreal hayfield: Responses of plant diversity and the microbial community. J. Appl. Ecol. 2005, 42, 337–347. [Google Scholar] [CrossRef]
- Lednev, S.; Semenkov, I.; Koroleva, T. Comparison on effect of mowing and spring burning on the occurrence of grassland plants in Western Siberia (preliminary results). E3S Web Conf. 2023, 463, 02017. [Google Scholar] [CrossRef]
- Semenkov, I.N.; Lednev, S.A.; Klink, G.V.; Kasymov, D.P.; Agaontsev, M.V.; Kostrova, S.N.; Koroleva, T.V. Influence of spring burns on the properties of humus horizon of chernozem in the southeast of Western Siberia. Eurasian Soil Sci. 2024, 57, 493–501. [Google Scholar] [CrossRef]
- Rybashlykova, L.P. Effect of conservation and grazing, on the structure of cenopopulations in the community of Festuca valesiaca Gaudin. Izv. Timirazevsk. Selʹskohozajstvennoj Akad. 2022, 1, 52–60. [Google Scholar] [CrossRef]
- Oba, G.; Vetaas, O.R.; Stenseth, N.C. Relationships between biomass and plant species richness in arid-zone grazing lands. J. Appl. Ecol. 2001, 38, 836–845. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Wang, L.; Chen, L.; Song, N.; Gu, J.; Xue, Y. A six-year grazing exclusion changed plant species diversity of a Stipa breviflora desert steppe community, northern China. PeerJ 2018, 6, e4359. [Google Scholar] [CrossRef] [PubMed]
- Bogun, S.A. Saiga population state in the Black Lands reserve: Problems and prospects for its conservation. In Scientific Papers of Khvalynsky National Park: Compilation of Scientific Papers; Amirit Publishers, Ltd., 11: Saratov-Khvalynsk, Russia, 2019; pp. 7–14. [Google Scholar]
- Laycock, W.A.; Bartos, D.L.; Klement, K.D. Species richness inside and outside long-term exclosures. In Seed and Soil Dynamics in Shrubland Ecosystems: Proceedings; Hild, A.L., Shaw, N.L., Meyer, S.E., Booth, D.T., McArthur, E.D., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2004; pp. 183–188. [Google Scholar]
- Ivanov, V.V. Steppes of Western Kazakhstan in Connection with the Dynamics of Their Cover; Akademiya Nauk USSR: Moskva, Russia; Saint Petersburg, Russia, 1958. [Google Scholar]
- Tashninova, L.N.; Buvaev, D.A. Chernye Zemli of Kalmykia: Complex researches and development GIS. Geol. Geogr. Glob. Energy 2012, 4, 150–165. [Google Scholar]
- Shriver, R.K.; Andrews, C.M.; Arkle, R.S.; Barnard, D.M.; Duniway, M.C.; Germino, M.J.; Pilliod, D.S.; Pyke, D.A.; Welty, J.L.; Bradford, J.B. Transient population dynamics impede restoration and may promote ecosystem transformation after disturbance. Ecol. Lett. 2019, 22, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Power, S.C.; Davies, G.M.; Wainwright, C.E.; Marsh, M.; Bakker, J.D. Restoration temporarily supports the resilience of sagebrush-steppe ecosystems subjected to repeated fires. J. Appl. Ecol. 2023, 60, 1607–1621. [Google Scholar] [CrossRef]
- Zonneveld, I.S. Vicinism and mass effect. J. Veg. Sci. 1995, 6, 441–444. [Google Scholar] [CrossRef]
- Ryabinina, N.O.; Kanishchev, S.N.; Shinkarenko, S.S. The current state and dynamics of geosystems in the south-east of the Russian plain (by the example of the natural parks in Volgograd region). South Russ. Ecol. Dev. 2018, 13, 116–127. [Google Scholar] [CrossRef]
- Dusaeva, G.H.; Kalmykova, O. Influence of fires on vegetation cover of steppes: Literature review. Byulleten’ Mosk. Obs. Ispyt. Prir. Otd. Biol. 2021, 126, 26–38. [Google Scholar]
- Ager, A.A.; Vaillant, N.M.; McMahan, A. Restoration of fire in managed forests: A model to prioritize landscapes and analyze tradeoffs. Ecosphere 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Gordijn, P.J.; O’Connor, T.G. Multidecadal effects of fire in a grassland biodiversity hotspot: Does pyrodiversity enhance plant diversity? Ecol. Appl. 2021, 31, e02391. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Scheiter, S.; Higgins, S.I.; Osborne, C.P.; Bradshaw, C.; Lunt, D.; Ripley, B.S.; Taylor, L.L.; Beerling, D.J. Fire and fire-adapted vegetation promoted C4 expansion in the late Miocene. New Phytol. 2012, 195, 653–666. [Google Scholar] [CrossRef]
- Bazilevich, N.; Titljanova, A.A. Biotic Turnover on Five Continents: Element Exchange Processes in Terrestrial Natural Ecosystems; Tishkov, A.A., Ed.; Nauka: Novosibirsk, Russia, 2008; ISBN 978-5-7692-0941-3. [Google Scholar]
- Ryabtsov, S.N. The Effect of Pyrogenic Load on the Vegetation Cover of the Steppe of the Southern Urals. Doctoral Thesis, Orenburg State Educational University, Orenburg, Russia, 2005. [Google Scholar]
- Buivolov, Y.A.; Bykova, E.P.; Gavrilenko, V.S.; Gribkov, A.V.; Bazhenov, Y.A.; Borodin, A.P.; Goroshko, O.A.; Kirilyuk, V.E.; Korsun, O.V.; Kreindlin, M.L.; et al. Analysis of Domestic and Foreign Experience in Fire Management in the Steppes and Related Ecosystems, in Particular, in the Conditions of Protected Areas; Institute of Global Climate and Ecology of Roshydromet and RAS: Moskva, Russia, 2012. [Google Scholar]
- Maksimova, V.F.; Neronov, V.V. Influence of anthropogenic factors on the dynamics of vegetation within the black lands of Kalmykia during the second half of the 20th century. Vestn. Mosk. Unviersiteta Seriya 5 Geogr. 2013, 2, 78–83. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Chronoseries Stage and Time since the Last Fire | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chronosequence | (sub)shrub | Xerophytic-Grass | ||||||||
| Stage | 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 |
| Years | 1 | 4–6 | 9–13 | 15–17 | 20–35 | >35 | 1 | 4–6 | 9–13 | 15–17 |
| Annuals | ||||||||||
| Adonis aestivalis L. | – | – | – | – | 0.1 | – | – | – | – | – |
| Alyssum desertorum Stapf | 0.5 | 0.5 | 0.5–1 | 0.1–0.5 | 0.5 | 0.1–0.5 | – | 0.1–0.5 | 0.5–30 | 0.1–0.5 |
| Androsace maxima L. | – | – | – | – | 0.5 | – | – | – | – | 0.1 |
| Bromus tectorum L. | – | 0.5 | 0.5 | 0.5 | – | 0.5 | – | 0.5–25 | 0.5–1 | 0.5 |
| Buglossoides arvensis (L.) I.M.Johnst. | – | – | – | – | – | – | – | 0.5 | – | 0.1–0.5 |
| Ceratocarpus arenarius L. | – | 0.5 | – | 0.1–0.5 | – | – | – | – | 0.1–1 | 0.1–0.5 |
| Chorispora tenella (Pall.) DC | – | – | – | 0.1–0.5 | – | – | – | – | – | – |
| Crepis sancta (L.) Bornm. | – | – | – | – | 0.1 | – | – | 0.1–0.5 | 0.1–3 | – |
| Delphinium consolida subsp. divaricatum (Ledeb.) A.Nyár. | – | – | – | – | – | – | – | 0.5 | – | – |
| Descurainia sophia L. | – | – | – | 0.5 | – | – | – | 0.5 | 0.1–0.5 | 0.1–0.5 |
| Eremopyrum orientale (L.) Jaub. and Spach | – | – | – | 0.5 | 0.5 | 0.5 | – | 0.5 | – | 0.1 |
| Eremopyrum triticeum (Gaertn.) Nevski | – | – | – | – | – | 1 | – | – | – | 0.5 |
| Erodium cicutarium (L.) L’Hér. | – | – | – | – | – | 0.5 | – | 0.1–0.5 | – | 0.1 |
| Filago arvensis L. | – | – | – | – | – | 0.1 | – | 0.5 | 0.5–1 | 0.1 |
| Holosteum umbellatum L. | – | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | – | 0.1–0.5 | 0.1–0.5 | 0.5–1 |
| Lappula patula (Lehm.) Menyh. | – | – | – | 0.5 | – | 0.5 | – | – | 0.5 | – |
| Lepidium perfoliatum L. | 0.5 | 0.5 | 0.1–0.5 | 0.5 | 0.5 | 0.5–5 | – | 0.1 | – | 0.1 |
| Medicago orthoceras (Kar. and Kir.) Trautv. | – | 0.5–1 | 0.1 | 0.1–0.5 | 0.1 | 0.5–1 | – | 0.1–0.5 | 0.1–10 | 0.5 |
| Meniocus linifolius (Stephan ex Willd.) DC. | – | 0.1 | – | 0.5 | – | – | – | 0.5 | 0.5 | – |
| Myosotis stricta Link ex Roem. and Schult. | – | – | – | – | – | 0.5 | – | 0.5 | 0.1–0.5 | 0.1–10 |
| Ranunculus falcatus L. | 0.5 | 0.5 | – | 0.5–1 | 0.5 | 0.1–0.5 | 0.1–0.5 | 0.5–1 | 0.1–0.5 | 0.5–1 |
| Ranunculus testiculatus Crantz | 0.5 | – | – | – | – | 0.1–0.5 | – | 0.5 | – | – |
| Salsola tragus L. | – | – | – | 0.5 | – | – | – | – | 0.5 | 0.5 |
| Senecio vernalis Waldst. and Kit. | – | – | – | – | – | – | – | – | 0.5 | – |
| Veronica triphyllos L. | – | – | – | 0.5 | – | 0.5 | 0.5 | 0.5–2 | 0.5 | 0.1–0.5 |
| Veronica verna L. | – | 0.5 | 0.5 | – | 0.5–1 | 0.5–1 | – | 0.1–0.5 | – | – |
| Annuals or biennials | ||||||||||
| Sisymbrium altissimum L. | – | – | – | – | – | – | – | – | 0.5 | – |
| Sisymbrium loeselii L. | – | – | – | – | – | 0.5 | – | – | 0.5 | – |
| Biennials | ||||||||||
| Falcaria vulgaris Bernh. | – | – | – | – | – | 0.5 | – | – | – | – |
| Perennials | ||||||||||
| Agropyron fragile (Roth) P.Candargy | – | – | – | 0.5–3 | – | 1 | – | 0.5 | 0.1 | – |
| Anabasis aphylla L. | – | 0.5 | – | 0.5 | – | 0.1–0.5 | – | – | – | 0.5–5 |
| Artemisia lercheana Weber ex Stechm. | 0.1 | – | – | 2–40 | 2–5 | 1–20 | – | 0.5 | – | – |
| Artemisia pauciflora Weber ex Stechmann | – | – | – | – | 0.5 | 1–3 | – | – | – | – |
| Artemisia santonicum L. | – | 1–5 | 1–5 | – | 5 | – | – | – | – | – |
| Astragalus dolichophyllus Pall. | – | 0.5 | – | 0.1–0.5 | 0.1–1 | – | – | – | – | – |
| Astragalus longipetalus Chater | – | – | – | 0.1–0.5 | – | – | – | – | – | – |
| Astragalus varius S.G.Gmel. | – | – | – | 0.5 | – | 0.5 | – | – | – | – |
| Bassia prostrata (L.) Beck | – | – | 0.1 | 1 | – | 0.5 | – | – | – | – |
| Carduus uncinatus M.Bieb. | – | – | – | – | – | – | – | 0.5 | 0.5–1 | – |
| Carex stenophylla Wahlenb. | 0.5–1 | 0.5–1 | 0.5–3 | 0.5 | 10–15 | 0.5–1 | 3–5 | 1–10 | 0.5–1 | 0.5–3 |
| Convolvulus arvensis L. | – | – | – | 0.5 | – | – | – | – | – | – |
| Elymus repens (L.) Gould | 0.5 | 0.5 | 0.5 | – | 0.1–0.5 | 0.5 | – | 0.5 | 1 | 0.5 |
| Gypsophila paniculata L. | – | – | – | – | – | – | – | 0.5 | 0.1 | – |
| Krascheninnikovia ceratoides (L.) Gueldenst. | – | – | – | – | – | – | – | – | – | 0.5 |
| Peganum harmala L. | – | – | – | 0.1 | – | – | – | – | – | – |
| Phlomis herba-venti L. | – | – | – | – | – | 0.5 | – | 0.1 | – | – |
| Poa bulbosa L. | 5–10 | 10–20 | 20–35 | 15–35 | 20–45 | 3–40 | 1–10 | 0.1–20 | 1–50 | 1–50 |
| Prangos odontalgica (Pall.) Herrnst. and Heyn | 0.5 | 0.5 | 0.1–0.5 | 0.5 | 0.5 | 0.1–1 | 0.1–0.5 | 0.5 | 0.1 | 0.1–0.5 |
| Ranunculus oxyspermus Willd. | 0.5–1 | 0.5 | 0.5–1 | 0.5–0.1 | 0.1–1 | 0.1–0.5 | 0.5–2 | 0.5–2 | 0.5–1 | 0.1–3 |
| Stipa lessingiana Trin. and Rupr. | – | – | – | – | – | – | – | – | 0.5–5 | – |
| Stipa sareptana A.K.Becker | – | 0.5–1 | 0.1–1 | 1 | 0.5 | 0.5 | 0.5–1 | 0.5–10 | 0.1–10 | 1–10 |
| Tanacetum achilleifolium (M.Bieb.) Sch.Bip. | 0.5–1 | – | 0.5–1 | 0.5 | 0.5–1 | 0.5 | – | – | – | – |
| Tulipa biflora Pall. | 0.1–0.5 | 0.5 | – | – | – | 0.5 | – | – | – | – |
| Tulipa sylvestris subsp. australis (Link) Pamp. | – | 0.5 | – | 0.5 | – | – | 0.5–2 | – | – | – |
| n | 4 | 3 | 4 | 7 | 4 | 7 | 4 | 10 | 7 | 5 |
| Projective Cover of Artemisia spp., % | Number of 10 × 10 Plots | Mean Number of Species on 10 × 10 m Plot | |||
|---|---|---|---|---|---|
| In Reserve | Outside Reserve | In Reserve | Outside Reserve | p-Value of Mann–Whitney Test | |
| <1 | 24 | 9 | 11.4 | 11.9 | 0.8 |
| [1–10) | 6 | 7 | 17.3 | 14.0 | 0.2 |
| Species | Species Representation * | p Value of Significance of Differences ** | Median Projective Cover, % | |||
|---|---|---|---|---|---|---|
| In the Reserve | Outside the Reserve | In the Reserve | Outside the Reserve | |||
| Communities with Sagebrush Cover <1% | ||||||
| Alyssum desertorum Stapf | 3.55 | 0.46 | 0.34 | 0.3 | 0.5 | |
| Bromus tectorum L. | 2.05 | 0.11 | 0.29 | 0 | 0 | |
| Carduus uncinatus M.Bieb. | 0.15 | 0.06 | 0.51 | 0 | 0 | |
| Carex stenophylla Wahlenb. | 2.38 | 1.61 | 0.9 | 0.5 | 0.5 | |
| Ceratocarpus arenarius L. | 0.13 | 0.17 | 0.78 | 0 | 0 | |
| Crepis sancta (L.) Bornm. | 0.28 | 0.11 | 0.35 | 0 | 0 | |
| Descurainia sophia (L.) Webb ex Prantl | 0.21 | 0.12 | 0.43 | 0 | 0 | |
| Erodium cicutarium (L.) L’Hér. | 0.03 | 0.01 | 0.87 | 0 | 0 | |
| Filago arvensis L. | 0.17 | 0.01 | 0.22 | 0 | 0 | |
| Gypsophila paniculata L. | 0 | 0.11 | 0.11 | 0 | 0 | |
| Holosteum umbellatum L. | 0.26 | 0.11 | 0.14 | 0.1 | 0 | |
| Medicago orthoceras (Kar. & Kir.) Trautv. | 1.15 | 0.17 | 0.18 | 0.3 | 0 | |
| Myosotis stricta Link ex Roem. and Schult. | 0.46 | 0.01 | 0.65 | 0 | 0 | |
| Poa bulbosa L. | 10.9 | 18.67 | 0.031 | 4 | 10 | |
| Polygonum sp. | 0.1 | 0.01 | 0.45 | 0 | 0 | |
| Prangos odontalgica (Pall.) Herrnst. and Heyn | 0.05 | 0.41 | <0.0001 | 0 | 0.5 | |
| Ranunculus falcatus L. | 0.32 | 0.39 | 0.47 | 0.5 | 0.5 | |
| Ranunculus oxyspermus Willd. | 0.73 | 0.5 | 0.93 | 0.5 | 0.5 | |
| Ranunculus testiculatus Crantz | 0.04 | 0.11 | 0.3 | 0 | 0 | |
| Salsola tragus L. | 0.08 | 0.06 | 0.72 | 0 | 0 | |
| Stipa sareptana A.K.Becker | 3.65 | 0.12 | 0.0009 | 1 | 0 | |
| Veronica triphyllos L. | 0.46 | 0.12 | 0.0052 | 0.5 | 0 | |
| Communities with sagebrush cover 1–10% | ||||||
| Achillea leptophylla M.Bieb. | 0.04 | 0.08 | 0.67 | 0 | 0 | |
| Agropyron fragile (Roth) P.Candargy | 2.75 | 0.08 | 0.22 | 0 | 0 | |
| Alyssum desertorum Stapf | 0.72 | 5.33 | 0.92 | 0.5 | 0.5 | |
| Anabasis aphylla L. | 0.04 | 0.17 | 0.22 | 0 | 0 | |
| Artemisia arenaria DC | 0.29 | 0.33 | 1 | 0 | 0 | |
| Artemisia lercheana Weber ex Stechm. | 2.17 | 1.17 | 0.36 | 1.5 | 0 | |
| Artemisia santonicum L. | 0.75 | 2 | 0.26 | 0 | 1 | |
| Astragalus dolichophyllus Pall. | 0.13 | 0.02 | 0.65 | 0 | 0 | |
| Astragalus varius S.G.Gmel. | 0.12 | 0.02 | 0.61 | 0 | 0 | |
| Bassia prostrata (L.) Beck | 0.12 | 0.1 | 0.61 | 0 | 0 | |
| Bromus tectorum L. | 0.04 | 0.27 | 0.016 | 0 | 0.3 | |
| Carduus uncinatus M.Bieb. | 0.08 | 0.08 | 1 | 0 | 0 | |
| Carex stenophylla Wahlenb. | 3.38 | 0.67 | 0.66 | 0.75 | 0.5 | |
| Ceratocarpus arenarius L. | 0.01 | 0.02 | 0.67 | 0 | 0 | |
| Crepis sancta (L.) Bornm. | 0.09 | 0.08 | 0.8 | 0 | 0 | |
| Descurainia sophia (L.) Webb ex Prantl | 0.04 | 0.08 | 0.67 | 0 | 0 | |
| Elymus repens (L.) Gould | 0.13 | 0.08 | 0.55 | 0 | 0 | |
| Eremopyrum orientale (L.) Jaub. and Spach | 0.08 | 0.02 | 0.94 | 0 | 0 | |
| Gypsophila paniculata L. | 0.04 | 0.08 | 0.67 | 0 | 0 | |
| Holosteum umbellatum L. | 0.33 | 0.17 | 0.21 | 0.5 | 0 | |
| Lepidium perfoliatum L. | 0.3 | 0.33 | 0.87 | 0.5 | 0.5 | |
| Medicago orthoceras (Kar. and Kir.) Trautv. | 0.35 | 0.37 | 0.66 | 0.1 | 0.3 | |
| Meniocus linifolius (Stephan ex Willd.) DC. | 0.08 | 0.1 | 0.56 | 0 | 0 | |
| Poa bulbosa L. | 21 | 35 | 0.015 | 20 | 35 | |
| Polygonum sp. | 0.09 | 0.12 | 0.43 | 0 | 0.05 | |
| Prangos odontalgica (Pall.) Herrnst. and Heyn | 0.46 | 0.27 | 0.18 | 0.5 | 0.3 | |
| Ranunculus oxyspermus Willd. | 0.34 | 0.42 | 0.52 | 0.05 | 0.5 | |
| Ranunculus testiculatus Crantz | 0.13 | 0.17 | 0.95 | 0 | 0 | |
| Salsola tragus L. | 0.04 | 0.02 | 0.73 | 0 | 0 | |
| Sisymbrium altissimum L. | 0.04 | 0.08 | 0.67 | 0 | 0 | |
| Sisymbrium loeselii L. | 0.12 | 0.08 | 0.75 | 0 | 0 | |
| Stipa sareptana A.K.Becker | 0.13 | 0.5 | 0.086 | 0 | 0.5 | |
| Tanacetum achilleifolium (M.Bieb.) Sch.Bip. | 0.25 | 0.17 | 0.87 | 0 | 0 | |
| Tulipa sylvestris subsp. australis (Link) Pamp. | 0.04 | 0.08 | 0.67 | 0 | 0 | |
| Veronica triphyllos L. | 0.21 | 0.18 | 1 | 0 | 0.05 | |
| Veronica verna L. | 0.33 | 0.25 | 0.8 | 0.25 | 0.25 | |
| Stage | p | r | N Species at the Stage | |||
|---|---|---|---|---|---|---|
| I | II | In Total | I | II | ||
| 1 | 2 | 0.0031 | 0.7289 | 14 | 15 | 36 |
| 1 | 3 | 0.1403 | 0.4745 | 11 | 15 | 33 |
| 1 | 4 | 0.5905 | 0.1648 | 13 | 15 | 39 |
| 1 | 5 | 0.1365 | 0.4786 | 11 | 15 | 22 |
| 1 | 6 | 0.3724 | 0.2584 | 14 | 15 | 33 |
| 2 | 3 | 0.0000 | 0.7773 | 24 | 36 | 33 |
| 2 | 4 | 0.1621 | 0.2824 | 26 | 36 | 39 |
| 2 | 5 | 0.2700 | 0.2836 | 17 | 36 | 22 |
| 2 | 6 | 0.0738 | 0.3716 | 24 | 36 | 33 |
| 3 | 4 | 0.0522 | 0.4008 | 24 | 33 | 39 |
| 3 | 5 | 0.5561 | 0.1591 | 16 | 33 | 22 |
| 3 | 6 | 0.0656 | 0.3993 | 22 | 33 | 33 |
| 4 | 5 | 0.0153 | 0.5771 | 17 | 39 | 22 |
| 4 | 6 | 0.0088 | 0.5033 | 26 | 39 | 33 |
| 5 | 6 | 0.0460 | 0.4898 | 17 | 22 | 33 |
| Chronosequence | Number of Species | Species Projective Cover | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Mean | Median | ||||||
| − | + | − | + | − | + | ||
| (Sub)shrub | 21 | 12 | 0.18 | 2.47 | 0.14 | 0.23 | 0.02 |
| Xerophytic-grass | 23 | 7 | 0.3 | 2.8 | 0.05 | 0.6 | 0.0003 |
| Plant Species | Projective Cover in the Group, % | p-Value | ||||
|---|---|---|---|---|---|---|
| Mean | Median | Without Bonferroni Correction | With Bonferroni Correction | |||
| I | II | I | II | |||
| Achillea leptophylla M.Bieb. | 0.07 | 0.03 | 0 | 0 | 0.44 | 1 |
| Agropyron fragile (Roth) P.Candargy | 1.3 | 2.1 | 0 | 0 | 0.8 | 1 |
| Alyssum desertorum Stapf | 2.9 | 4.4 | 0.5 | 0.5 | 0.61 | 1 |
| Amaranthus sp. | 0 | 0.03 | 0 | 0 | 0.41 | 1 |
| Anabasis aphylla L. | 0.4 | 0.03 | 0 | 0 | 0.19 | 1 |
| Androsace maxima L. | 0.01 | 0 | 0 | 0 | 0.29 | 1 |
| Artemisia arenaria DC. | 0.4 | 0.1 | 0 | 0 | 0.66 | 1 |
| Artemisia austriaca Jacq. | 0 | 0.05 | 0 | 0 | 0.41 | 1 |
| Artemisia lercheana Weber ex Stechm. | 5.5 | 0 | 0 | 0 | 0 | 0.2 |
| Artemisia santonica Lam. | 0.9 | 0.5 | 0 | 0 | 0.45 | 1 |
| Astragalus dolichophyllus Pall. | 0.04 | 0.03 | 0 | 0 | 0.46 | 1 |
| Astragalus longipetalus Chater | 0.03 | 0.06 | 0 | 0 | 0.46 | 1 |
| Astragalus varius S.G.Gmel. | 0.01 | 0.03 | 0 | 0 | 0.48 | 1 |
| Bassia prostrata (L.) Beck | 0.11 | 0 | 0 | 0 | 0.05 | 1 |
| Bromus tectorum L. | 0.2 | 2.6 | 0 | 0 | 0.46 | 1 |
| Buglossoides arvensis (L.) I.M.Johnst. | 0 | 0.05 | 0 | 0 | 0.22 | 1 |
| Carduus uncinatus M.Bieb. | 0.1 | 0.2 | 0 | 0 | 0.39 | 1 |
| Carex stenophylla Wahlenb. | 0.6 | 2.1 | 0 | 0.5 | 0.36 | 1 |
| Centaurea diffusa Lam. | 0.03 | 0 | 0 | 0 | 0.29 | 1 |
| Ceratocarpus arenarius L. | 0.11 | 0.14 | 0 | 0 | 0.96 | 1 |
| Chorispora tenella (Pall.) DC | 0.08 | 0 | 0 | 0 | 0.02 | 1 |
| Convolvulus arvensis L. | 0.03 | 0.05 | 0 | 0 | 0.72 | 1 |
| Crepis sancta (L.) Bornm. | 0.1 | 0.4 | 0 | 0.5 | 0.02 | 1 |
| Delphinium consolida subsp. divaricatum (Ledeb.) A.Nyár. | 0 | 0.03 | 0 | 0 | 0.41 | 1 |
| Descurainia sophia L. | 0.14 | 0.24 | 0 | 0.1 | 0.24 | 1 |
| Elymus repens (L.) Gould | 0.13 | 0.13 | 0 | 0 | 0.8 | 1 |
| Eremopyrum orientale (L.) Jaub. and Spach | 0.03 | 0 | 0 | 0 | 0.29 | 1 |
| Eremopyrum triticeum (Gaertn.) Nevski | 0.07 | 0 | 0 | 0 | 0.12 | 1 |
| Erodium cicutarium (L.) L’Hér. | 0 | 0.03 | 0 | 0 | 0.22 | 1 |
| Erodium hoefftianum C.A.Mey. | 0.03 | 0 | 0 | 0 | 0.29 | 1 |
| Filago arvensis L. | 0.01 | 0.21 | 0 | 0 | 0.03 | 1 |
| Gypsophila paniculata L. | 0.07 | 0.06 | 0 | 0 | 0.91 | 1 |
| Holosteum umbellatum L. | 0.2 | 0.27 | 0 | 0.5 | 0.3 | 1 |
| Lappula patula (Lehm.) Menyh. | 0.03 | 0.03 | 0 | 0 | 0.9 | 1 |
| Lepidium perfoliatum L. | 0.22 | 0.1 | 0.1 | 0 | 0.02 | 1 |
| Leymus racemosus (Lam.) Tzvelev | 0 | 0.05 | 0 | 0 | 0.22 | 1 |
| Linaria sp. | 0.03 | 0 | 0 | 0 | 0.29 | 1 |
| Medicago orthoceras (Kar. and Kir.) Trautv. | 0.3 | 1.6 | 0 | 0.5 | 0.02 | 1 |
| Meniocus linifolius (Stephan ex Willd.) DC. | 0.1 | 0.1 | 0 | 0 | 1 | 1 |
| Myosotis stricta Link ex Roem. and Schult. | 0.01 | 0.06 | 0 | 0 | 0.4 | 1 |
| Nonea caspica (Willd.) G.Don | 0 | 0.08 | 0 | 0 | 0.12 | 1 |
| Phlomis herba-venti L. | 0 | 0.01 | 0 | 0 | 0.41 | 1 |
| Poa bulbosa L. | 28 | 13 | 30 | 5 | 0 | 0.13 |
| Polygonum sp. | 0.15 | 0.11 | 0 | 0 | 0.21 | 1 |
| Prangos odontalgica (Pall.) Herrnst. and Heyn | 0.25 | 0.14 | 0.1 | 0 | 0.18 | 1 |
| Ranunculus falcatus L. | 0.23 | 0.29 | 0 | 0.5 | 0.44 | 1 |
| Ranunculus oxyspermus Willd. | 0.31 | 0.42 | 0.5 | 0.5 | 0.63 | 1 |
| Ranunculus testiculatus Crantz | 0 | 0.08 | 0 | 0 | 0.12 | 1 |
| Salsola tragus L. | 0.04 | 0.05 | 0 | 0 | 0.88 | 1 |
| Senecio sp. | 0 | 0.1 | 0 | 0 | 0.07 | 1 |
| Sisymbrium altissimum L. | 0.1 | 0.13 | 0 | 0 | 0.69 | 1 |
| Sisymbrium loeselii L. | 0.1 | 1.2 | 0 | 0 | 0.32 | 1 |
| Stipa lessingiana Trin. and Rupr. | 0 | 0.32 | 0 | 0 | 0.12 | 1 |
| Stipa sareptana A.K.Becker | 0.2 | 3 | 0 | 0.5 | 0 | 0.23 |
| Tanacetum achilleifolium med | 0.2 | 0 | 0 | 0 | 0.02 | 1 |
| Tulipa biflora Pall. | 0 | 0.03 | 0 | 0 | 0.41 | 1 |
| Tulipa sylvestris subsp. australis (Link) Pamp. | 0.03 | 0.08 | 0 | 0 | 0.44 | 1 |
| Veronica triphyllos L. | 0.2 | 0.5 | 0 | 0.5 | 0.03 | 1 |
| Veronica verna L. | 0.14 | 0.08 | 0 | 0 | 0.43 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klink, G.V.; Lednev, S.A.; Semenkov, I.N.; Konyushkova, M.V.; Karpachevskiy, A.M.; Chemidov, M.M.; Ulanova, S.S.; Fedorova, N.L.; Sharapova, A.V.; Bogun, S.A.; et al. Influence of Fires on Desert Plant Communities at the Chernye Zemli (SW Russia). Fire 2024, 7, 96. https://doi.org/10.3390/fire7030096
Klink GV, Lednev SA, Semenkov IN, Konyushkova MV, Karpachevskiy AM, Chemidov MM, Ulanova SS, Fedorova NL, Sharapova AV, Bogun SA, et al. Influence of Fires on Desert Plant Communities at the Chernye Zemli (SW Russia). Fire. 2024; 7(3):96. https://doi.org/10.3390/fire7030096
Chicago/Turabian StyleKlink, Galya V., Sergey A. Lednev, Ivan N. Semenkov, Maria V. Konyushkova, Andrey M. Karpachevskiy, Mergen M. Chemidov, Svetlana S. Ulanova, Natal’ya L. Fedorova, Anna V. Sharapova, Sergey A. Bogun, and et al. 2024. "Influence of Fires on Desert Plant Communities at the Chernye Zemli (SW Russia)" Fire 7, no. 3: 96. https://doi.org/10.3390/fire7030096
APA StyleKlink, G. V., Lednev, S. A., Semenkov, I. N., Konyushkova, M. V., Karpachevskiy, A. M., Chemidov, M. M., Ulanova, S. S., Fedorova, N. L., Sharapova, A. V., Bogun, S. A., & Koroleva, T. V. (2024). Influence of Fires on Desert Plant Communities at the Chernye Zemli (SW Russia). Fire, 7(3), 96. https://doi.org/10.3390/fire7030096