
Dissecting the Inorganic Nanoparticle-Driven Interferences on Adhesome Dynamics

Abstract
:1. Introduction
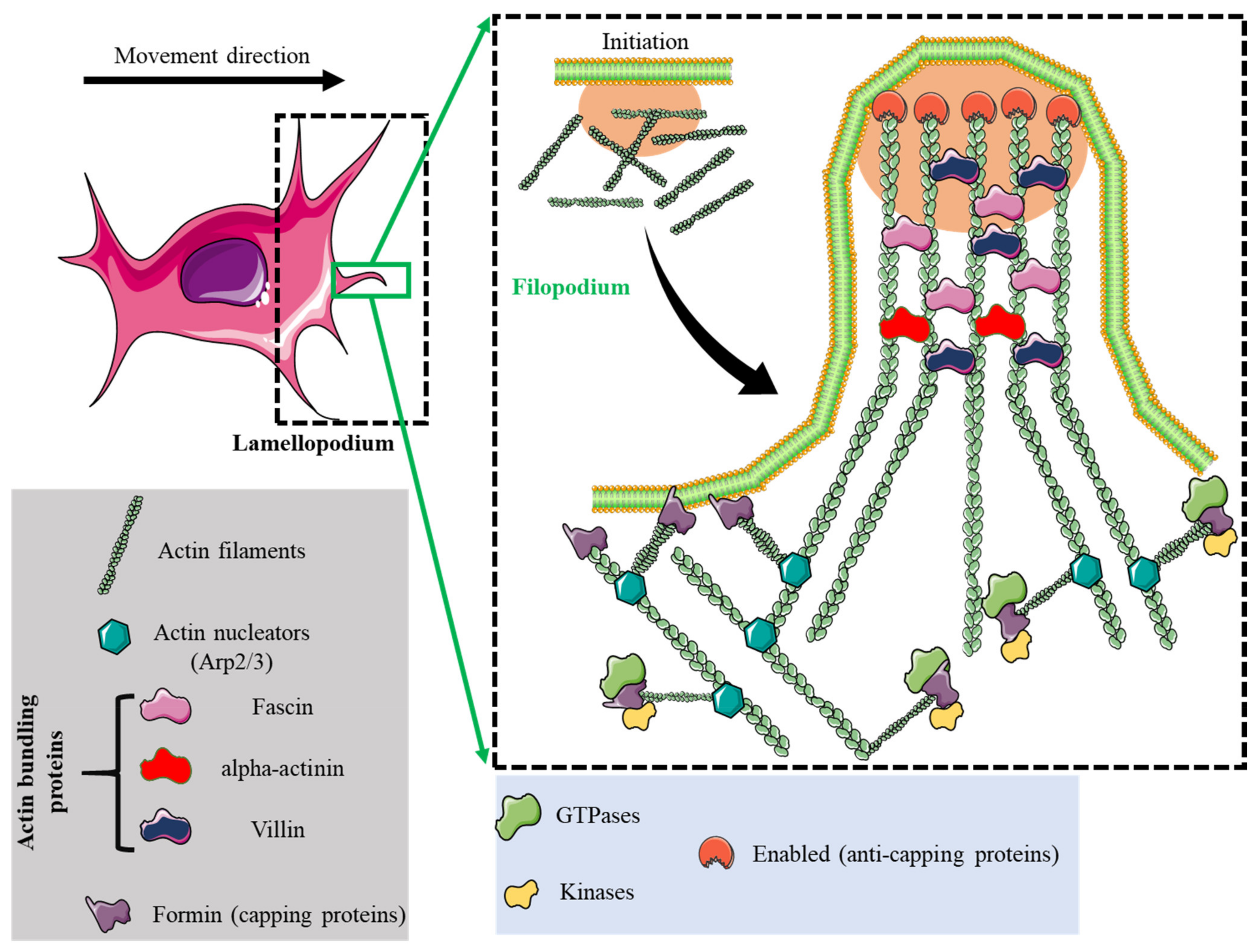
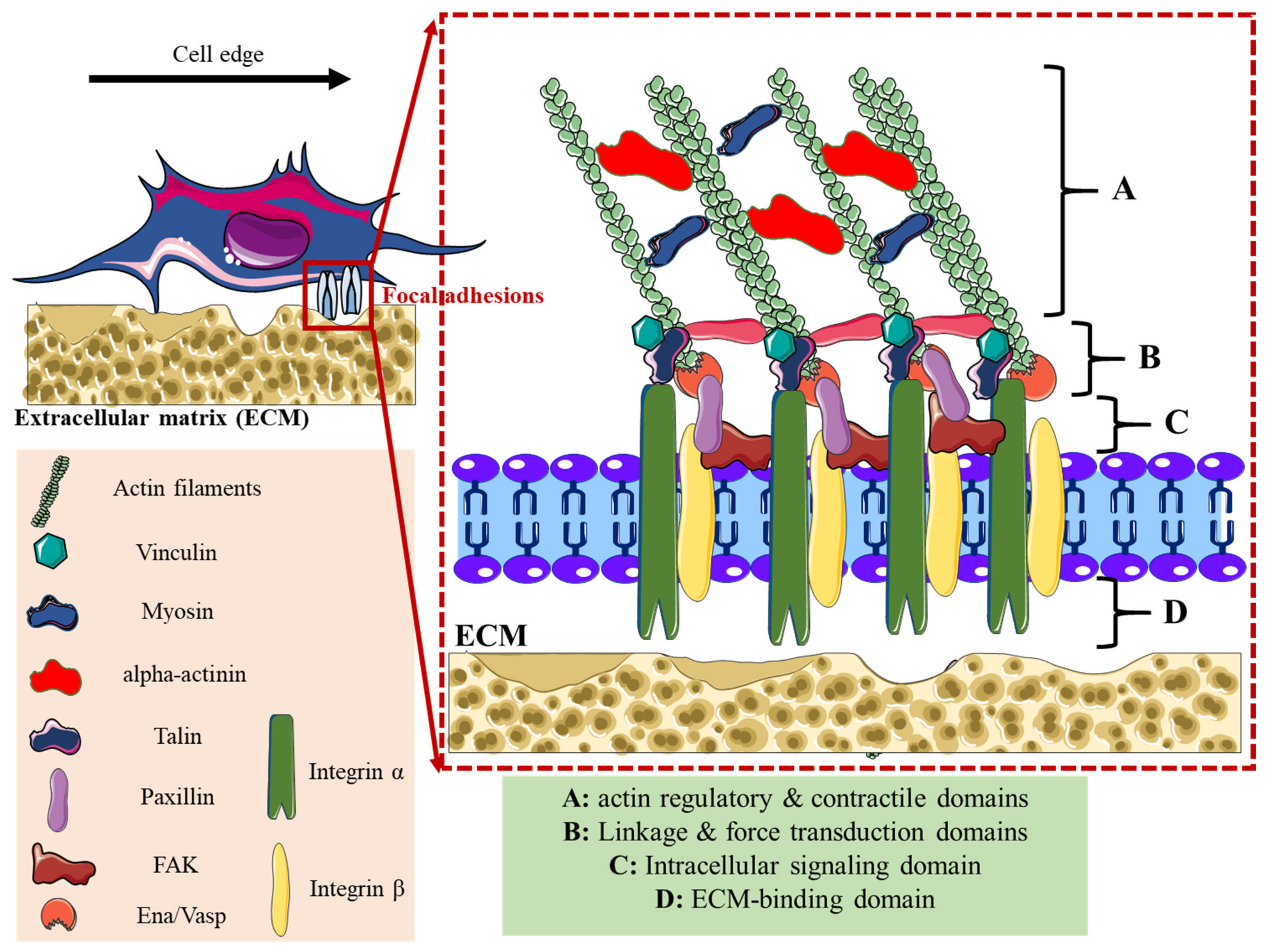
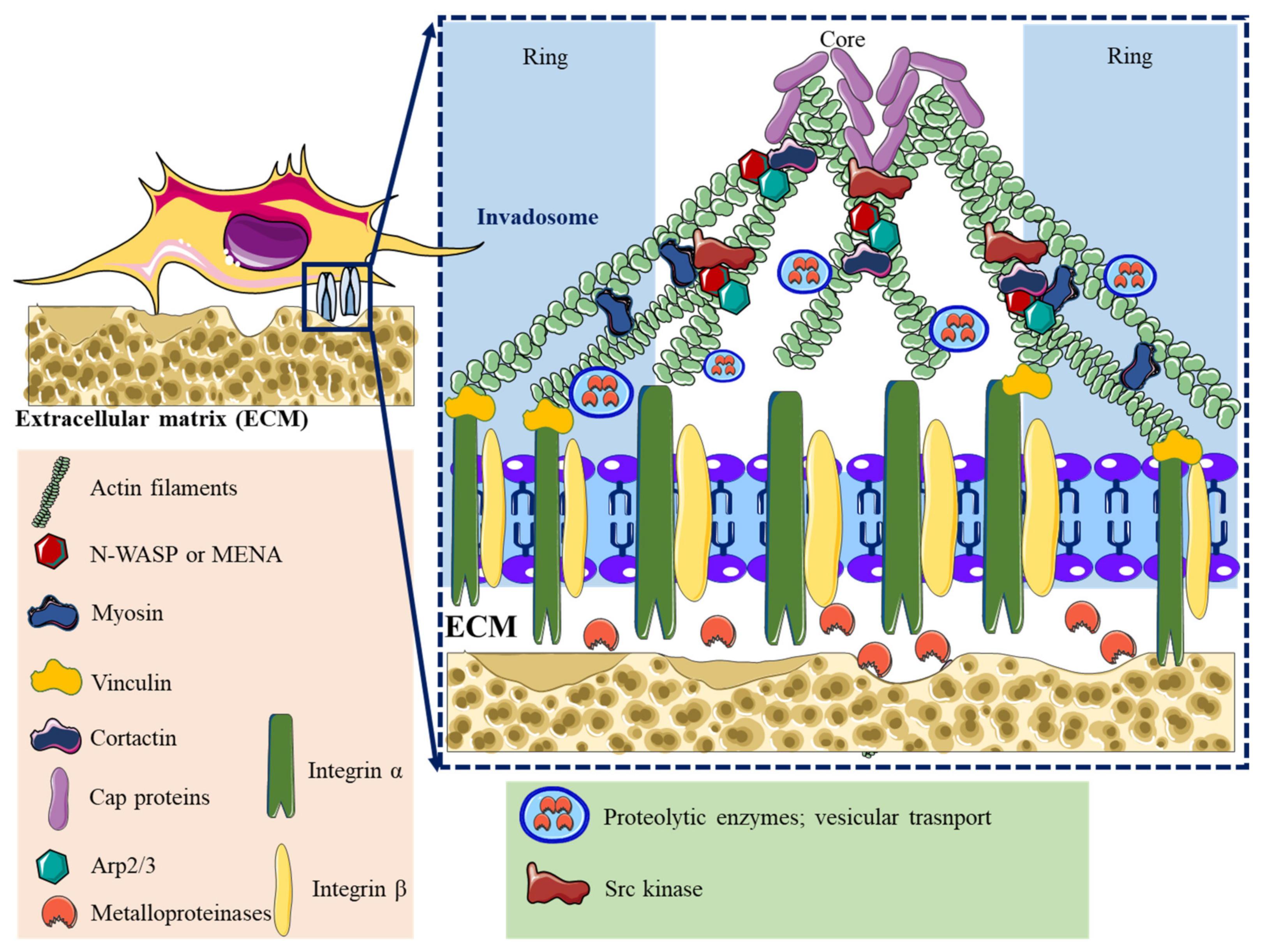
2. Adhesome Structure, Function, and Dynamics
3. Impact of Inorganic Nanoparticles on Adhesome Dynamics
3.1. Inorganic Nanoparticles and Focal Adhesions Dynamic
3.2. Nanopaterneting as a Platform for FA Study
3.3. Inorganic Nanoparticles and Invadosome Dynamic
4. Perspectives
Funding
Conflicts of Interest
References
- Habibi, N.; Quevedo, D.F.; Gregory, J.V.; Lahann, J. Emerging Methods in Therapeutics Using Multifunctional Nanoparticles. WIREs Nanomed. Nanobiotechnol. 2020, 12, e1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.U.; Khan, M.; Cho, M.H.; Khan, M.M. Selected Nanotechnologies and Nanostructures for Drug Delivery, Nanomedicine and Cure. Bioprocess Biosyst. Eng. 2020, 43, 1339–1357. [Google Scholar] [CrossRef]
- Wicki, A.; Witzigmann, D.; Balasubramanian, V.; Huwyler, J. Nanomedicine in Cancer Therapy: Challenges, Opportunities, and Clinical Applications. J. Control. Release 2015, 200, 138–157. [Google Scholar] [CrossRef]
- Hare, J.I.; Lammers, T.; Ashford, M.B.; Puri, S.; Storm, G.; Barry, S.T. Challenges and Strategies in Anti-Cancer Nanomedicine Development: An Industry Perspective. Adv. Drug Deliv. Rev. 2017, 108, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Gharagozloo, M.; Majewski, S.; Foldvari, M. Therapeutic Applications of Nanomedicine in Autoimmune Diseases: From Immunosuppression to Tolerance Induction. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1003–1018. [Google Scholar] [CrossRef]
- Qamar, N.; Arif, A.; Bhatti, A.; John, P. Nanomedicine: An Emerging Era of Theranostics and Therapeutics for Rheumatoid Arthritis. Rheumatology 2019, 58, 1715–1721. [Google Scholar] [CrossRef] [PubMed]
- Poovaiah, N.; Davoudi, Z.; Peng, H.; Schlichtmann, B.; Mallapragada, S.; Narasimhan, B.; Wang, Q. Treatment of Neurodegenerative Disorders through the Blood–Brain Barrier Using Nanocarriers. Nanoscale 2018, 10, 16962–16983. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, S.; Archunan, G.; Sivakumar, M.; Selvan, S.T.; Fred, A.L.; Kumar, S.; Gulyás, B.; Padmanabhan, P. Theranostic Applications of Nanoparticles in Neurodegenerative Disorders. Int. J. Nanomed. 2018, 13, 5561–5576. [Google Scholar] [CrossRef] [Green Version]
- Steichen, S.D.; Caldorera-Moore, M.; Peppas, N.A. A Review of Current Nanoparticle and Targeting Moieties for the Delivery of Cancer Therapeutics. Eur. J. Pharm. Sci. 2013, 48, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amreddy, N.; Babu, A.; Muralidharan, R.; Panneerselvam, J.; Srivastava, A.; Ahmed, R.; Mehta, M.; Munshi, A.; Ramesh, R. Chapter Five—Recent Advances in Nanoparticle-Based Cancer Drug and Gene Delivery. In Advances in Cancer Research; Tew, K.D., Fisher, P.B., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 137, pp. 115–170. ISBN 0065-230X. [Google Scholar]
- Deirram, N.; Zhang, C.; Kermaniyan, S.S.; Johnston, A.P.R.; Such, G.K. PH-Responsive Polymer Nanoparticles for Drug Delivery. Macromol. Rapid Commun. 2019, 40, 1800917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, M.; Eslami, M.; Sahandi-Zangabad, P.; Mirab, F.; Farajisafiloo, N.; Shafaei, Z.; Ghosh, D.; Bozorgomid, M.; Dashkhaneh, F.; Hamblin, M.R. PH-Sensitive Stimulus-Responsive Nanocarriers for Targeted Delivery of Therapeutic Agents. WIREs Nanomed. Nanobiotechnol. 2016, 8, 696–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Saw, P.E.; Tao, W.; Li, Y.; Ji, X.; Bhasin, S.; Liu, Y.; Ayyash, D.; Rasmussen, J.; Huo, M.; et al. ROS-Responsive Polyprodrug Nanoparticles for Triggered Drug Delivery and Effective Cancer Therapy. Adv. Mater. 2017, 29, 1700141. [Google Scholar] [CrossRef]
- Lv, X.; Zhu, Y.; Ghandehari, H.; Yu, A.; Wang, Y. An ROS-Responsive and Self-Accelerating Drug Release Nanoplatform for Overcoming Multidrug Resistance. Chem. Commun. 2019, 55, 3383–3386. [Google Scholar] [CrossRef] [PubMed]
- Halappanavar, S.; Vogel, U.; Wallin, H.; Yauk, C.L. Promise and Peril in Nanomedicine: The Challenges and Needs for Integrated Systems Biology Approaches to Define Health Risk. WIREs Nanomed. Nanobiotechnol. 2018, 10, e1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelaz, B.; Charron, G.; Pfeiffer, C.; Zhao, Y.; de la Fuente, J.M.; Liang, X.-J.; Parak, W.J.; Del Pino, P. Interfacing Engineered Nanoparticles with Biological Systems: Anticipating Adverse Nano-Bio Interactions. Small 2013, 9, 1573–1584. [Google Scholar] [CrossRef]
- Su, H.; Wang, Y.; Gu, Y.; Bowman, L.; Zhao, J.; Ding, M. Potential Applications and Human Biosafety of Nanomaterials Used in Nanomedicine. J. Appl. Toxicol. 2018, 38, 3–24. [Google Scholar] [CrossRef]
- Tang, D.D.; Gerlach, B.D. The Roles and Regulation of the Actin Cytoskeleton, Intermediate Filaments and Microtubules in Smooth Muscle Cell Migration. Respir. Res. 2017, 18, 54. [Google Scholar] [CrossRef] [Green Version]
- Fife, C.M.; McCarroll, J.A.; Kavallaris, M. Movers and Shakers: Cell Cytoskeleton in Cancer Metastasis. Br. J. Pharmacol. 2014, 171, 5507–5523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burbage, M.; Keppler, S.J. Shaping the Humoral Immune Response: Actin Regulators Modulate Antigen Presentation and Influence B-T Interactions. Mol. Immunol. 2018, 101, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Shenoy, A.R. The Cytoskeleton in Cell-Autonomous Immunity: Structural Determinants of Host Defence. Nat. Rev. Immunol. 2015, 15, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Kasahara, K.; Inagaki, M. Intermediate Filaments and IF-Associated Proteins: From Cell Architecture to Cell Proliferation. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 479–493. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, A.; Szczepanski, M.A.; Klimaszewska-Wisniewska, A.; Gackowska, L.; Zuryn, A.; Grzanka, A. Cytoskeletal Reorganization and Cell Death in Mitoxantrone-Treated Lung Cancer Cells. Acta Histochem. 2016, 118, 784–796. [Google Scholar] [CrossRef]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma’ayan, A.; Iyengar, R.; Geiger, B. Functional Atlas of the Integrin Adhesome. Nat. Cell Biol. 2007, 9, 858–867. [Google Scholar] [CrossRef]
- Winograd-Katz, S.E.; Fässler, R.; Geiger, B.; Legate, K.R. The Integrin Adhesome: From Genes and Proteins to Human Disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 273–288. [Google Scholar] [CrossRef]
- Singer, I.I.; Scott, S.; Kawka, D.W.; Kazazis, D.M. Adhesomes: Specific Granules Containing Receptors for Laminin, C3bi/Fibrinogen, Fibronectin, and Vitronectin in Human Polymorphonuclear Leukocytes and Monocytes. J. Cell Biol. 1989, 109, 3169–3182. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, C.A.; Bergeron, K.-F.; Whittle, J.; Brandhorst, B.P.; Burke, R.D.; Hynes, R.O. The Echinoderm Adhesome. Dev. Biol. 2006, 300, 252–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemet, G.; Hamidi, H.; Ivaska, J. Filopodia in Cell Adhesion, 3D Migration and Cancer Cell Invasion. Curr. Opin. Cell Biol. 2015, 36, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Innocenti, M. New Insights into the Formation and the Function of Lamellipodia and Ruffles in Mesenchymal Cell Migration. Cell Adhes. Migr. 2018, 12, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Burridge, K.; Guilluy, C. Focal Adhesions, Stress Fibers and Mechanical Tension. Exp. Cell Res. 2016, 343, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Alonso, F.; Spuul, P.; Daubon, T.; Kramer, I.; Génot, E. Variations on the Theme of Podosomes: A Matter of Context. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Cmoch, A.; Groves, P.; Pikuła, S. Biogenesis of Invadopodia and Their Cellular Functions. Postepy Biochem. 2014, 60, 62–68. [Google Scholar]
- Phng, L.-K.; Gebala, V.; Bentley, K.; Philippides, A.; Wacker, A.; Mathivet, T.; Sauteur, L.; Stanchi, F.; Belting, H.-G.; Affolter, M.; et al. Formin-Mediated Actin Polymerization at Endothelial Junctions Is Required for Vessel Lumen Formation and Stabilization. Dev. Cell 2015, 32, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahasrabudhe, A.; Ghate, K.; Mutalik, S.; Jacob, A.; Ghose, A. Formin 2 Regulates the Stabilization of Filopodial Tip Adhesions in Growth Cones and Affects Neuronal Outgrowth and Pathfinding In Vivo. Development 2016, 143, 449–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grobe, H.; Wüstenhagen, A.; Baarlink, C.; Grosse, R.; Grikscheit, K. A Rac1-FMNL2 Signaling Module Affects Cell-Cell Contact Formation Independent of Cdc42 and Membrane Protrusions. PLoS ONE 2018, 13, e0194716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disanza, A.; Bisi, S.; Winterhoff, M.; Milanesi, F.; Ushakov, D.S.; Kast, D.; Marighetti, P.; Romet-Lemonne, G.; Müller, H.-M.; Nickel, W.; et al. CDC42 Switches IRSp53 from Inhibition of Actin Growth to Elongation by Clustering of VASP. EMBO J. 2013, 32, 2735–2750. [Google Scholar] [CrossRef] [Green Version]
- Chacón, M.R.; Navarro, A.I.; Cuesto, G.; del Pino, I.; Scott, R.; Morales, M.; Rico, B. Focal Adhesion Kinase Regulates Actin Nucleation and Neuronal Filopodia Formation during Axonal Growth. Development 2012, 139, 3200–3210. [Google Scholar] [CrossRef] [Green Version]
- Young, L.E.; Latario, C.J.; Higgs, H.N. Roles for Ena/VASP Proteins in FMNL3-Mediated Filopodial Assembly. J. Cell Sci. 2018, 131, jcs220814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzik, M.; McClain, L.M.; Gupton, S.L.; Gertler, F.B. Ena/VASP Regulates MDia2-Initiated Filopodial Length, Dynamics, and Function. Mol. Biol. Cell 2014, 25, 2604–2619. [Google Scholar] [CrossRef] [Green Version]
- Alieva, N.O.; Efremov, A.K.; Hu, S.; Oh, D.; Chen, Z.; Natarajan, M.; Ong, H.T.; Jégou, A.; Romet-Lemonne, G.; Groves, J.T.; et al. Myosin IIA and Formin Dependent Mechanosensitivity of Filopodia Adhesion. Nat. Commun. 2019, 10, 3593. [Google Scholar] [CrossRef] [Green Version]
- Hogg, N.; Laschinger, M.; Giles, K.; McDowall, A. T-Cell Integrins: More than Just Sticking Points. J. Cell Sci. 2003, 116, 4695–4705. [Google Scholar] [CrossRef] [Green Version]
- Pröls, F.; Sagar; Scaal, M. Signaling Filopodia in Vertebrate Embryonic Development. Cell. Mol. Life Sci. 2016, 73, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Romero, S.; Quatela, A.; Bornschlögl, T.; Guadagnini, S.; Bassereau, P.; Van Nhieu, G.T. Filopodium Retraction Is Controlled by Adhesion to Its Tip. J. Cell Sci. 2012, 125, 4999–5004. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.; Guo, W.-H.; Wang, Y.-L. Fibroblasts Probe Substrate Rigidity with Filopodia Extensions before Occupying an Area. Proc. Natl. Acad. Sci. USA 2014, 111, 17176–17181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinardi, L.; Rietdorf, J.; Nitsch, L.; Bono, M.; Tacchetti, C.; Way, M.; Marchisio, P.C. A Dynamic Podosome-like Structure of Epithelial Cells. Exp. Cell Res. 2004, 295, 360–374. [Google Scholar] [CrossRef]
- Wang, Y.; McNiven, M.A. Invasive Matrix Degradation at Focal Adhesions Occurs via Protease Recruitment by a FAK–P130Cas Complex. J. Cell Biol. 2012, 196, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Luxenburg, C.; Geblinger, D.; Klein, E.; Anderson, K.; Hanein, D.; Geiger, B.; Addadi, L. The Architecture of the Adhesive Apparatus of Cultured Osteoclasts: From Podosome Formation to Sealing Zone Assembly. PLoS ONE 2007, 2, e179. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.P.; Eddy, R.; Entenberg, D.; Kai, M.; Gertler, F.B.; Condeelis, J. Tks5 and SHIP2 Regulate Invadopodium Maturation, but Not Initiation, in Breast Carcinoma Cells. Curr. Biol. 2013, 23, 2079–2089. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.; Ebenezer, D.L.; Berdyshev, E.V.; Bronova, I.A.; Shaaya, M.; Harijith, A.; Natarajan, V. Role of Sphingosine Kinase 1 and S1P Transporter Spns2 in HGF-Mediated Lamellipodia Formation in Lung Endothelium. J. Biol. Chem. 2016, 291, 27187–27203. [Google Scholar] [CrossRef] [Green Version]
- Usatyuk, P.V.; Fu, P.; Mohan, V.; Epshtein, Y.; Jacobson, J.R.; Gomez-Cambronero, J.; Wary, K.K.; Bindokas, V.; Dudek, S.M.; Salgia, R.; et al. Role of C-Met/Phosphatidylinositol 3-Kinase (PI3k)/Akt Signaling in Hepatocyte Growth Factor (HGF)-Mediated Lamellipodia Formation, Reactive Oxygen Species (ROS) Generation, and Motility of Lung Endothelial Cells. J. Biol. Chem. 2014, 289, 13476–13491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Kunnumakkara, A.B.; Aggarwal, B.B.; Shakibaei, M. Induction of the Epithelial-to-Mesenchymal Transition of Human Colorectal Cancer by Human TNF-β (Lymphotoxin) and Its Reversal by Resveratrol. Nutrients 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazquez, E.; Watanabe, Y.; Broders-Bondon, F.; Paul-Gilloteaux, P.; Heysch, J.; Baral, V.; Bondurand, N.; Dufour, S. Endothelin-3 Stimulates Cell Adhesion and Cooperates with Β1-Integrins during Enteric Nervous System Ontogenesis. Sci. Rep. 2016, 6, 37877. [Google Scholar] [CrossRef] [PubMed]
- Savin, V.J.; Sharma, M.; Zhou, J.; Gennochi, D.; Fields, T.; Sharma, R.; McCarthy, E.T.; Srivastava, T.; Domen, J.; Tormo, A.; et al. Renal and Hematological Effects of CLCF-1, a B-Cell-Stimulating Cytokine of the IL-6 Family. J. Immunol. Res. 2015, 2015, 714964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiso, M.; Tanaka, S.; Saji, S.; Toi, M.; Sato, F. Long Isoform of VEGF Stimulates Cell Migration of Breast Cancer by Filopodia Formation via NRP1/ARHGAP17/Cdc42 Regulatory Network. Int. J. Cancer 2018, 143, 2905–2918. [Google Scholar] [CrossRef]
- Feng, H.; Chen, L.; Wang, Q.; Shen, B.; Liu, L.; Zheng, P.; Xu, S.; Liu, X.; Chen, J.; Teng, J. Calumenin-15 Facilitates Filopodia Formation by Promoting TGF-β Superfamily Cytokine GDF-15 Transcription. Cell Death Dis. 2013, 4, e870. [Google Scholar] [CrossRef] [Green Version]
- Maisel, S.; Broka, D.; Schroeder, J. Intravesicular Epidermal Growth Factor Receptor Subject to Retrograde Trafficking Drives Epidermal Growth Factor-Dependent Migration. Oncotarget 2017, 9, 6463–6477. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.-D.; Liao, Y.-C.; Shih, Y.-W.; Tsai, L.-Y. Nobiletin Attenuates Metastasis via Both ERK and PI3K/Akt Pathways in HGF-Treated Liver Cancer HepG2 Cells. Phytomedicine 2013, 20, 743–752. [Google Scholar] [CrossRef]
- Marivin, A.; Berthelet, J.; Cartier, J.; Paul, C.; Gemble, S.; Morizot, A.; Boireau, W.; Saleh, M.; Bertoglio, J.; Solary, E.; et al. CIAP1 Regulates TNF-Mediated Cdc42 Activation and Filopodia Formation. Oncogene 2014, 33, 5534–5545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oswald, J.; Büttner, M.; Jasinski-Bergner, S.; Jacobs, R.; Rosenstock, P.; Kielstein, H. Leptin Affects Filopodia and Cofilin in NK-92 Cells in a Dose- and Time-Dependent Manner. Eur. J. Histochem. 2018, 62, 2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigiracciolo, D.C.; Santolla, M.F.; Lappano, R.; Vivacqua, A.; Cirillo, F.; Galli, G.R.; Talia, M.; Muglia, L.; Pellegrino, M.; Nohata, N.; et al. Focal Adhesion Kinase (FAK) Activation by Estrogens Involves GPER in Triple-Negative Breast Cancer Cells. J. Exp. Clin. Cancer Res. 2019, 38, 58. [Google Scholar] [CrossRef] [Green Version]
- Nalluri, S.M.; O’Connor, J.W.; Virgi, G.A.; Stewart, S.E.; Ye, D.; Gomez, E.W. TGFβ1-Induced Expression of Caldesmon Mediates Epithelial–Mesenchymal Transition. Cytoskeleton 2018, 75, 201–212. [Google Scholar] [CrossRef]
- Johansson, M.W.; Khanna, M.; Bortnov, V.; Annis, D.S.; Nguyen, C.L.; Mosher, D.F. IL-5-Stimulated Eosinophils Adherent to Periostin Undergo Stereotypic Morphological Changes and ADAM8-Dependent Migration. Clin. Exp. Allergy 2017, 47, 1263–1274. [Google Scholar] [CrossRef]
- Daubon, T.; Spuul, P.; Alonso, F.; Fremaux, I.; Génot, E. VEGF-A Stimulates Podosome-Mediated Collagen-IV Proteolysis in Microvascular Endothelial Cells. J. Cell Sci. 2016, 129, 2586–2598. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.S.; Killebrew, D.A.; Clary, G.P.; Seawell, J.A.; Meeker, R.B. Differential Regulation of Macrophage Phenotype by Mature and Pro-Nerve Growth Factor. J. Neuroimmunol. 2015, 285, 76–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serafino, A.; Andreola, F.; Pittaluga, E.; Krasnowska, E.K.; Nicotera, G.; Sferrazza, G.; Sinibaldi Vallebona, P.; Pierimarchi, P.; Garaci, E. Thymosin A1 Modifies Podosome Architecture and Promptly Stimulates the Expression of Podosomal Markers in Mature Macrophages. Expert Opin. Biol. Ther. 2015, 15, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Kung, M.-L.; Tsai, H.-E.; Hu, T.-H.; Kuo, H.-M.; Liu, L.-F.; Chen, S.-C.; Lin, P.-R.; Ma, Y.-L.; Wang, E.-M.; Liu, G.-S.; et al. Hepatoma-Derived Growth Factor Stimulates Podosome Rosettes Formation in NIH/3T3 Cells through the Activation of Phosphatidylinositol 3-Kinase/Akt Pathway. Biochem. Biophys. Res. Commun. 2012, 425, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Le Roux-Goglin, E.; Varon, C.; Spuul, P.; Asencio, C.; Mégraud, F.; Génot, E. Helicobacter Infection Induces Podosome Assembly in Primary Hepatocytes In Vitro. Eur. J. Cell Biol. 2012, 91, 161–170. [Google Scholar] [CrossRef]
- Tatin, F.; Grise, F.; Reuzeau, E.; Genot, E.; Moreau, V. Sodium Fluoride Induces Podosome Formation in Endothelial Cells. Biol. Cell 2010, 102, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sa, G.; Liu, Z.; Ren, J.; Wan, Q.; Xiong, X.; Yu, Z.; Chen, H.; Zhao, Y.; He, S. Keratinocyte Growth Factor (KGF) Induces Podosome Formation via Integrin-Erk1/2 Signaling in Human Immortalized Oral Epithelial Cells. Cell. Signal. 2019, 61, 39–47. [Google Scholar] [CrossRef]
- Griera, M.; Martin-Villar, E.; Banon-Rodríguez, I.; Blundell, M.P.; Jones, G.E.; Anton, I.M.; Thrasher, A.J.; Rodriguez-Puyol, M.; Calle, Y. Integrin Linked Kinase (ILK) Regulates Podosome Maturation and Stability in Dendritic Cells. Int. J. Biochem. Cell Biol. 2014, 50, 47–54. [Google Scholar] [CrossRef]
- Mu, X.; Wang, X.; Huang, W.; Wang, R.-T.; Essandoh, K.; Li, Y.; Pugh, A.M.; Peng, J.; Deng, S.; Wang, Y.; et al. Circulating Exosomes Isolated from Septic Mice Induce Cardiovascular Hyperpermeability Through Promoting Podosome Cluster Formation. Shock 2018, 49, 429–441. [Google Scholar] [CrossRef]
- Singh, A.; Gill, G.; Kaur, H.; Amhmed, M.; Jakhu, H. Role of Osteopontin in Bone Remodeling and Orthodontic Tooth Movement: A Review. Prog. Orthod. 2018, 19, 18. [Google Scholar] [CrossRef]
- Mader, C.C.; Oser, M.; Magalhaes, M.A.O.; Bravo-Cordero, J.J.; Condeelis, J.; Koleske, A.J.; Gil-Henn, H. An EGFR–Src–Arg–Cortactin Pathway Mediates Functional Maturation of Invadopodia and Breast Cancer Cell Invasion. Cancer Res. 2011, 71, 1730–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, M.A.; Lwin, T.M.; Chang, A.T.; Kim, J.; Danis, E.; Ohno-Machado, L.; Yang, J. Twist1-Induced Invadopodia Formation Promotes Tumor Metastasis. Cancer Cell 2011, 19, 372–386. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, J.; Tumbarello, D.A.; Schmidt, R.P.; Turner, C.E. Hic-5 Promotes Invadopodia Formation and Invasion during TGF-β–Induced Epithelial–Mesenchymal Transition. J. Cell Biol. 2012, 197, 421–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, D.; Branch, K.M.; Weaver, A.M. Signaling Inputs to Invadopodia and Podosomes. J. Cell Sci. 2013, 126, 2979–2989. [Google Scholar] [CrossRef] [Green Version]
- Rajadurai, C.V.; Havrylov, S.; Zaoui, K.; Vaillancourt, R.; Stuible, M.; Naujokas, M.; Zuo, D.; Tremblay, M.L.; Park, M. Met Receptor Tyrosine Kinase Signals through a Cortactin–Gab1 Scaffold Complex, to Mediate Invadopodia. J. Cell Sci. 2012, 125, 2940–2953. [Google Scholar] [CrossRef] [Green Version]
- Makowiecka, A.; Simiczyjew, A.; Nowak, D.; Mazur, A.J. Varying Effects of EGF, HGF and TGFβ on Formation of Invadopodia and Invasiveness of Melanoma Cell Lines of Different Origin. Eur. J. Histochem. 2016, 60, 2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhu, M.; Yu, S.; Hai, L.; Zhang, L.; Zhang, C.; Zhao, P.; Zhou, H.; Wang, S.; Yang, X. Arg Kinase Mediates CXCL12/CXCR4-Induced Invadopodia Formation and Invasion of Glioma Cells. Exp. Cell Res. 2020, 389, 216–229. [Google Scholar] [CrossRef]
- Love, A.M.; Prince, D.J.; Jessen, J.R. Vangl2-Dependent Regulation of Membrane Protrusions and Directed Migration Requires a Fibronectin Extracellular Matrix. Development 2018, 145, dev165472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R. Central Role of PI3K-SYK Interaction in Fibrinogen-Induced Lamellipodia and Filopodia Formation in Platelets. FEBS Open Bio. 2016, 6, 1285–1296. [Google Scholar] [CrossRef]
- Wang, W.Y.; Pearson, A.T.; Kutys, M.L.; Choi, C.K.; Wozniak, M.A.; Baker, B.M.; Chen, C.S. Extracellular Matrix Alignment Dictates the Organization of Focal Adhesions and Directs Uniaxial Cell Migration. APL Bioeng. 2018, 2, 046107. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.; Neubauer, S.; Rechenmacher, F.; Kessler, H.; Missirlis, D. Recruitment of ανβ3 Integrin to α5β1 Integrin-Induced Clusters Enables Focal Adhesion Maturation and Cell Spreading. J. Cell Sci. 2020, 133, jcs232702. [Google Scholar] [CrossRef]
- Stritt, S.; Thielmann, I.; Dütting, S.; Stegner, D.; Nieswandt, B. Phospholipase D Is a Central Regulator of Collagen I-Induced Cytoskeletal Rearrangement and Podosome Formation in Megakaryocytes. J. Thromb. Haemost. 2014, 12, 1364–1371. [Google Scholar] [CrossRef]
- Schachtner, H.; Calaminus, S.D.J.; Sinclair, A.; Monypenny, J.; Blundell, M.P.; Leon, C.; Holyoake, T.L.; Thrasher, A.J.; Michie, A.M.; Vukovic, M.; et al. Megakaryocytes Assemble Podosomes That Degrade Matrix and Protrude through Basement Membrane. Blood 2013, 121, 2542–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Shoji, S.; Yamamoto, K.; Nada, S.; Okada, M.; Yamamoto, T.; Honda, Z. Essential Roles of Lyn in Fibronectin-Mediated Filamentous Actin Assembly and Cell Motility in Mast Cells. J. Immunol. 1998, 161, 3694–3701. [Google Scholar] [PubMed]
- Beaty, B.T.; Sharma, V.P.; Bravo-Cordero, J.J.; Simpson, M.A.; Eddy, R.J.; Koleske, A.J.; Condeelis, J. Β1 Integrin Regulates Arg to Promote Invadopodial Maturation and Matrix Degradation. MBoC 2013, 24, 1661–1675. [Google Scholar] [CrossRef]
- Artym, V.V.; Swatkoski, S.; Matsumoto, K.; Campbell, C.B.; Petrie, R.J.; Dimitriadis, E.K.; Li, X.; Mueller, S.C.; Bugge, T.H.; Gucek, M.; et al. Dense Fibrillar Collagen Is a Potent Inducer of Invadopodia via a Specific Signaling Network. J. Cell Biol. 2015, 208, 331–350. [Google Scholar] [CrossRef]
- Di Martino, J.; Moreau, V.; Saltel, F. Type I Collagen Fibrils: An Inducer of Invadosomes. Oncotarget 2015, 6, 28519–28520. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xu, Y.; Wei, Y.; Qiu, Q.; Chew, T.-L.; Kang, Y.; Cheng, C. The CD44s Splice Isoform Is a Central Mediator for Invadopodia Activity. J. Cell Sci. 2016, 129, 1355–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burridge, K. Focal Adhesions: A Personal Perspective on a Half Century of Progress. FEBS J. 2017, 284, 3355–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, Y.-L.; Chen, L.-C.; Shen, T.-L. Emerging Roles of Focal Adhesion Kinase in Cancer. Biomed. Res. Int. 2015, 2015, 690690. [Google Scholar] [CrossRef]
- Tomakidi, P.; Schulz, S.; Proksch, S.; Weber, W.; Steinberg, T. Focal Adhesion Kinase (FAK) Perspectives in Mechanobiology: Implications for Cell Behaviour. Cell Tissue Res. 2014, 357, 515–526. [Google Scholar] [CrossRef]
- Schlaepfer, D.D.; Mitra, S.K.; Ilic, D. Control of Motile and Invasive Cell Phenotypes by Focal Adhesion Kinase. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1692, 77–102. [Google Scholar] [CrossRef]
- Sarangi, B.R.; Gupta, M.; Doss, B.L.; Tissot, N.; Lam, F.; Mège, R.-M.; Borghi, N.; Ladoux, B. Coordination between Intra- and Extracellular Forces Regulates Focal Adhesion Dynamics. Nano Lett. 2017, 17, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.; Artym, V.V.; Green, J.A.; Yamada, K.M. The Matrix Reorganized: Extracellular Matrix Remodeling and Integrin Signaling. Curr. Opin. Cell Biol. 2006, 18, 463–471. [Google Scholar] [CrossRef]
- Hu, Y.-L.; Lu, S.; Szeto, K.W.; Sun, J.; Wang, Y.; Lasheras, J.C.; Chien, S. FAK and Paxillin Dynamics at Focal Adhesions in the Protrusions of Migrating Cells. Sci. Rep. 2014, 4, 6024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciobanasu, C.; Faivre, B.; Le Clainche, C. Integrating Actin Dynamics, Mechanotransduction and Integrin Activation: The Multiple Functions of Actin Binding Proteins in Focal Adhesions. Eur. J. Cell Biol. 2013, 92, 339–348. [Google Scholar] [CrossRef]
- Stutchbury, B.; Atherton, P.; Tsang, R.; Wang, D.-Y.; Ballestrem, C. Distinct Focal Adhesion Protein Modules Control Different Aspects of Mechanotransduction. J. Cell Sci. 2017, 130, 1612–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.W.; Lee, T.T.; Weng, S.; Fu, J.; García, A.J. Effects of Substrate Stiffness and Actomyosin Contractility on Coupling between Force Transmission and Vinculin-Paxillin Recruitment at Single Focal Adhesions. Mol. Biol. Cell 2017, 28, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Grashoff, C.; Hoffman, B.D.; Brenner, M.D.; Zhou, R.; Parsons, M.; Yang, M.T.; McLean, M.A.; Sligar, S.G.; Chen, C.S.; Ha, T.; et al. Measuring Mechanical Tension across Vinculin Reveals Regulation of Focal Adhesion Dynamics. Nature 2010, 466, 263–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atherton, P.; Stutchbury, B.; Wang, D.-Y.; Jethwa, D.; Tsang, R.; Meiler-Rodriguez, E.; Wang, P.; Bate, N.; Zent, R.; Barsukov, I.L.; et al. Vinculin Controls Talin Engagement with the Actomyosin Machinery. Nat. Commun. 2015, 6, 10038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omachi, T.; Ichikawa, T.; Kimura, Y.; Ueda, K.; Kioka, N. Vinculin Association with Actin Cytoskeleton Is Necessary for Stiffness-Dependent Regulation of Vinculin Behavior. PLoS ONE 2017, 12, e0175324. [Google Scholar] [CrossRef]
- Calderwood, D.A.; Campbell, I.D.; Critchley, D.R. Talins and Kindlins: Partners in Integrin-Mediated Adhesion. Nat. Rev. Mol. Cell Biol. 2013, 14, 503–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Z.C.-K.; Kwan, H.-L.R.; Wong, Y.S.; Jiang, Z.; Zhou, Z.; Tam, K.W.; Chan, Y.-S.; Chan, C.B.; Lee, C.W. Site-Directed MT1-MMP Trafficking and Surface Insertion Regulate AChR Clustering and Remodeling at Developing NMJs. eLife 2020, 9, e54379. [Google Scholar] [CrossRef]
- El Azzouzi, K.; Wiesner, C.; Linder, S. Metalloproteinase MT1-MMP Islets Act as Memory Devices for Podosome Reemergence. J. Cell Biol. 2016, 213, 109–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infante, E.; Castagnino, A.; Ferrari, R.; Monteiro, P.; Agüera-González, S.; Paul-Gilloteaux, P.; Domingues, M.J.; Maiuri, P.; Raab, M.; Shanahan, C.M.; et al. LINC Complex-Lis1 Interplay Controls MT1-MMP Matrix Digest-on-Demand Response for Confined Tumor Cell Migration. Nat. Commun. 2018, 9, 2443. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.-L.; Chen, C.-L.; Pan, Y.-R.; Chiu, W.-T.; Chen, H.-C. Biogenesis of Podosome Rosettes through Fission. Sci. Rep. 2018, 8, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Dries, K.; Schwartz, S.L.; Byars, J.; Meddens, M.B.M.; Bolomini-Vittori, M.; Lidke, D.S.; Figdor, C.G.; Lidke, K.A.; Cambi, A. Dual-Color Superresolution Microscopy Reveals Nanoscale Organization of Mechanosensory Podosomes. Mol. Biol. Cell 2013, 24, 2112–2123. [Google Scholar] [CrossRef]
- Shattil, S.J.; O’Toole, T.; Eigenthaler, M.; Thon, V.; Williams, M.; Babior, B.M.; Ginsberg, M.H. Beta 3-Endonexin, a Novel Polypeptide That Interacts Specifically with the Cytoplasmic Tail of the Integrin Beta 3 Subunit. J. Cell Biol. 1995, 131, 807–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, R.M.; Li, C.; Bhattacharya, S.; Naren, A.P.; Johnson, L.R. Spermine, a Molecular Switch Regulating EGFR, Integrin Β3, Src, and FAK Scaffolding. Cell. Signal. 2012, 24, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Proag, A.; Bouissou, A.; Mangeat, T.; Voituriez, R.; Delobelle, P.; Thibault, C.; Vieu, C.; Maridonneau-Parini, I.; Poincloux, R. Working Together: Spatial Synchrony in the Force and Actin Dynamics of Podosome First Neighbors. ACS Nano 2015, 9, 3800–3813. [Google Scholar] [CrossRef]
- Bhuwania, R.; Cornfine, S.; Fang, Z.; Krüger, M.; Luna, E.J.; Linder, S. Supervillin Couples Myosin-Dependent Contractility to Podosomes and Enables Their Turnover. J. Cell Sci. 2012, 125, 2300–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, B.; Willemse, M.; Fransen, J.; Cambi, A.; van den Dries, K. Super-Resolution Correlative Light and Electron Microscopy (SR-CLEM) Reveals Novel Ultrastructural Insights Into Dendritic Cell Podosomes. Front. Immunol. 2018, 9, 1908. [Google Scholar] [CrossRef]
- Cervero, P.; Wiesner, C.; Bouissou, A.; Poincloux, R.; Linder, S. Lymphocyte-Specific Protein 1 Regulates Mechanosensory Oscillation of Podosomes and Actin Isoform-Based Actomyosin Symmetry Breaking. Nat. Commun. 2018, 9, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petropoulos, C.; Oddou, C.; Emadali, A.; Hiriart-Bryant, E.; Boyault, C.; Faurobert, E.; Vande Pol, S.; Kim-Kaneyama, J.-R.; Kraut, A.; Coute, Y.; et al. Roles of Paxillin Family Members in Adhesion and ECM Degradation Coupling at Invadosomes. J. Cell Biol. 2016, 213, 585–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walde, M.; Monypenny, J.; Heintzmann, R.; Jones, G.E.; Cox, S. Vinculin Binding Angle in Podosomes Revealed by High Resolution Microscopy. PLoS ONE 2014, 9, e88251. [Google Scholar] [CrossRef] [Green Version]
- Praekelt, U.; Kopp, P.M.; Rehm, K.; Linder, S.; Bate, N.; Patel, B.; Debrand, E.; Manso, A.M.; Ross, R.S.; Conti, F.; et al. New Isoform-Specific Monoclonal Antibodies Reveal Different Sub-Cellular Localisations for Talin1 and Talin2. Eur. J. Cell Biol. 2012, 91, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Labernadie, A.; Thibault, C.; Vieu, C.; Maridonneau-Parini, I.; Charrière, G.M. Dynamics of Podosome Stiffness Revealed by Atomic Force Microscopy. Proc. Natl. Acad. Sci. USA 2010, 107, 21016–21021. [Google Scholar] [CrossRef] [Green Version]
- Horton, M.A.; Nesbit, M.A.; Helfrich, M.H. Interaction of Osteopontin with Osteoclast Integrinsa. Ann. N. Y. Acad. Sci. 1995, 760, 190–200. [Google Scholar] [CrossRef]
- Juin, A.; Billottet, C.; Moreau, V.; Destaing, O.; Albiges-Rizo, C.; Rosenbaum, J.; Génot, E.; Saltel, F. Physiological Type I Collagen Organization Induces the Formation of a Novel Class of Linear Invadosomes. MBoC 2011, 23, 297–309. [Google Scholar] [CrossRef]
- Ferrari, R.; Infante, E.; Chavrier, P. Nucleus–Invadopodia Duo During Cancer Invasion. Trends Cell Biol. 2019, 29, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Castro, A.; Marchesin, V.; Monteiro, P.; Lodillinsky, C.; Rossé, C.; Chavrier, P. Cellular and Molecular Mechanisms of MT1-MMP-Dependent Cancer Cell Invasion. Annu. Rev. Cell Dev. Biol 2016, 3220, 555–576. [Google Scholar] [CrossRef]
- Hastie, E.L.; Sherwood, D.R. A New Front in Cell Invasion: The Invadopodial Membrane. Eur. J. Cell Biol. 2016, 95, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meirson, T.; Gil-Henn, H. Targeting Invadopodia for Blocking Breast Cancer Metastasis. Drug Resist. Updates 2018, 39, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.G.; Correia, I.; Krasavina, O.; Watson, N.; Matsudaira, P. Macrophage Podosomes Assemble at the Leading Lamella by Growth and Fragmentation. J. Cell Biol. 2003, 161, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Calle, Y.; Chou, H.; Thrasher, A.J.; Jones, G.E. Wiskott–Aldrich Syndrome Protein and the Cytoskeletal Dynamics of Dendritic Cells. J. Pathol. 2004, 204, 460–469. [Google Scholar] [CrossRef]
- Levy-Apter, E.; Finkelshtein, E.; Vemulapalli, V.; Li, S.S.-C.; Bedford, M.T.; Elson, A. Adaptor Protein GRB2 Promotes Src Tyrosine Kinase Activation and Podosomal Organization by Protein-Tyrosine Phosphatase ϵ in Osteoclasts. J. Biol. Chem. 2014, 289, 36048–36058. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, S. Requirement for a Complex of Wiskott-Aldrich Syndrome Protein (WASP) with WASP Interacting Protein in Podosome Formation in Macrophages. J. Immunol. 2007, 178, 2987–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pignatelli, J.; Bravo-Cordero, J.J.; Roh-Johnson, M.; Gandhi, S.J.; Wang, Y.; Chen, X.; Eddy, R.J.; Xue, A.; Singer, R.H.; Hodgson, L.; et al. Macrophage-Dependent Tumor Cell Transendothelial Migration Is Mediated by Notch1/MenaINV-Initiated Invadopodium Formation. Sci. Rep. 2016, 6, 37874. [Google Scholar] [CrossRef]
- Oser, M.; Dovas, A.; Cox, D.; Condeelis, J. Nck1 and Grb2 Localization Patterns Can Distinguish Invadopodia from Podosomes. Eur. J. Cell Biol. 2011, 90, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Weidmann, M.D.; Surve, C.R.; Eddy, R.J.; Chen, X.; Gertler, F.B.; Sharma, V.P.; Condeelis, J.S. Mena(INV) Dysregulates Cortactin Phosphorylation to Promote Invadopodium Maturation. Sci. Rep. 2016, 6, 36142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.K.; Oudin, M.J.; Tadros, J.; Neil, J.; Del Rosario, A.; Joughin, B.A.; Ritsma, L.; Wyckoff, J.; Vasile, E.; Eddy, R.; et al. PTP1B-Dependent Regulation of Receptor Tyrosine Kinase Signaling by the Actin-Binding Protein Mena. MBoC 2015, 26, 3867–3878. [Google Scholar] [CrossRef] [Green Version]
- Rajadurai, C.V.; Havrylov, S.; Coelho, P.P.; Ratcliffe, C.D.H.; Zaoui, K.; Huang, B.H.; Monast, A.; Chughtai, N.; Sangwan, V.; Gertler, F.B.; et al. 5′-Inositol Phosphatase SHIP2 Recruits Mena to Stabilize Invadopodia for Cancer Cell Invasion. J. Cell Biol. 2016, 214, 719–734. [Google Scholar] [CrossRef] [Green Version]
- Gorai, S.; Paul, D.; Haloi, N.; Borah, R.; Santra, M.K.; Manna, D. Mechanistic Insights into the Phosphatidylinositol Binding Properties of the Pleckstrin Homology Domain of Lamellipodin. Mol. BioSyst. 2016, 12, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Cordero, J.J.; Magalhaes, M.A.O.; Eddy, R.J.; Hodgson, L.; Condeelis, J. Functions of Cofilin in Cell Locomotion and Invasion. Nat. Rev. Mol. Cell Biol. 2013, 14, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Beaty, B.T.; Condeelis, J. Digging a Little Deeper: The Stages of Invadopodium Formation and Maturation. Eur. J. Cell Biol. 2014, 93, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.R.; Owen, D.; Mott, H.R. Cdc42 in Actin Dynamics: An Ordered Pathway Governed by Complex Equilibria and Directional Effector Handover. Small GTPases 2017, 8, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, D.A.; Courtneidge, S.A. The “ins” and “Outs” of Podosomes and Invadopodia: Characteristics, Formation and Function. Nat. Rev. Mol. Cell Biol. 2011, 12, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Jacob, A.; Linklater, E.; Bayless, B.A.; Lyons, T.; Prekeris, R. The Role and Regulation of Rab40b-Tks5 Complex during Invadopodia Formation and Cancer Cell Invasion. J. Cell Sci. 2016, 129, 4341–4353. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.C.; McNeilly, R.E.; Coppolino, M.G. SNAP23, Syntaxin4, and Vesicle-Associated Membrane Protein 7 (VAMP7) Mediate Trafficking of Membrane Type 1-Matrix Metalloproteinase (MT1-MMP) during Invadopodium Formation and Tumor Cell Invasion. Mol. Biol Cell. 2014, 25, 2061–2070. [Google Scholar] [CrossRef]
- Steffen, A.; Le Dez, G.; Poincloux, R.; Recchi, C.; Nassoy, P.; Rottner, K.; Galli, T.; Chavrier, P. MT1-MMP-Dependent Invasion Is Regulated by TI-VAMP/VAMP7. Curr. Biol. 2008, 18, 926–931. [Google Scholar] [CrossRef]
- Vieira, L.F.D.A.; Lins, M.P.; Viana, I.M.M.N.; Dos Santos, J.E.; Smaniotto, S.; Reis, M.D.D.S. Metallic Nanoparticles Reduce the Migration of Human Fibroblasts In Vitro. Nanoscale Res. Lett. 2017, 12, 200. [Google Scholar] [CrossRef] [Green Version]
- Lo, H.-M.; Ma, M.-C.; Shieh, J.; Chen, H.-L.; Wu, W. Naked Physically Synthesized Gold Nanoparticles Affect Migration, Mitochondrial Activity, and Proliferation of Vascular Smooth Muscle Cells. Int. J. Nanomed. 2018, 13, 3163–3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Královec, K.; Havelek, R.; Koutová, D.; Veverka, P.; Kubíčková, L.; Brázda, P.; Kohout, J.; Herynek, V.; Vosmanská, M.; Kaman, O. Magnetic Nanoparticles of Ga-Substituted ε-Fe2O3 for Biomedical Applications: Magnetic Properties, Transverse Relaxivity, and Effects of Silica-Coated Particles on Cytoskeletal Networks. J. Biomed. Mater. Res. Part A 2020, 108, 1563–1578. [Google Scholar] [CrossRef] [PubMed]
- Sheykhzadeh, S.; Luo, M.; Peng, B.; White, J.; Abdalla, Y.; Tang, T.; Mäkilä, E.; Voelcker, N.H.; Tong, W.Y. Transferrin-Targeted Porous Silicon Nanoparticles Reduce Glioblastoma Cell Migration across Tight Extracellular Space. Sci. Rep. 2020, 10, 2320. [Google Scholar] [CrossRef] [Green Version]
- Popara, J.; Accomasso, L.; Vitale, E.; Gallina, C.; Roggio, D.; Iannuzzi, A.; Raimondo, S.; Rastaldo, R.; Alberto, G.; Catalano, F.; et al. Silica Nanoparticles Actively Engage with Mesenchymal Stem Cells in Improving Acute Functional Cardiac Integration. Nanomedicine 2018, 13, 1121–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenific, C.M.; Wittmann, T.; Debnath, J. Autophagy in Adhesion and Migration. J. Cell Sci. 2016, 129, 3685–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi, M.N.; Mowers, E.E.; Drake, L.E.; Collier, C.; Chen, H.; Zamora, M.; Mui, S.; Macleod, K.F. Autophagy Promotes Focal Adhesion Disassembly and Cell Motility of Metastatic Tumor Cells through the Direct Interaction of Paxillin with LC3. Cell Rep. 2016, 15, 1660–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenific, C.M.; Stehbens, S.J.; Goldsmith, J.; Leidal, A.M.; Faure, N.; Ye, J.; Wittmann, T.; Debnath, J. NBR1 Enables Autophagy-Dependent Focal Adhesion Turnover. J. Cell Biol. 2016, 212, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Dower, C.M.; Wills, C.A.; Frisch, S.M.; Wang, H.-G. Mechanisms and Context Underlying the Role of Autophagy in Cancer Metastasis. Autophagy 2018, 14, 1110–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yin, S.; Zhang, L.; Shi, K.; Tang, J.; Zhang, Z.; He, Q. A Tumor-Activatable Particle with Antimetastatic Potential in Breast Cancer via Inhibiting the Autophagy-Dependent Disassembly of Focal Adhesion. Biomaterials 2018, 168, 1–9. [Google Scholar] [CrossRef]
- Shin, T.H.; Lee, D.Y.; Ketebo, A.A.; Lee, S.; Manavalan, B.; Basith, S.; Ahn, C.; Kang, S.H.; Park, S.; Lee, G. Silica-Coated Magnetic Nanoparticles Decrease Human Bone Marrow-Derived Mesenchymal Stem Cell Migratory Activity by Reducing Membrane Fluidity and Impairing Focal Adhesion. Nanomaterials 2019, 9, 1475. [Google Scholar] [CrossRef] [Green Version]
- Mulens-Arias, V.; Rojas, J.M.; Sanz-Ortega, L.; Portilla, Y.; Pérez-Yagüe, S.; Barber, D.F. Polyethylenimine-Coated Superparamagnetic Iron Oxide Nanoparticles Impair in Vitro and in Vivo Angiogenesis. Nanomed. Nanotechnol. Biol. Med. 2019, 21, 102063. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Hartmann, R.; De Aberasturi, D.J.; Yang, F.; Soenen, S.J.H.; Manshian, B.B.; Franz, J.; Valdeperez, D.; Pelaz, B.; Feliu, N.; et al. Colloidal Gold Nanoparticles Induce Changes in Cellular and Subcellular Morphology. ACS Nano 2017, 11, 7807–7820. [Google Scholar] [CrossRef]
- Královec, K.; Melounková, L.; Slováková, M.; Mannová, N.; Sedlák, M.; Bartáček, J.; Havelek, R. Disruption of Cell Adhesion and Cytoskeletal Networks by Thiol-Functionalized Silica-Coated Iron Oxide Nanoparticles. Int. J. Mol. Sci. 2020, 21, 9350. [Google Scholar] [CrossRef] [PubMed]
- Ketebo, A.A.; Shin, T.H.; Jun, M.; Lee, G.; Park, S. Effect of Silica-Coated Magnetic Nanoparticles on Rigidity Sensing of Human Embryonic Kidney Cells. J. Nanobiotechnology 2020, 18, 170. [Google Scholar] [CrossRef] [PubMed]
- Marcus, J.; Bejerano-Sagie, M.; Patterson, N.; Bagchi, S.; Verkhusha, V.V.; Connolly, D.; Goldberg, G.L.; Golden, A.; Sharma, V.P.; Condeelis, J.; et al. Septin 9 Isoforms Promote Tumorigenesis in Mammary Epithelial Cells by Increasing Migration and ECM Degradation through Metalloproteinase Secretion at Focal Adhesions. Oncogene 2019, 38, 5839. [Google Scholar] [CrossRef]
- Mulens-Arias, V.; Balfourier, A.; Nicolás-Boluda, A.; Carn, F.; Gazeau, F. Disturbance of Adhesomes by Gold Nanoparticles Reveals a Size- and Cell Type-Bias. Biomater. Sci. 2019, 7, 389–408. [Google Scholar] [CrossRef]
- Soenen, S.J.H.; Nuytten, N.; De Meyer, S.F.; De Smedt, S.C.; De Cuyper, M. High Intracellular Iron Oxide Nanoparticle Concentrations Affect Cellular Cytoskeleton and Focal Adhesion Kinase-Mediated Signaling. Small 2010, 6, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Tay, C.Y.; Cai, P.; Setyawati, M.I.; Fang, W.; Tan, L.P.; Hong, C.H.L.; Chen, X.; Leong, D.T. Nanoparticles Strengthen Intracellular Tension and Retard Cellular Migration. Nano Lett. 2014, 14, 83–88. [Google Scholar] [CrossRef]
- Discher, D.E.; Janmey, P.; Wang, Y. Tissue Cells Feel and Respond to the Stiffness of Their Substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Ingber, D.E. Tensegrity: The Architectural Basis of Cellular Mechanotransduction. Annu. Rev. Physiol. 1997, 59, 575–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and Extracellular Matrix Homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Fouchard, J.; Bimbard, C.; Bufi, N.; Durand-Smet, P.; Proag, A.; Richert, A.; Cardoso, O.; Asnacios, A. Three-Dimensional Cell Body Shape Dictates the Onset of Traction Force Generation and Growth of Focal Adhesions. Proc. Natl. Acad. Sci. USA 2014, 111, 13075–13080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Plotnikov, S.V.; Moalim, A.Y.; Waterman, C.M.; Liu, J. Two Distinct Actin Networks Mediate Traction Oscillations to Confer Focal Adhesion Mechanosensing. Biophys. J. 2017, 112, 780–794. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.; Schoelermann, J.; Mustafa, K.; Cimpan, M.R. TiO2 Nanoparticles Disrupt Cell Adhesion and the Architecture of Cytoskeletal Networks of Human Osteoblast-like Cells in a Size Dependent Manner. J. Biomed. Mater. Res. Part A 2018, 106, 2582–2593. [Google Scholar] [CrossRef]
- Miyauchi, T.; Yamada, M.; Yamamoto, A.; Iwasa, F.; Suzawa, T.; Kamijo, R.; Baba, K.; Ogawa, T. The Enhanced Characteristics of Osteoblast Adhesion to Photofunctionalized Nanoscale TiO2 Layers on Biomaterials Surfaces. Biomaterials 2010, 31, 3827–3839. [Google Scholar] [CrossRef]
- Lipski, A.M.; Pino, C.J.; Haselton, F.R.; Chen, I.-W.; Shastri, V.P. The Effect of Silica Nanoparticle-Modified Surfaces on Cell Morphology, Cytoskeletal Organization and Function. Biomaterials 2008, 29, 3836–3846. [Google Scholar] [CrossRef] [Green Version]
- Perez, J.E.; Fage, F.; Pereira, D.; Abou-Hassan, A.; Asnacios, S.; Asnacios, A.; Wilhelm, C. Transient Cell Stiffening Triggered by Magnetic Nanoparticle Exposure. J. Nanobiotechnol. 2021, 19, 117. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Wang, H.; Wang, Y.; Ding, K.; Liu, H.; Yuan, L.; Shi, X.; Wang, M.; Wang, Y.; Chen, H. Maintaining the Pluripotency of Mouse Embryonic Stem Cells on Gold Nanoparticle Layers with Nanoscale but Not Microscale Surface Roughness. Nanoscale 2014, 6, 6959. [Google Scholar] [CrossRef]
- Chen, W.; Villa-Diaz, L.G.; Sun, Y.; Weng, S.; Kim, J.K.; Lam, R.H.W.; Han, L.; Fan, R.; Krebsbach, P.H.; Fu, J. Nanotopography Influences Adhesion, Spreading, and Self-Renewal of Human Embryonic Stem Cells. ACS Nano 2012, 6, 4094–4103. [Google Scholar] [CrossRef]
- Azatov, M.; Sun, X.; Suberi, A.; Fourkas, J.T.; Upadhyaya, A. Topography on a Subcellular Scale Modulates Cellular Adhesions and Actin Stress Fiber Dynamics in Tumor Associated Fibroblasts. Phys. Biol. 2017, 14, 065003. [Google Scholar] [CrossRef]
- Bello, D.G.; Fouillen, A.; Badia, A.; Nanci, A. A Nanoporous Titanium Surface Promotes the Maturation of Focal Adhesions and Formation of Filopodia with Distinctive Nanoscale Protrusions by Osteogenic Cells. Acta Biomater. 2017, 60, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Schwieder, M.; Blümmel, J.; Cavalcanti-Adam, E.A.; López-Garcia, M.; Kessler, H.; Geiger, B.; Spatz, J.P. Cell Interactions with Hierarchically Structured Nano-Patterned Adhesive Surfaces. Soft Matter 2009, 5, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abagnale, G.; Steger, M.; Nguyen, V.H.; Hersch, N.; Sechi, A.; Joussen, S.; Denecke, B.; Merkel, R.; Hoffmann, B.; Dreser, A.; et al. Surface Topography Enhances Differentiation of Mesenchymal Stem Cells towards Osteogenic and Adipogenic Lineages. Biomaterials 2015, 61, 316–326. [Google Scholar] [CrossRef]
- Posa, F.; Baha-Schwab, E.H.; Wei, Q.; Di Benedetto, A.; Neubauer, S.; Reichart, F.; Kessler, H.; Spatz, J.P.; Albiges-Rizo, C.; Mori, G.; et al. Surface Co-Presentation of BMP-2 and Integrin Selective Ligands at the Nanoscale Favors A5β1 Integrin-Mediated Adhesion. Biomaterials 2021, 267, 120484. [Google Scholar] [CrossRef]
- Kang, H.; Wong, D.S.H.; Yan, X.; Jung, H.J.; Kim, S.; Lin, S.; Wei, K.; Li, G.; Dravid, V.P.; Bian, L. Remote Control of Multimodal Nanoscale Ligand Oscillations Regulates Stem Cell Adhesion and Differentiation. ACS Nano 2017, 11, 9636–9649. [Google Scholar] [CrossRef]
- Kang, H.; Jung, H.J.; Wong, D.S.H.; Kim, S.K.; Lin, S.; Chan, K.F.; Zhang, L.; Li, G.; Dravid, V.P.; Bian, L. Remote Control of Heterodimeric Magnetic Nanoswitch Regulates the Adhesion and Differentiation of Stem Cells. J. Am. Chem. Soc. 2018, 140, 5909–5913. [Google Scholar] [CrossRef]
- Okada, T.; Ogura, T. Nanoscale Imaging of the Adhesion Core Including Integrin Β1 on Intact Living Cells Using Scanning Electron-Assisted Dielectric-Impedance Microscopy. PLoS ONE 2018, 13, e0204133. [Google Scholar] [CrossRef]
- Rojas, J.M.; Sanz-Ortega, L.; Mulens-Arias, V.; Gutiérrez, L.; Pérez-Yagüe, S.; Barber, D.F. Superparamagnetic Iron Oxide Nanoparticle Uptake Alters M2 Macrophage Phenotype, Iron Metabolism, Migration and Invasion. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Mulens-Arias, V.; Rojas, J.M.; Pérez-Yagüe, S.; Morales, M.P.; Barber, D.F. Polyethylenimine-Coated SPIONs Trigger Macrophage Activation through TLR-4 Signaling and ROS Production and Modulate Podosome Dynamics. Biomaterials 2015, 52, 494–506. [Google Scholar] [CrossRef]
- Akatsuka, S.; Yamashita, Y.; Ohara, H.; Liu, Y.-T.; Izumiya, M.; Abe, K.; Ochiai, M.; Jiang, L.; Nagai, H.; Okazaki, Y.; et al. Fenton Reaction Induced Cancer in Wild Type Rats Recapitulates Genomic Alterations Observed in Human Cancer. PLoS ONE 2012, 7, e43403. [Google Scholar] [CrossRef] [PubMed]
- Brillo, V.; Chieregato, L.; Leanza, L.; Muccioli, S.; Costa, R. Mitochondrial Dynamics, ROS, and Cell Signaling: A Blended Overview. Life 2021, 11, 332. [Google Scholar] [CrossRef]
- Xu, J.; Yang, J.; Nyga, A.; Ehteramyan, M.; Moraga, A.; Wu, Y.; Zeng, L.; Knight, M.M.; Shelton, J.C. Cobalt (II) Ions and Nanoparticles Induce Macrophage Retention by ROS-Mediated down-Regulation of RhoA Expression. Acta Biomater. 2018, 72, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Linder, S.; Aepfelbacher, M. Podosomes: Adhesion Hot-Spots of Invasive Cells. Trends Cell Biol. 2003, 13, 376–385. [Google Scholar] [CrossRef]
- Mulens-Arias, V.; Rojas, J.M.; Pérez-Yagüe, S.; Morales, M.d.P.; Barber, D.F. Polyethylenimine-Coated SPION Exhibits Potential Intrinsic Anti-Metastatic Properties Inhibiting Migration and Invasion of Pancreatic Tumor Cells. J. Control. Release 2015, 216, 78–92. [Google Scholar] [CrossRef]
- Simone, T.M.; Higgins, C.E.; Czekay, R.-P.; Law, B.K.; Higgins, S.P.; Archambeault, J.; Kutz, S.M.; Higgins, P.J. SERPINE1: A Molecular Switch in the Proliferation-Migration Dichotomy in Wound-”Activated” Keratinocytes. Adv. Wound Care 2014, 3, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Shih, Y.-P.; Takada, Y.; Lo, S.H. Silencing of DLC1 Upregulates PAI-1 Expression and Reduces Migration in Normal Prostate Cells. Mol. Cancer Res. 2012, 10, 34–39. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Lamellipodia | Filopodia | Focal Adhesions | Podosomes | Invadopodia | |
|---|---|---|---|---|---|
| Structure | Sheet-like protrusions that attach to ECM driven by branched actin arrangements | Often originating at lamellipodium as finger-like extensions driven by linear actin polymerization | Clusters of transmembrane receptors, integrins, and cytosolic proteins driven by parallel actin filaments branched at the end | A discrete actin-rich core surrounded by a ring of actin-associated and signaling proteins, driven by branched and unbranched actin filaments | A discrete actin-rich core is surrounded by a ring of actin-associated and signaling proteins. Often linked to tumor cells driven by parallel actin filaments within the tip and branched at the base |
| Cellular location | Leading-edge | Embedded within lamellipodia | Leading-edge of the cell | Ventral cell surface, often clustered behind the leading edge of the cell | Ventral cell surface, often situated under the nucleus |
| Dimesions | Width: 0.1–0.2 μm | Width: 0.1–0.3 μm; length: 3–10 μm | Width: 2–6 μm | Width: 0.5–2 μm; length: 0.5–1 μm [45] | Width: 0.5–2 μm; length: >2 μm |
| Pericellular proteolysis | Minimal | No | Minimal [46] | Yes, through MT1MMP and UPAR | Yes, through MMP2, MMP9, MT1MMP, seprase, UPAR, ADAM12, ADAM15, and ADAM19 |
| Duration of structure | Minutes | Minutes | Hours, it depends on the rate of cell migration | Minutes [47] | Hours [48] |
| Soluble stimuli | HGF [49,50], TNFα and TNFβ [51], endothelin-3 [52], CLCF-1 [53] | VEGF-A [54], GDF-5 [55], EGF [56], HGF [57], TNFα [58] and TNFβ [51], leptin [59] | Estrogen [60], TGFβ1 [61], endothelin-3 [52] | IL-5 [62], VEGF-A [63], pro-NGF [64], thymosin α1 [65], hepatoma-derived growth factor (HDGF) [66], TGFβ1 [67], NaF [68], KGF [69], SDF-1α [70], exosomes [71], osteopontin [72] | EGF [73], PDGF [74], TGFβ [75], VEGF [76], HGF [77,78], SDF-1 [79] |
| ECM stimuli | Fibronectin [80], fibrinogen [81] | Fibronectin [80], fibrinogen [81] | Fibronectin [82,83], | Collagen I [84], fibronectin [85,86], fibrinogen [85], | Fibrinogen [87], Collagen I [88,89], hyaluronan [90] |
| Nanoparticle | Cell | FA Markers | Probable Signaling Direction | Time (h) | Effect |
|---|---|---|---|---|---|
| SiO2 | Human mesenchymal stem cells | Vinculin | Inside–out | 16 | Increased FA size and maturation [147] |
| SiO2 (50, 100, 300 nm) | Bovine aortic endothelial cells (BAEC) | F-actin/vinculin | Outside–in | 24 | Decreased FA size [169] |
| SiO2 (50, 100, 300 nm) | Mouse calvarial preosteoblasts (MC3T3-E1) | F-actin/vinculin | Outside–in | 24 | Increased FA size [169] |
| SiO2@IONP | Human bone marrow-derived mesenchymal stem cells (hBM-MSCs) | F-actin/vinculin | Inside–out | 12 | Decreased FA size, decreased phospho-Src, reduced phospho-FAK, reduced traction forces and cell migration [153] |
| Ga-substituted ε-Fe2O3 | A549 | F-actin/vinculin | Inside–out | 24 | Decreased FA area and reduced cell adhesion [145] |
| Resovist, Endorem, and magnetoliposomes | Primary human blood outgrowth endothelial cells (hBOECs) | F-actin/vinculin | Inside–out | 24 | Decreased FA formation and maturation Reduced phospho-FAK Reduced cell migration [160] |
| AuNP (16, 30, 40 nm) | Mouse endothelial cell(SVEC4-10) | F-actin/vinculin and F-actin/zyxin | Outside–in | 2 and 24 | Decreased total FA size, reduced MT1-MMP recruitment Reduced MMP2 and ECM degradation [159] |
| AuNP (16, 30, 40 nm) | Mouse mesenchymal stem cells | F-actin/vinculin and F-actin/zyxin | Outside–in | 2 and 24 | Decreased total and mature FA, reduced ECM degradation [159] |
| AuNP | Human umbilical vein endothelial cells (HUVECs) | F-actin/vinculin | Inside–out | 24 | Reduced FA size and impaired F-actin and tubulin polymerization [155] |
| SiO2 | Oral mucosa cells TR156 | F-actin/vinculin | Inside–out | 12 | Increased F-actin filament traction, increased FA formation Decreased cell migration [161] |
| TiO2 | Oral mucosa cells TR156 | F-actin/vinculin | Inside–out | 12 | Increased F-actin filament traction, increased FA formation Decreased cell migration [161] |
| Hydroxyapatite | Oral mucosa cells TR156 | F-actin/vinculin | Inside–out | 12 | Increased F-actin filament traction, increased FA formation Decreased cell migration [161] |
| TiO2 | Human osteoblast-like cells SaOS-2 | F-actin/p-FAK | Inside–out | 24 | Decreased FA area, reduced cell migration Reduced vinculin [167] |
| Polyethylenimine@IONP | HUVEC | F-actin/vinculin p-cortactin | Inside–out | 6 | Lower phospho-cortactin+ FA, reduced cell migrationReduced phospho-Src and phospho-cortactin ROS involvement [154] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulens-Arias, V. Dissecting the Inorganic Nanoparticle-Driven Interferences on Adhesome Dynamics. J. Nanotheranostics 2021, 2, 174-195. https://doi.org/10.3390/jnt2030011
Mulens-Arias V. Dissecting the Inorganic Nanoparticle-Driven Interferences on Adhesome Dynamics. Journal of Nanotheranostics. 2021; 2(3):174-195. https://doi.org/10.3390/jnt2030011
Chicago/Turabian StyleMulens-Arias, Vladimir. 2021. "Dissecting the Inorganic Nanoparticle-Driven Interferences on Adhesome Dynamics" Journal of Nanotheranostics 2, no. 3: 174-195. https://doi.org/10.3390/jnt2030011
APA StyleMulens-Arias, V. (2021). Dissecting the Inorganic Nanoparticle-Driven Interferences on Adhesome Dynamics. Journal of Nanotheranostics, 2(3), 174-195. https://doi.org/10.3390/jnt2030011