
Nanostructured Silicon Enabled HR-MS for the Label-Free Detection of Biomarkers in Colorectal Cancer Plasma Small Extracellular Vesicles

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Circulating EVs from Plasma Samples
2.2. Transmission Electron Microscopy Phosphotungstic Acid (TEM PTA) Negative Staining Protocol
2.3. Nanoparticle Tracking (NTA) for EVs Size Characterisation
2.4. Extraction of Lipids and Proteins from EVs
2.5. DIOS Fabrication
2.6. NIMS Fabrication
2.7. FT-ICR-MS Measurement and Sample Deposition
2.8. Data Processing and Statistics
3. Results
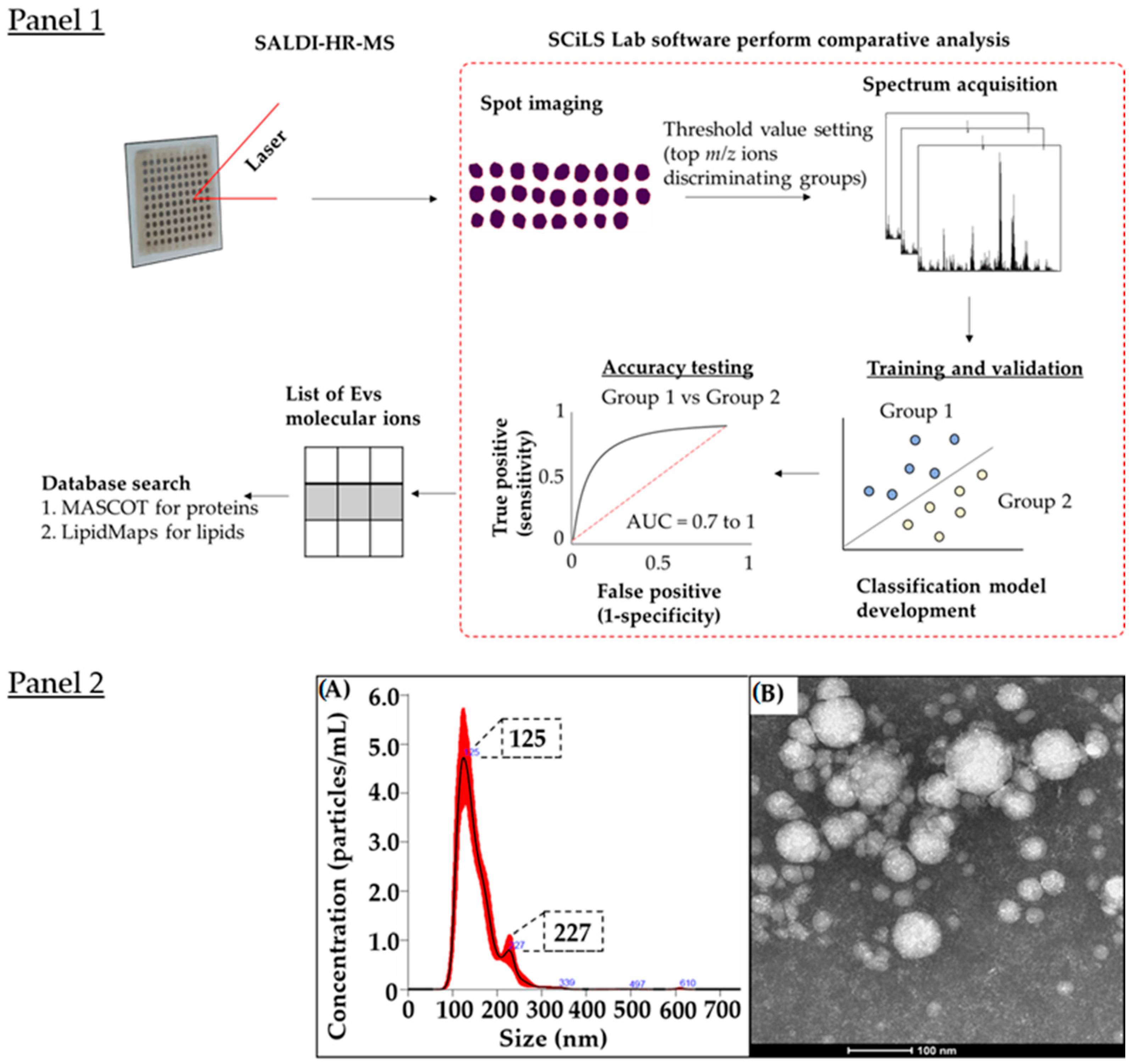
3.1. EVs Characterisation
3.2. SALDI Surfaces for Lipid and Peptide Detection
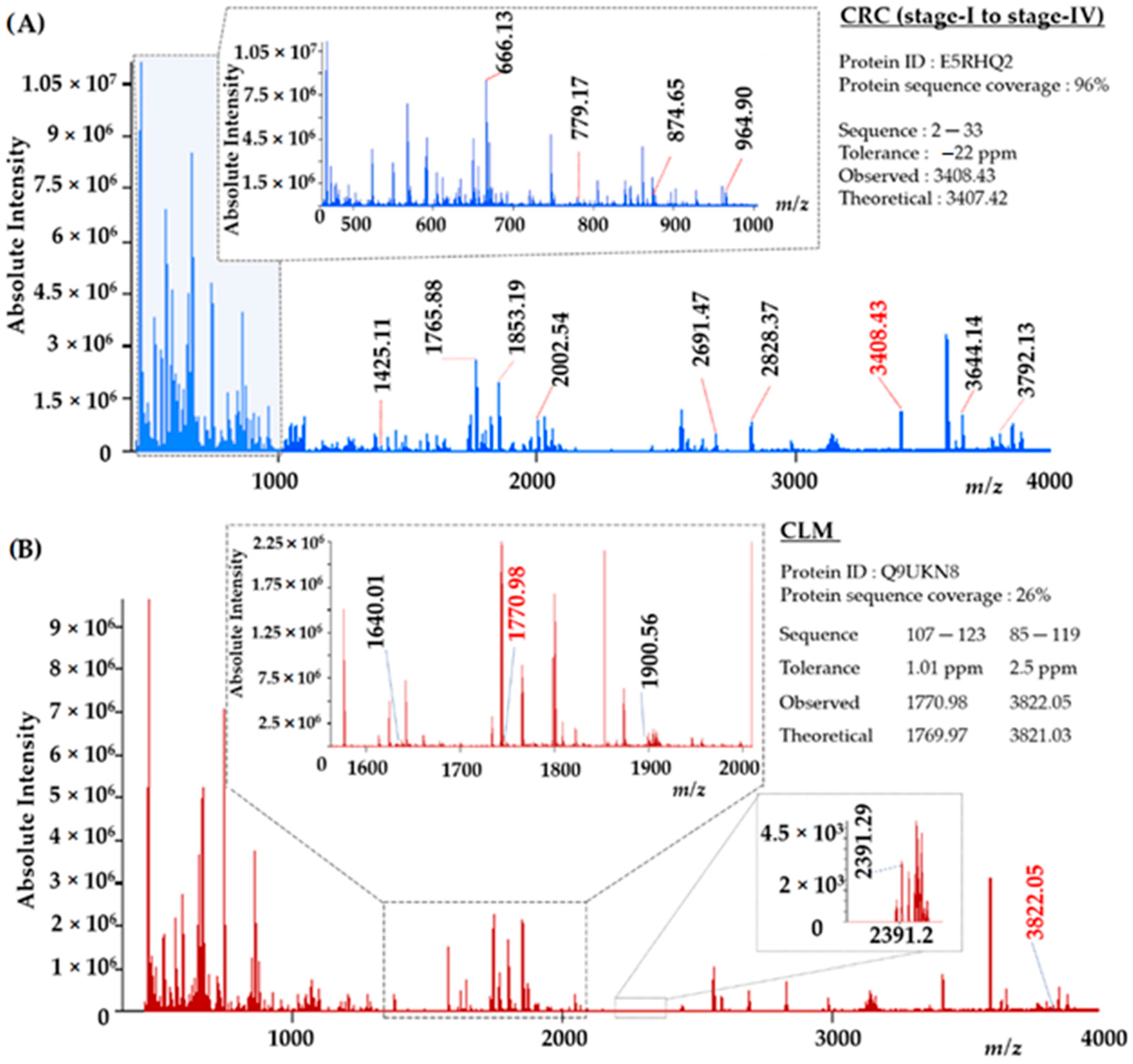
3.3. NIMS-HR-MS for EVs Peptide Mass Fingerprint Identification
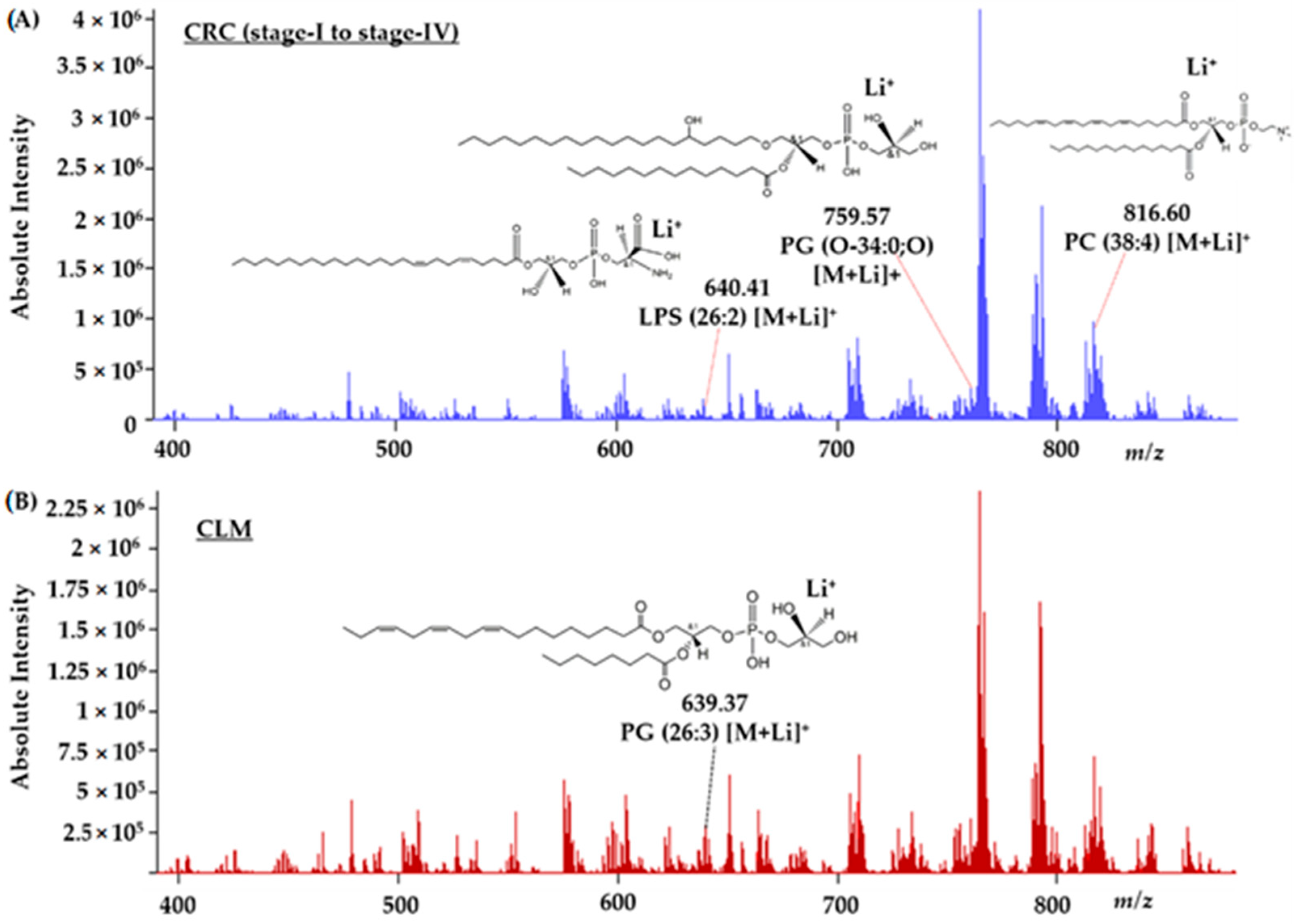
3.4. Interpretation of Lipid Species Associated with Metastatic CRC Pathology Using DIOS-FT-ICR-MS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, H.; Lavotshkin, S.; Lyden, D. The secreted factors responsible for pre-metastatic niche formation: Old sayings and new thoughts. Semin. Cancer Biol. 2011, 21, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Jahn, K.A.; Su, Y.; Braet, F. Multifaceted nature of membrane microdomains in colorectal cancer. World J. Gastroenterol. WJG 2011, 17, 681. [Google Scholar] [CrossRef] [PubMed]
- Gao, F. HER Family Receptor Activation and Dimerisation in Colorectal Cancer and Cancer-Derived Exosomes. Ph.D. Thesis, UCL (University College London), London, UK, December 2019. [Google Scholar]
- Matsumura, T.; Sugimachi, K.; Iinuma, H.; Takahashi, Y.; Kurashige, J.; Sawada, G.; Ueda, M.; Uchi, R.; Ueo, H.; Takano, Y. Exosomal microRNA in serum is a novel biomarker of recurrence in human colorectal cancer. Br. J. Cancer 2015, 113, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Han, B.; Gao, S.; Wang, X.; Wang, Z.; Wang, F.; Zhang, J.; Xu, D.; Sun, B. Exosome-encapsulated microRNAs as circulating biomarkers for colorectal cancer. Oncotarget 2017, 8, 60149. [Google Scholar] [CrossRef] [Green Version]
- Belov, L.; Matic, K.J.; Hallal, S.; Best, O.G.; Mulligan, S.P.; Christopherson, R.I. Extensive surface protein profiles of extracellular vesicles from cancer cells may provide diagnostic signatures from blood samples. J. Extracell. Vesicles 2016, 5, 25355. [Google Scholar] [CrossRef]
- Silva, J.; Garcia, V.; Rodriguez, M.; Compte, M.; Cisneros, E.; Veguillas, P.; Garcia, J.; Dominguez, G.; Campos-Martin, Y.; Cuevas, J. Analysis of exosome release and its prognostic value in human colorectal cancer. Genes Chromosom. Cancer 2012, 51, 409–418. [Google Scholar] [CrossRef]
- Hong, B.S.; Cho, J.-H.; Kim, H.; Choi, E.-J.; Rho, S.; Kim, J.; Kim, J.H.; Choi, D.-S.; Kim, Y.-K.; Hwang, D. Colorectal cancer cell-derived microvesicles are enriched in cell cycle-related mRNAs that promote proliferation of endothelial cells. BMC Genom. 2009, 10, 556. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.-S.; Lee, J.-M.; Park, G.W.; Lim, H.-W.; Bang, J.Y.; Kim, Y.-K.; Kwon, K.-H.; Kwon, H.J.; Kim, K.P.; Gho, Y.S. Proteomic analysis of microvesicles derived from human colorectal cancer cells. J. Proteome Res. 2007, 6, 4646–4655. [Google Scholar] [CrossRef]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef]
- Lydic, T.A.; Townsend, S.; Adda, C.G.; Collins, C.; Mathivanan, S.; Reid, G.E. Rapid and comprehensive ‘shotgun’lipidome profiling of colorectal cancer cell derived exosomes. Methods 2015, 87, 83–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrowska, M.; Wlosowicz, A.; Gawin, M.; Widlak, P. MS-based proteomic analysis of serum and plasma: Problem of high abundant components and lights and shadows of albumin removal. Emerg. Sample Treat. Proteom. 2019, 1073, 57–76. [Google Scholar]
- Tu, C.; Rudnick, P.A.; Martinez, M.Y.; Cheek, K.L.; Stein, S.E.; Slebos, R.J.; Liebler, D.C. Depletion of abundant plasma proteins and limitations of plasma proteomics. J. Proteome Res. 2010, 9, 4982–4991. [Google Scholar] [CrossRef] [Green Version]
- Gundry, R.L.; White, M.Y.; Murray, C.I.; Kane, L.A.; Fu, Q.; Stanley, B.A.; Van Eyk, J.E. Preparation of proteins and peptides for mass spectrometry analysis in a bottom-up proteomics workflow. Curr. Protoc. Mol. Biol. 2010, 90, 10.25.11–10.25.23. [Google Scholar]
- Guinan, T.; Kirkbride, P.; Pigou, P.E.; Ronci, M.; Kobus, H.; Voelcker, N.H. Surface-assisted laser desorption ionization mass spectrometry techniques for application in forensics. Mass Spectrom. Rev. 2015, 34, 627–640. [Google Scholar] [CrossRef]
- Müller, W.H.; Verdin, A.; Kune, C.; Far, J.; De Pauw, E.; Malherbe, C.; Eppe, G. Dual-polarity SALDI FT-ICR MS imaging and Kendrick mass defect data filtering for lipid analysis. Anal. Bioanal. Chem. 2021, 413, 2821–2830. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhao, Y.; Angeles, A.; Hamuro, L.L.; Arnold, M.E.; Shen, J.X. A novel and cost effective method of removing excess albumin from plasma/serum samples and its impacts on LC-MS/MS bioanalysis of therapeutic proteins. Anal. Chem. 2014, 86, 8336–8343. [Google Scholar] [CrossRef]
- Minhas, R.S.; Rudd, D.A.; Al Hmoud, H.Z.; Guinan, T.M.; Kirkbride, K.P.; Voelcker, N.H. Rapid detection of anabolic and narcotic doping agents in saliva and urine by means of nanostructured silicon SALDI mass spectrometry. ACS Appl. Mater. Interfaces 2020, 12, 31195–31204. [Google Scholar] [CrossRef]
- Krismastuti, F.S.H.; Cavallaro, A.; Prieto-Simon, B.; Voelcker, N.H. Toward multiplexing detection of wound healing biomarkers on porous silicon resonant microcavities. Adv. Sci. 2016, 3, 1500383. [Google Scholar] [CrossRef] [Green Version]
- Cerruti, C.D.; Touboul, D.; Guérineau, V.; Petit, V.W.; Laprévote, O.; Brunelle, A. MALDI imaging mass spectrometry of lipids by adding lithium salts to the matrix solution. Anal. Bioanal. Chem. 2011, 401, 75–87. [Google Scholar] [CrossRef]
- Mittal, P.; Price, Z.K.; Lokman, N.A.; Ricciardelli, C.; Oehler, M.K.; Klingler-Hoffmann, M.; Hoffmann, P. Matrix assisted laser desorption/ionization mass spectrometry imaging (MALDI MSI) for monitoring of drug response in primary cancer spheroids. Proteomics 2019, 19, 1900146. [Google Scholar] [CrossRef] [PubMed]
- Guinan, T.; Ronci, M.; Vasani, R.; Kobus, H.; Voelcker, N. Comparison of the performance of different silicon-based SALDI substrates for illicit drug detection. Talanta 2015, 132, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.J.; Woo, H.-K.; Yanes, O.; Shriver, L.; Thomas, D.; Uritboonthai, W.; Apon, J.V.; Steenwyk, R.; Manchester, M.; Siuzdak, G. Detection of carbohydrates and steroids by cation-enhanced nanostructure-initiator mass spectrometry (NIMS) for biofluid analysis and tissue imaging. Anal. Chem. 2010, 82, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, H.-K.; Northen, T.R.; Yanes, O.; Siuzdak, G. Nanostructure-initiator mass spectrometry: A protocol for preparing and applying NIMS surfaces for high-sensitivity mass analysis. Nat. Protoc. 2008, 3, 1341–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, K. Laser desorption/ionization mass spectrometry on nanostructured semiconductor substrates: DIOS™ and QuickMass™. Int. J. Mass Spectrom. 2010, 290, 72–84. [Google Scholar] [CrossRef]
- Guinan, T.; Kirkbride, P.; Della Vedova, C.; Kershaw, S.; Kobus, H.; Voelcker, N. Direct detection of illicit drugs from biological fluids by desorption/ionization mass spectrometry with nanoporous silicon microparticles. Analyst 2015, 140, 7926–7933. [Google Scholar] [CrossRef] [PubMed]
- Li, K.P.; Fang, Y.P.; Liao, J.Q.; Duan, J.D.; Feng, L.G.; Luo, X.Z.; Liang, Z.J. Upregulation of miR-598 promotes cell proliferation and cell cycle progression in human colorectal carcinoma by suppressing INPP5E expression. Mol. Med. Rep. 2018, 17, 2991–2997. [Google Scholar] [CrossRef] [Green Version]
- Eichhorn, P.J.; Creyghton, M.P.; Bernards, R. Protein phosphatase 2A regulatory subunits and cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2009, 1795, 1–15. [Google Scholar] [CrossRef]
- Liang, W.-l.; Cao, J.; Xu, B.; Yang, P.; Shen, F.; Sun, Z.; Li, W.-l.; Wang, Q.; Liu, F. miR-892a regulated PPP2R2A expression and promoted cell proliferation of human colorectal cancer cells. Biomed. Pharmacother. 2015, 72, 119–124. [Google Scholar] [CrossRef]
- Lugini, L.; Valtieri, M.; Federici, C.; Cecchetti, S.; Meschini, S.; Condello, M.; Signore, M.; Fais, S. Exosomes from human colorectal cancer induce a tumor-like behavior in colonic mesenchymal stromal cells. Oncotarget 2016, 7, 50086. [Google Scholar] [CrossRef] [Green Version]
- Saglam, O.; Garrett, C.R.; Boulware, D.; Sayegh, Z.; Shibata, D.; Malafa, M.; Yeatman, T.; Cheng, J.Q.; Sebti, S.; Coppola, D. Activation of the serine/threonine protein kinase AKT during the progression of colorectal neoplasia. Clin. Color. Cancer 2007, 6, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Messaritakis, I.; Stogiannitsi, M.; Koulouridi, A.; Sfakianaki, M.; Voutsina, A.; Sotiriou, A.; Athanasakis, E.; Xynos, E.; Mavroudis, D.; Tzardi, M. Evaluation of the detection of Toll-like receptors (TLRs) in cancer development and progression in patients with colorectal cancer. PLoS ONE 2018, 13, e0197327. [Google Scholar] [CrossRef] [PubMed]
- Hackl, C.; Lang, S.A.; Moser, C.; Mori, A.; Fichtner-Feigl, S.; Hellerbrand, C.; Dietmeier, W.; Schlitt, H.J.; Geissler, E.K.; Stoeltzing, O. Activating transcription factor-3 (ATF3) functions as a tumor suppressor in colon cancer and is up-regulated upon heat-shock protein 90 (Hsp90) inhibition. BMC Cancer 2010, 10, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Ying, L.; Li, X.; Qiao, B.; Meng, Q.; Yu, L.; Yuan, X.; Ren, S.-T.; Chan, D.W.; Shi, L. Overexpression of the transcription factor ATF3 with a regulatory molecular signature associates with the pathogenic development of colorectal cancer. Oncotarget 2017, 8, 47020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Yang, L.; Li, L.; Bai, Y.; Cai, C.; Liu, H. A plasma lipidomics strategy reveals perturbed lipid metabolic pathways and potential lipid biomarkers of human colorectal cancer. J. Chromatogr. B 2017, 1068, 41–48. [Google Scholar] [CrossRef]
- Berdyshev, E.V.; Gorshkova, I.A.; Usatyuk, P.; Zhao, Y.; Saatian, B.; Hubbard, W.; Natarajan, V. De novo biosynthesis of dihydrosphingosine-1-phosphate by sphingosine kinase 1 in mammalian cells. Cell. Signal. 2006, 18, 1779–1792. [Google Scholar] [CrossRef]
- Pyne, N.J.; Pyne, S. Sphingosine 1-phosphate and cancer. Nat. Rev. Cancer 2010, 10, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, P.; Xuan, J.; Zhu, C.; Liu, J.; Shan, L.; Du, Q.; Ren, Y.; Ye, J. Cholesterol enhances colorectal cancer progression via ROS elevation and MAPK signaling pathway activation. Cell. Physiol. Biochem. 2017, 42, 729–742. [Google Scholar] [CrossRef]
- Papadatos-Pastos, D.; Rabbie, R.; Ross, P.; Sarker, D. The role of the PI3K pathway in colorectal cancer. Crit. Rev. Oncol./Hematol. 2015, 94, 18–30. [Google Scholar] [CrossRef]
- Bahrami, A.; Khazaei, M.; Hasanzadeh, M.; ShahidSales, S.; Joudi Mashhad, M.; Farazestanian, M.; Sadeghnia, H.R.; Rezayi, M.; Maftouh, M.; Hassanian, S.M. Therapeutic potential of targeting PI3K/AKT pathway in treatment of colorectal cancer: Rational and progress. J. Cell. Biochem. 2018, 119, 2460–2469. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishnan, S.T.; Rudd, D.; Rahmani, R.; Antunez, E.E.; Minhas, R.S.; Kirana, C.; Maddern, G.J.; Fenix, K.; Hauben, E.; Voelcker, N.H. Nanostructured Silicon Enabled HR-MS for the Label-Free Detection of Biomarkers in Colorectal Cancer Plasma Small Extracellular Vesicles. J. Nanotheranostics 2022, 3, 189-202. https://doi.org/10.3390/jnt3040013
Krishnan ST, Rudd D, Rahmani R, Antunez EE, Minhas RS, Kirana C, Maddern GJ, Fenix K, Hauben E, Voelcker NH. Nanostructured Silicon Enabled HR-MS for the Label-Free Detection of Biomarkers in Colorectal Cancer Plasma Small Extracellular Vesicles. Journal of Nanotheranostics. 2022; 3(4):189-202. https://doi.org/10.3390/jnt3040013
Chicago/Turabian StyleKrishnan, Sanduru Thamarai, David Rudd, Rana Rahmani, E. Eduardo Antunez, Rajpreet Singh Minhas, Chandra Kirana, Guy J. Maddern, Kevin Fenix, Ehud Hauben, and Nicolas H. Voelcker. 2022. "Nanostructured Silicon Enabled HR-MS for the Label-Free Detection of Biomarkers in Colorectal Cancer Plasma Small Extracellular Vesicles" Journal of Nanotheranostics 3, no. 4: 189-202. https://doi.org/10.3390/jnt3040013
APA StyleKrishnan, S. T., Rudd, D., Rahmani, R., Antunez, E. E., Minhas, R. S., Kirana, C., Maddern, G. J., Fenix, K., Hauben, E., & Voelcker, N. H. (2022). Nanostructured Silicon Enabled HR-MS for the Label-Free Detection of Biomarkers in Colorectal Cancer Plasma Small Extracellular Vesicles. Journal of Nanotheranostics, 3(4), 189-202. https://doi.org/10.3390/jnt3040013