
GC-MS Metabolomics and Antifungal Characteristics of Autochthonous Lactobacillus Strains



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Cultivation Conditions
2.2. Extraction of Extracellular Samples
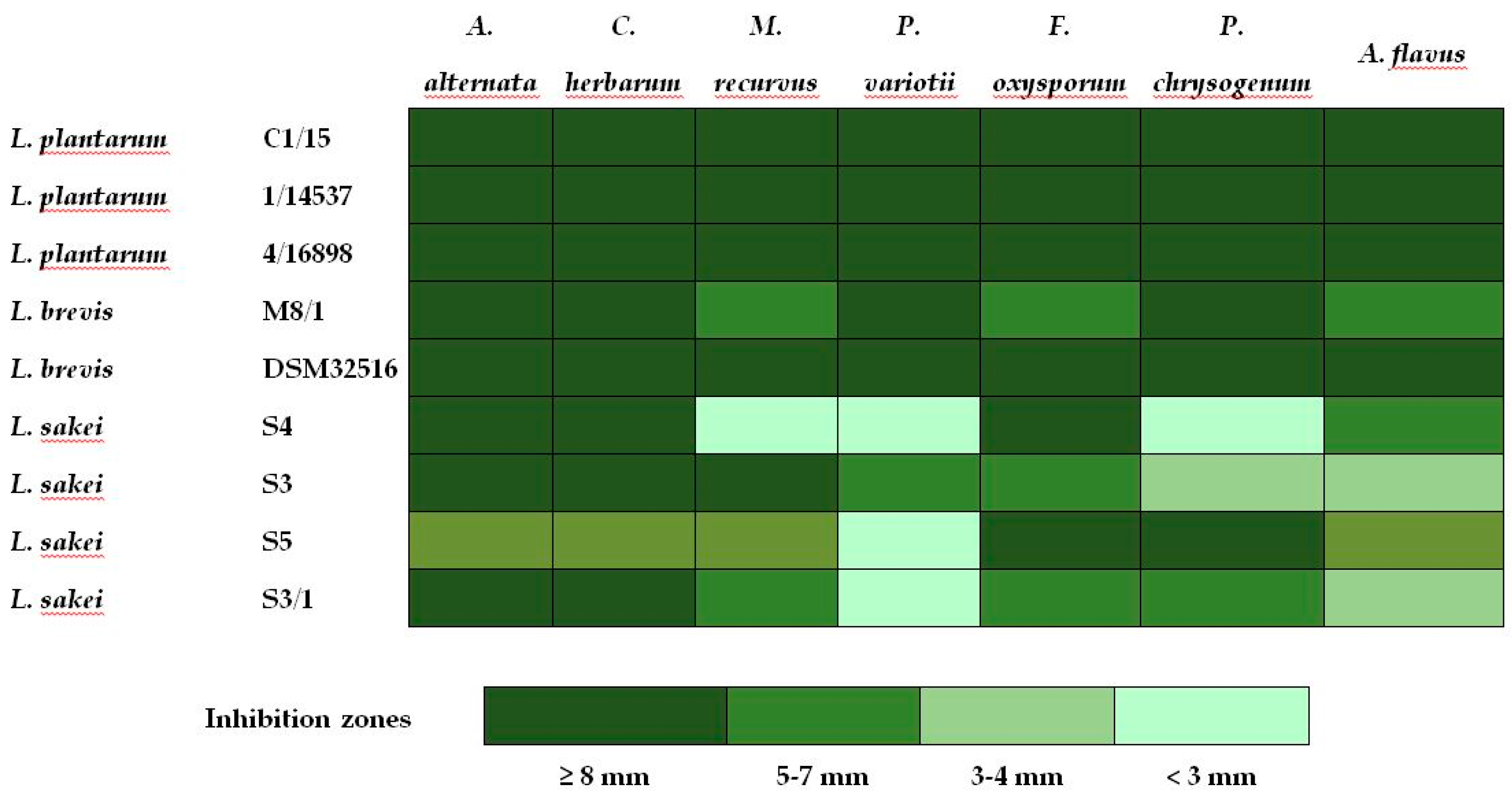
2.3. In Vitro Antifungal Activity of Lactobacillus
2.4. Sample Preparation for GC-MS Analysis
2.5. GC-MS Analysis
2.6. Multivariate Statistical Data Analysis
3. Results
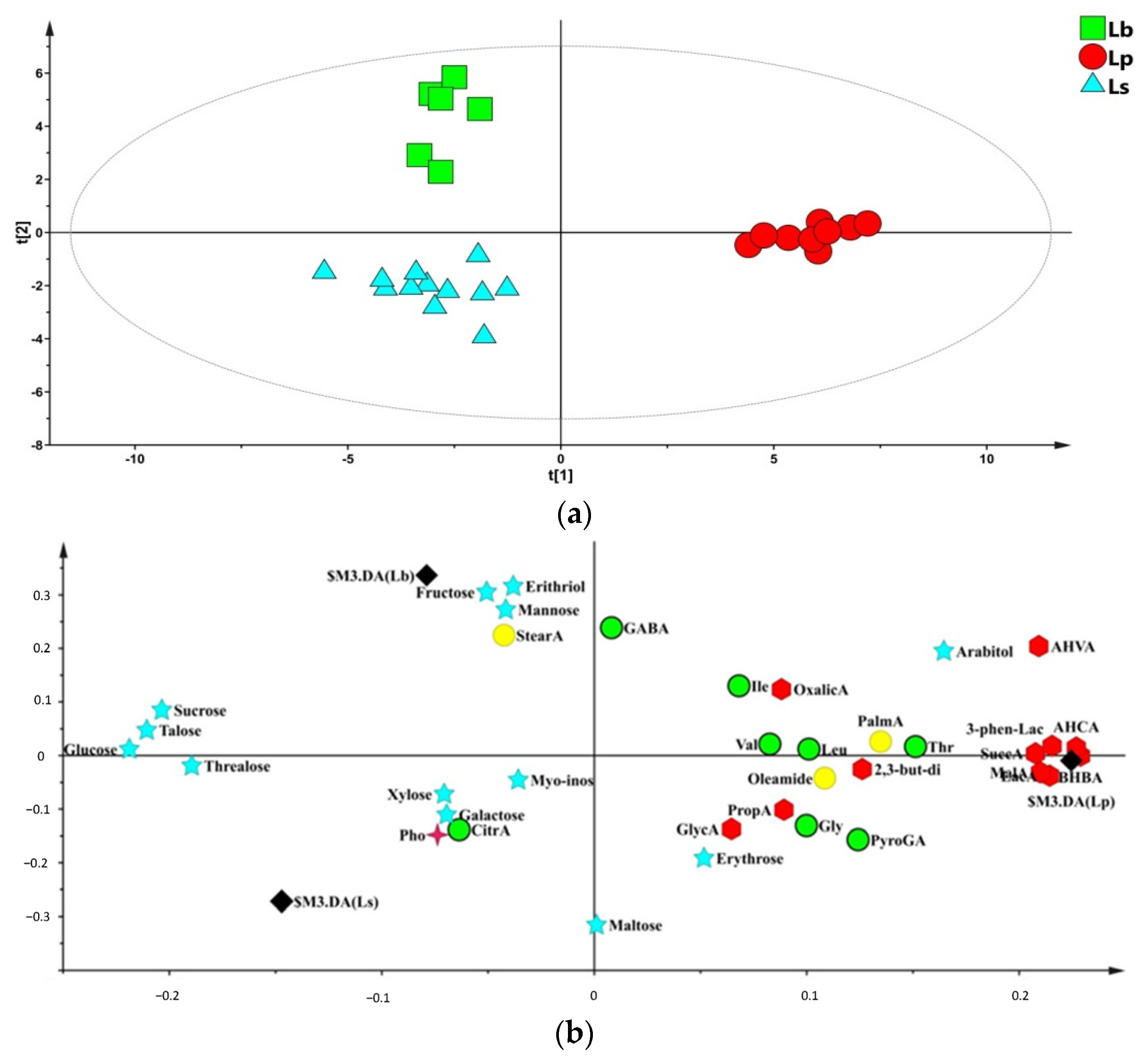
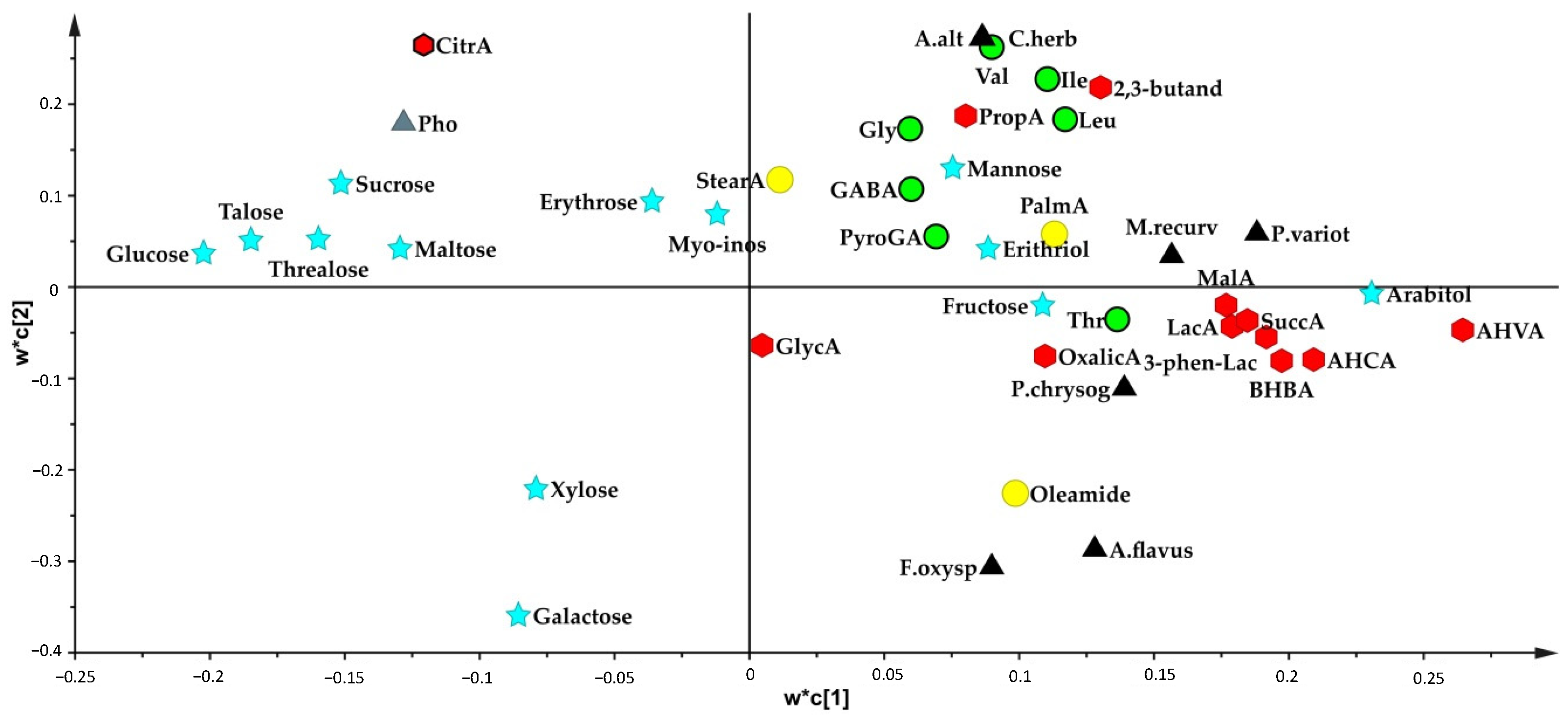
3.1. GC-MS Metabolite Profiles of Lactobacillus Species
3.2. Metabolite Profiles of Lactobacillus Strains
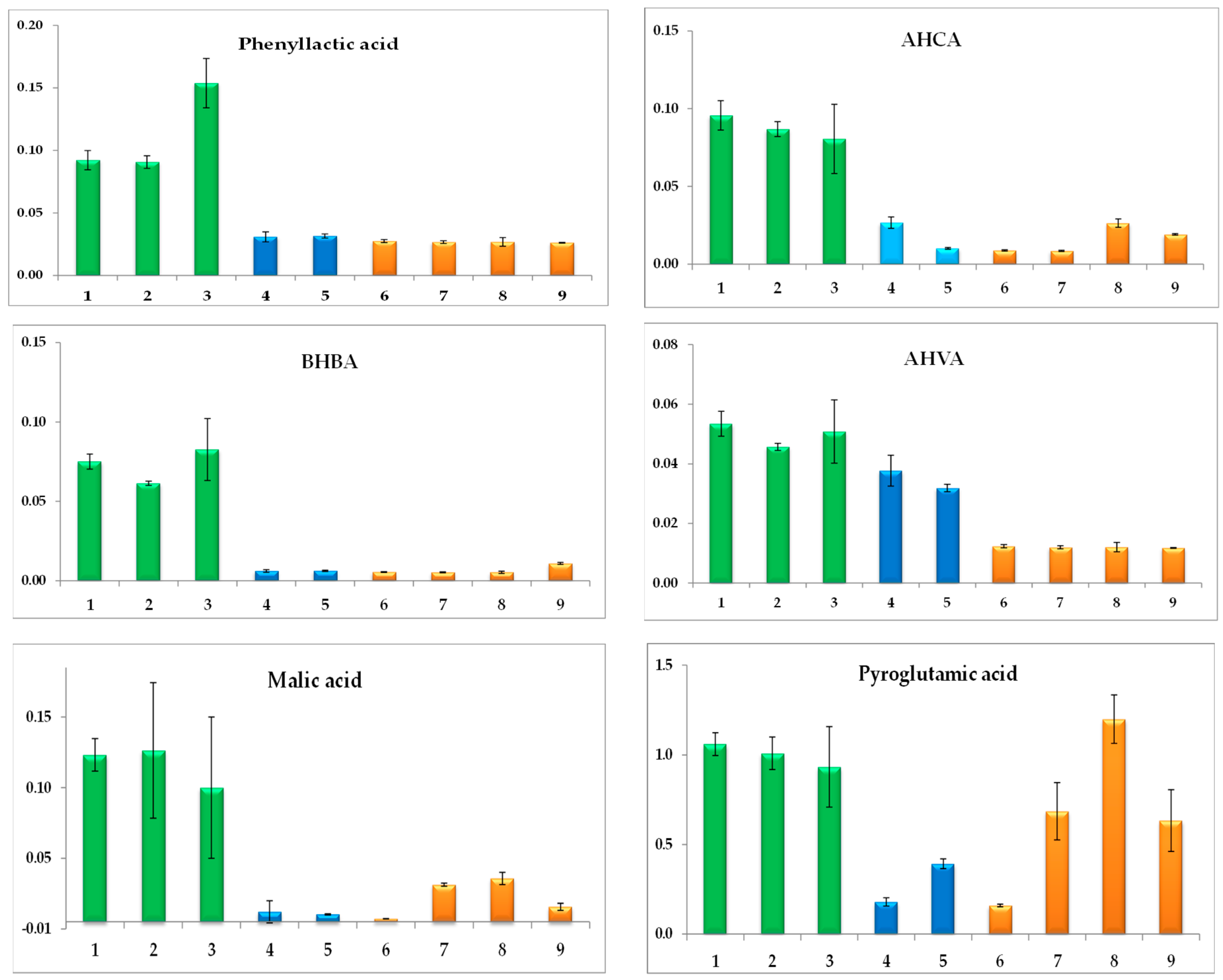
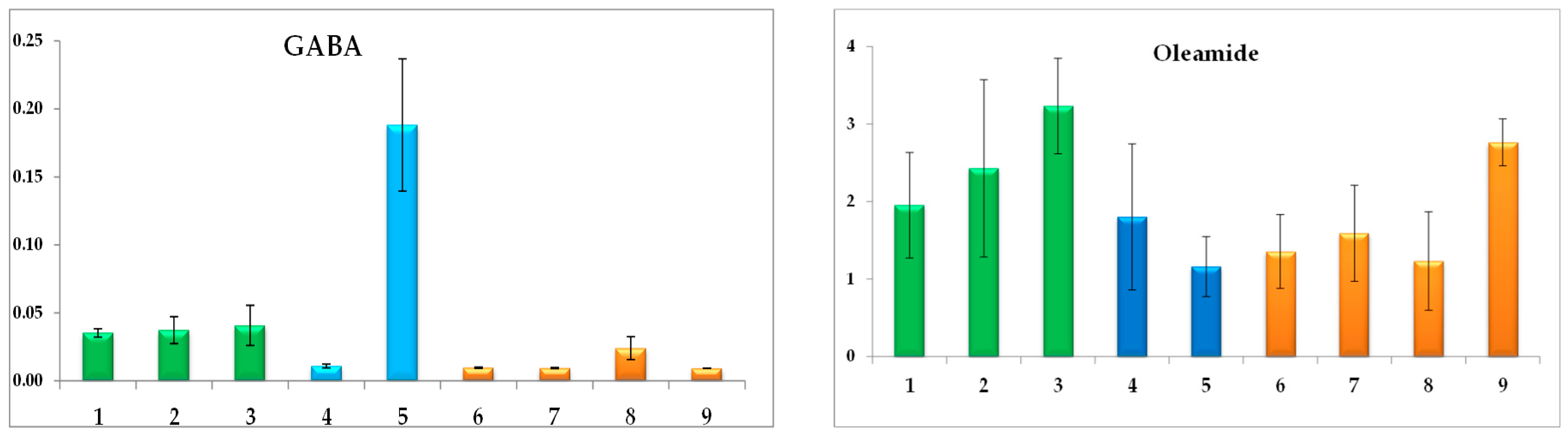
3.3. Antifungal Activity and Correlations with Metabolite Profiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stiles, M.E. Biopreservation by lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 331–345. [Google Scholar] [CrossRef]
- Crowley, S.; Mahony, J.; van Sinderen, D. Current perspectives on antifungal lactic acid bacteria as natural bio-preservatives. Trends Food Sci. Technol. 2013, 33, 93–109. [Google Scholar] [CrossRef]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Cosentino, S.; Viale, S.; Deplano, M.; Fadda, M.E.; Pisano, M.B. Application of autochthonous Lactobacillus strains as biopreservatives to control fungal spoilage in Caciotta cheese. BioMed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champomier-Vergès, M.C.; Chaillou, S.; Cornet, M.; Zagorec, M. Lactobacillus sakei: Recent developments and future prospects. Res. Microbiol. 2001, 152, 839–848. [Google Scholar] [CrossRef]
- Zagorec, M.; Champomier-Vergès, M.C. Lactobacillus sakei: A starter for sausage fermentation, a protective culture for meat products. Microorganisms 2017, 5, 56. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria–Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Reyes-De-Corcuera, J.I.; Etxeberria, E.; Danyluk, M.D.; Rodrick, G.E. Metabolomic analysis in food science: A review. Trends Food Sci. Technol. 2009, 20, 557–566. [Google Scholar] [CrossRef]
- Chaudhary, A.; Verma, K.; Saharan, B.S. A GC-MS Based Metabolic Profiling of Probiotic Lactic Acid Bacteria Isolated from Traditional Food Products. J. Pure Appl. Microbiol. 2020, 14, 657–672. [Google Scholar] [CrossRef] [Green Version]
- Siedler, S.; Balti, R.; Neves, A.R. Bioprotective mechanisms of lactic acid bacteria against fungal spoilage of food. Curr. Opin. Biotechnol. 2019, 56, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Pisano, M.B.; Scano, P.; Murgia, A.; Cosentino, S.; Caboni, P. Metabolomics and microbiological profile of Italian mozzarella cheese produced with buffalo and cow milk. Food Chem. 2016, 192, 618–624. [Google Scholar] [CrossRef]
- Quere, F.; Deschamps AUrdaci, M.C. DNA probe and PCR-specific reaction for Lactobacillus plantarum. J. Appl. Microbiol. 1997, 82, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Young, J.P.W.; Downer, H.L.; Eardly, B.D. Phylogeny of the phototrophic Rhizobium strain BTAil by polymerase chain reaction-based sequencing of a 16S rRNA gene segment. J. Bacteriol. 1991, 173, 2271–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, S.; Cretenet, M.; Vernoux, J.P. Current Knowledge on Antimicrobial Metabolites Produced from Aromatic Amino Acid Metabolism in Fermented Products. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 337–346. [Google Scholar]
- Chaudhari, S.S.; Gokhale, D.V. Phenyllactic acid: A potential antimicrobial compound in lactic acid bacteria. J. Bacteriol. Mycol. Open Access 2016, 2, 00037. [Google Scholar] [CrossRef] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Tajabadi, N.; Baradaran, A.; Ebrahimpour, A.; Rahim, R.A.; Bakar, F.A.; Manap, M.Y.A.; Mohammed, A.S.; Saari, N. Overexpression and optimization of glutamate decarboxylase in Lactobacillus plantarum Taj-Apis 362 for high gamma-aminobutyric acid production. Microbial. Biotechnol. 2015, 8, 623–632. [Google Scholar] [CrossRef]
- Nieminen, M.T.; Novak-Frazer, L.; Rautemaa, V.; Rajendran, R.; Sorsa, T.; Ramage, G. A Novel Antifungal Is Active against Candida albicans Biofilms and Inhibits Mutagenic Acetaldehyde Production In Vitro. PLoS ONE 2014, 9, e97864. [Google Scholar] [CrossRef]
- Sakko, M.; Moore, C.; Novak-Frazer, L.; Rautemaa, V.; Sorsa, T.; Hietala, P.; Järvinen, A.; Bowyer, P.; Tjäderhane, L.; Rautemaa, R. 2-hydroxyisocaproic acid is fungicidal for Candida and Aspergillus species. Mycoses 2014, 57, 214–221. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Donelli, G. Probiotics to counteract biofilm-associated infections: Promising and conflicting data. Int. J. Oral Sci. 2014, 6, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, J.H.; Hwang, S.E.; Han, J.T.; Kim, C.J.; Rho, J.R.; Shin, J.E. Production of oleamide, a functional lipid, by Streptomyces sp. KK90378. J. Microbiol. Biotechnol. 2001, 11, 1018–1023. [Google Scholar]
- Ano, Y.; Nakayama, H. Preventive effects of dairy products on dementia and the underlying mechanisms. Int. J. Mol. Sci. 2018, 19, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cravatt, B.F.; Prospero-Garcia, O.; Siuzdak, G.; Gilula, N.B.; Henriksen, S.J.; Boger, D.L.; Lerner, R.A. Chemical characterization of a family of brain lipids that induce sleep. Science 1995, 268, 1506–1509. [Google Scholar] [CrossRef] [Green Version]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Symposium review: Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, L. Lactic Acid Bacteria: Classification and Physiology. In Lactic acid Bacteria, Microbiological and Functional Aspects; Salminem, S., von Wright, A., Ouwehand, A., Eds.; Marcel Dekker: New York, NY, USA, 2004; Volume 139, pp. 1–66. [Google Scholar]
- Diana, M.; Rafecas, M.; Arco, C.; Quίlez, J. Free amino acid profile of Spanish artisanal cheese: Importance of gamma-aminobutyric acid (GABA) and ornithine content. J. Compos. Anal. 2014, 35, 94–100. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strains | Origin | Molecular Identification | MBDS # |
|---|---|---|---|---|
| Lactobacillus plantarum | 4/16898 | Raw sheep’s milk | Species- specific PCR [12] | UNICAB27 |
| Lactobacillus plantarum | 1/14537 | Raw sheep’s milk | Species- specific PCR [12] | UNICAB32 |
| Lactobacillus plantarum | C1/15 | Sheep’s cheese | Species- specific PCR [12] | UNICAB212 |
| Lactobacillus brevis | DSM 32516 | Sheep’s cheese | 16S rRNA gene sequencing Universal primers F357-R518 [13] | UNICAB24 |
| Lactobacillus brevis | M8/1 | Sheep’s cheese | 16S rRNA gene sequencing Universal primers F357-R518 [13] | UNICAB456 |
| Lactobacillus sakei | S3 | Artisanal sausage | 16S rRNA gene sequencing Universal primers Y1-Y2 [14] | UNICAB457 |
| Lactobacillus sakei | S5 | Artisanal sausage | 16S rRNA gene sequencing Universal primers Y1-Y2 [14] | UNICAB458 |
| Lactobacillus sakei | S4 | Artisanal sausage | 16S rRNA gene sequencing Universal primers F357-R518 [13] | UNICAB459 |
| Lactobacillus sakei | S3/1 | Artisanal sausage | 16S rRNA gene sequencing Universal primers Y1-Y2 [14] | UNICAB460 |
| L. plantarum b | L. brevis | L. sakei | ||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite | Class c | VIP | Metabolite | Class | VIP | Metabolite | Class | VIP |
| 3-hydroxy butyric acid (BHBA) | OA | 1.25 | Fructose | S | 1.66 | Maltose | S | 1.65 |
| 2-hydroxy isovaleric acid (AHVA) | OA | 1.51 | Erithritol | S | 1.64 | Sucrose | S | 1.23 |
| Arabitol | S | 1.32 | 2-hydroxy isovaleric acid (AHVA) | OA | 1.51 | Glucose | S | 1.20 |
| 2-hydroxy isocaproic acid (AHCA) | OA | 1.24 | Mannose | S | 1.45 | Talose | S | 1.20 |
| Lactic Acid | OA | 1.20 | Arabitol | S | 1.32 | Pyroglutamic acid | AA | 1.09 |
| 3-phenyllactic acid | OA | 1.18 | 4-aminobutyric acid (GABA) | AA | 1.24 | Erythrose | S | 1.04 |
| Malic acid | OA | 1.17 | Sucrose | S | 1.23 | Threalose | S | 1.03 |
| Succinic acid | OA | 1.13 | Stearic acid | FA | 1.21 | |||
| Pyroglutamic acid | AA | 1.09 | Talose | S | 1.20 | |||
| Erythrose | S | 1.04 | Threalose | S | 1.03 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scano, P.; Pisano, M.B.; Murgia, A.; Cosentino, S.; Caboni, P. GC-MS Metabolomics and Antifungal Characteristics of Autochthonous Lactobacillus Strains. Dairy 2021, 2, 326-335. https://doi.org/10.3390/dairy2030026
Scano P, Pisano MB, Murgia A, Cosentino S, Caboni P. GC-MS Metabolomics and Antifungal Characteristics of Autochthonous Lactobacillus Strains. Dairy. 2021; 2(3):326-335. https://doi.org/10.3390/dairy2030026
Chicago/Turabian StyleScano, Paola, M. Barbara Pisano, Antonio Murgia, Sofia Cosentino, and Pierluigi Caboni. 2021. "GC-MS Metabolomics and Antifungal Characteristics of Autochthonous Lactobacillus Strains" Dairy 2, no. 3: 326-335. https://doi.org/10.3390/dairy2030026
APA StyleScano, P., Pisano, M. B., Murgia, A., Cosentino, S., & Caboni, P. (2021). GC-MS Metabolomics and Antifungal Characteristics of Autochthonous Lactobacillus Strains. Dairy, 2(3), 326-335. https://doi.org/10.3390/dairy2030026