
Insights into Androgen Receptor Action in Lung Cancer

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
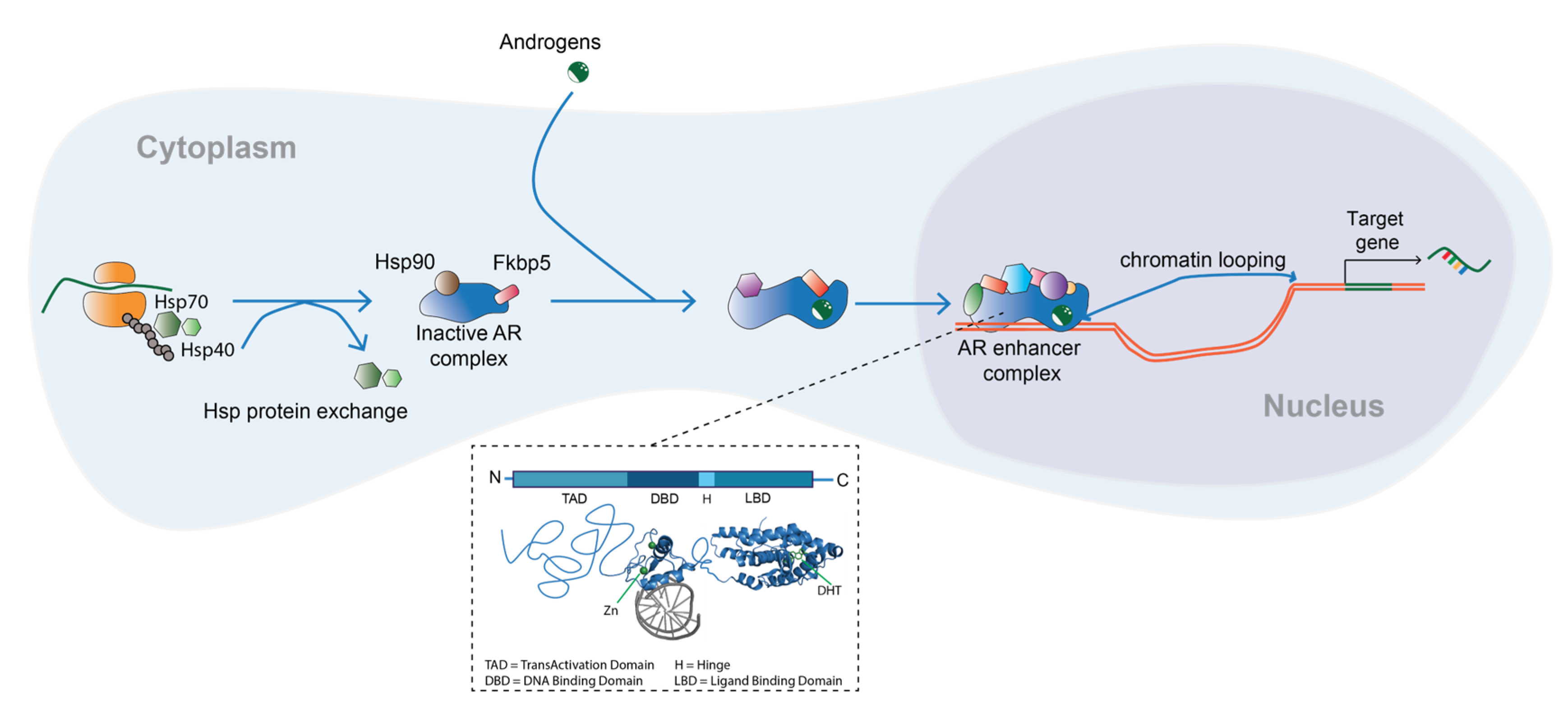
2. Androgens and the Androgen Receptor
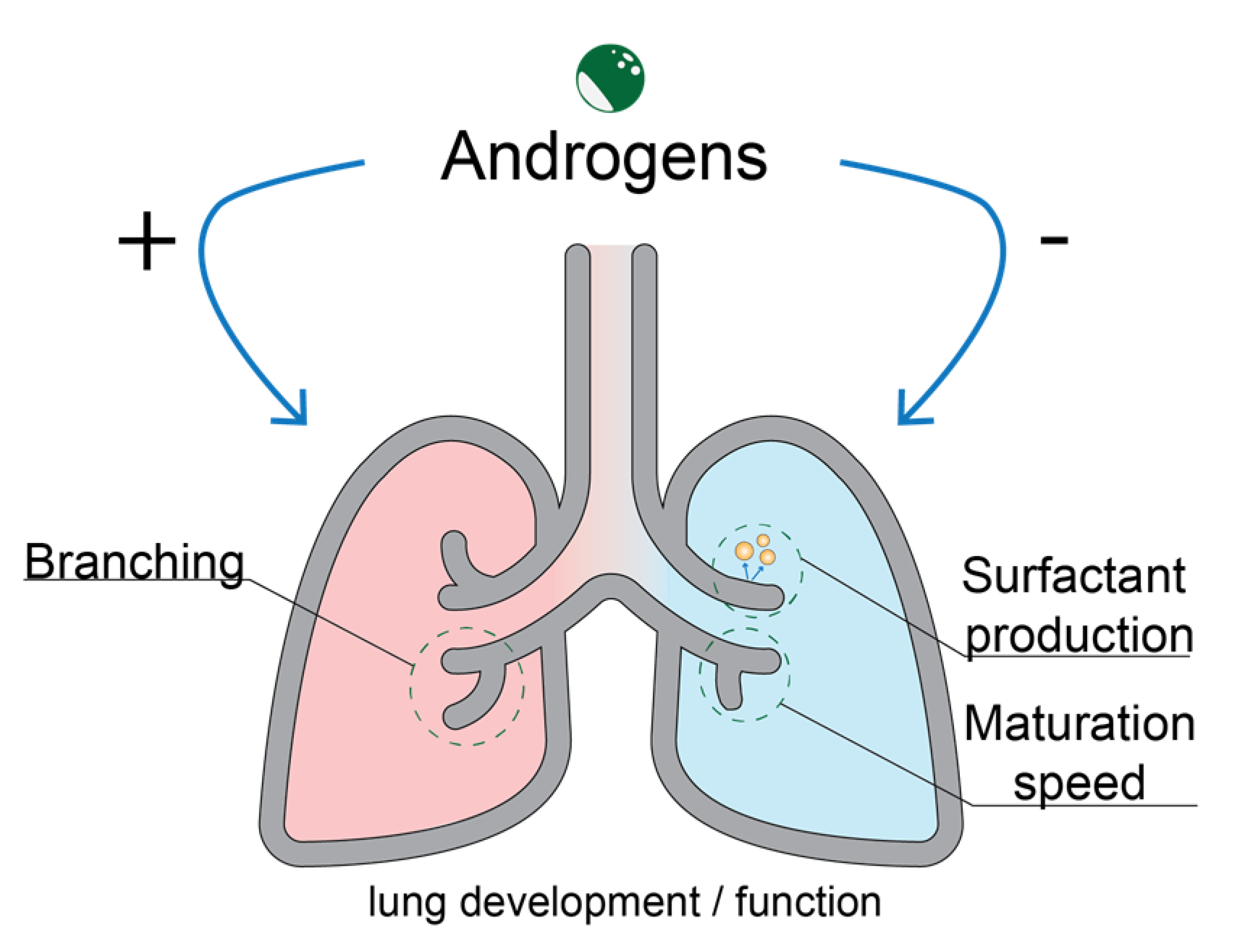
3. Role of the Androgen Receptor in Lung Development
4. Androgens and the Androgen Receptor in Lung Cancer
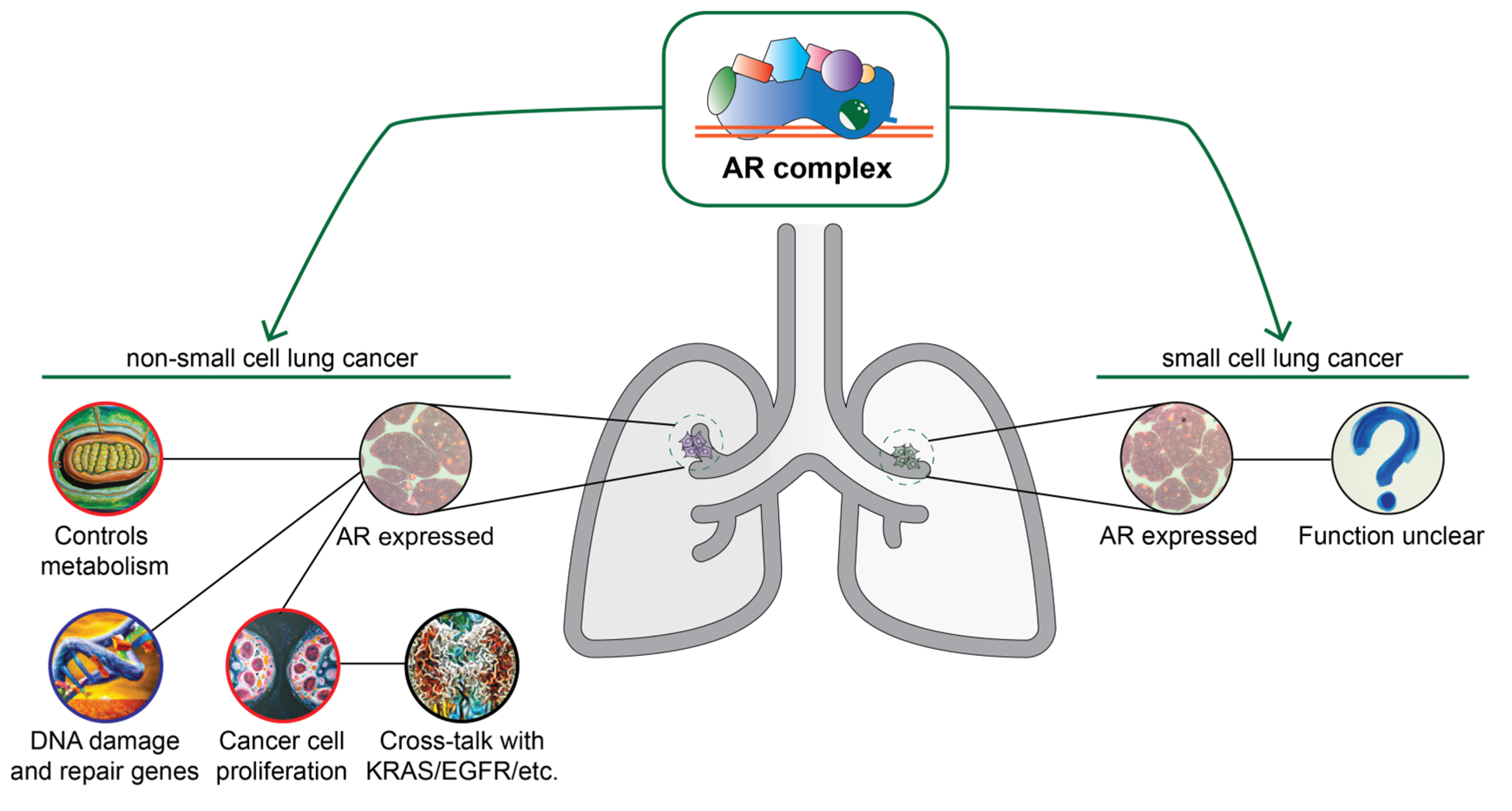
4.1. Androgen Receptor Signaling in Lung Cancer
4.2. Androgen Receptor as an Important Factor in Lung Cancer Biology: Evidence from Clinical Studies
4.3. Targeting of the Androgen Receptor Pathway
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA A Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Bade, B.C.; Dela Cruz, C.S. Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clin. Chest. Med. 2020, 41, 1–24. [Google Scholar] [CrossRef]
- Rades, D.; Setter, C.; Dahl, O.; Schild, S.E.; Noack, F. The prognostic impact of tumor cell expression of estrogen receptor-alpha, progesterone receptor, and androgen receptor in patients irradiated for nonsmall cell lung cancer. Cancer 2012, 118, 157–163. [Google Scholar] [CrossRef]
- Rodak, O.; Peris-Díaz, M.D.; Olbromski, M.; Podhorska-Okołów, M.; Dzięgiel, P. Current landscape of non-small cell lung cancer: Epidemiology, histological classification, targeted therapies, and immunotherapy. Cancers 2021, 13, 4705. [Google Scholar] [CrossRef]
- Recchia, A.G.; Musti, A.M.; Lanzino, M.; Panno, M.L.; Turano, E.; Zumpano, R.; Belfiore, A.; Ando, S.; Maggiolini, M. A cross-talk between the androgen receptor and the epidermal growth factor receptor leads to p38MAPK-dependent activation of mTOR and cyclinD1 expression in prostate and lung cancer cells. Int. J. Biochem. Cell Biol. 2009, 41, 603–614. [Google Scholar] [CrossRef]
- van Meerbeeck, J.P.; Fennell, D.A.; De Ruysscher, D.K.M. Small-cell lung cancer. Lancet 2011, 378, 1741–1755. [Google Scholar] [CrossRef]
- Hsu, L.H.; Chu, N.M.; Kao, S.H. Estrogen, Estrogen Receptor and Lung Cancer. Int. J. Mol. Sci. 2017, 18, 1713. [Google Scholar] [CrossRef] [PubMed]
- Kohno, T.; Nakaoku, T.; Tsuta, K.; Tsuchihara, K.; Matsumoto, S.; Yoh, K.; Goto, K. Beyond ALK-RET, ROS1 and other oncogene fusions in lung cancer. Transl. Lung Cancer Res. 2015, 4, 156. [Google Scholar]
- Dou, M.; Zhu, K.; Fan, Z.; Zhang, Y.; Chen, X.; Zhou, X.; Ding, X.; Li, L.; Gu, Z.; Guo, M.; et al. Reproductive Hormones and Their Receptors May Affect Lung Cancer. Cell Physiol. Biochem. 2017, 44, 1425–1434. [Google Scholar] [CrossRef]
- Carey, M.; Card, J.; Voltz, J.; Germolec, D.; Korach, K.; Zeldin, D. The impact of sex and sex hormones on lung physiology and disease: Lessons from animal studies. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 293, L272–L278. [Google Scholar] [CrossRef] [Green Version]
- Grant, L.; Banerji, S.; Murphy, L.; Dawe, D.; Harlos, C.; Myal, Y.; Nugent, Z.; Blanchard, A.; Penner, C.; Qing, G.; et al. Androgen Receptor and Ki67 Expression and Survival Outcomes in Non-small Cell Lung Cancer. Horm. Cancer 2018, 9, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Harlos, C.; Musto, G.; Lambert, P.; Ahmed, R.; Pitz, M.W. Androgen pathway manipulation and survival in patients with lung cancer. Horm. Cancer 2015, 6, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Hwang, N.M.; Stabile, L.P. Estrogen Receptor ß in Cancer: To ß (e) or not to ß (e)? Endocrinology 2021, 162, bqab162. [Google Scholar] [CrossRef]
- Rodriguez-Lara, V.; Avila-Costa, M.R. An overview of lung cancer in women and the impact of estrogen in lung carcinogenesis and lung cancer treatment. Front. Med. 2021, 8, 600121. [Google Scholar] [CrossRef] [PubMed]
- Stabile, L.P.; Siegfried, J.M. Estrogen Receptor Pathways in Lung Cancer. Curr. Oncol. Rep. 2004, 6, 259–267. [Google Scholar] [CrossRef]
- Azzouni, F.; Godoy, A.; Li, Y.; Mohler, J. The 5 alpha-reductase isozyme family: A review of basic biology and their role in human diseases. Adv. Urol. 2012, 2012, 530121. [Google Scholar] [CrossRef] [Green Version]
- Hyde, Z.; Flicker, L.; McCaul, K.A.; Almeida, O.P.; Hankey, G.J.; Chubb, S.A.; Yeap, B.B. Associations between testosterone levels and incident prostate, lung, and colorectal cancer. A population-based study. Cancer Epidemiol Biomark. Prev 2012, 21, 1319–1329. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.X.; Yeap, B.B. Dihydrotestosterone and cancer risk. Curr. Opin. Endocrinol Diabetes Obes. 2018, 25, 209–217. [Google Scholar] [CrossRef]
- Godoy, A.; Kawinski, E.; Li, Y.; Oka, D.; Alexiev, B.; Azzouni, F.; Titus, M.A.; Mohler, J.L. 5alpha-reductase type 3 expression in human benign and malignant tissues: A comparative analysis during prostate cancer progression. Prostate 2011, 71, 1033–1046. [Google Scholar] [CrossRef] [Green Version]
- Kulik, M.; Bothe, M.; Kibar, G.; Fuchs, A.; Schöne, S.; Prekovic, S.; Mayayo-Peralta, I.; Chung, H.-R.; Zwart, W.; Helsen, C. Androgen and glucocorticoid receptor direct distinct transcriptional programs by receptor-specific and shared DNA binding sites. Nucleic Acids Res. 2021, 49, 3856–3875. [Google Scholar] [CrossRef] [PubMed]
- Rahman, F.; Christian, H.C. Non-classical actions of testosterone: An update. Trends Endocrinol. Metab. 2007, 18, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Davey, R.A.; Grossmann, M. Androgen receptor structure, function and biology: From bench to bedside. Clin. Biochem. Rev. 2016, 37, 3. [Google Scholar] [PubMed]
- Wadosky, K.M.; Koochekpour, S. Androgen receptor splice variants and prostate cancer: From bench to bedside. Oncotarget 2017, 8, 18550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wach, S.; Taubert, H.; Cronauer, M. Role of androgen receptor splice variants, their clinical relevance and treatment options. World J. Urol. 2020, 38, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Prekovic, S.; Van den Broeck, T.; Moris, L.; Smeets, E.; Claessens, F.; Joniau, S.; Helsen, C.; Attard, G. Treatment-induced changes in the androgen receptor axis: Liquid biopsies as diagnostic/prognostic tools for prostate cancer. Mol. Cell Endocrinol. 2018, 462, 56–63. [Google Scholar] [CrossRef]
- Hu, C.; Fang, D.; Xu, H.; Wang, Q.; Xia, H. The androgen receptor expression and association with patient’s survival in different cancers. Genomics 2020, 112, 1926–1940. [Google Scholar] [CrossRef]
- Chang, C.; Lee, S.O.; Wang, R.-S.; Yeh, S.; Chang, T.-M. Androgen receptor (AR) physiological roles in male and female reproductive systems: Lessons learned from AR-knockout mice lacking AR in selective cells. Biol. Reprod. 2013, 89, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Shukla, G.; Plaga, A.; Shankar, E.; Gupta, S. Androgen receptor-related diseases: What do we know? Andrology 2016, 4, 366–381. [Google Scholar] [CrossRef] [Green Version]
- Breza, M.; Koutsis, G. Kennedy’s disease (spinal and bulbar muscular atrophy): A clinically oriented review of a rare disease. J. Neurol. 2019, 266, 565–573. [Google Scholar] [CrossRef]
- Chang, C.; Lee, S.O.; Yeh, S.; Chang, T.M. Androgen receptor (AR) differential roles in hormone-related tumors including prostate, bladder, kidney, lung, breast and liver. Oncogene 2014, 33, 3225–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; Xie, Y.; Lee, W.; Bookout, A.L.; Girard, L.; Raso, G.; Behrens, C.; Wistuba, I.I.; Gadzar, A.F.; Minna, J.D.; et al. Research resource: Diagnostic and therapeutic potential of nuclear receptor expression in lung cancer. Mol. Endocrinol 2012, 26, 1443–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seaborn, T.; Simard, M.; Provost, P.R.; Piedboeuf, B.; Tremblay, Y. Sex hormone metabolism in lung development and maturation. Trends Endocrinol. Metab. 2010, 21, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Boucher, E.; Provost, P.R.; Plante, J.; Tremblay, Y. Androgen receptor and 17beta-HSD type 2 regulation in neonatal mouse lung development. Mol. Cell Endocrinol. 2009, 311, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Patrone, C.; Cassel, T.N.; Pettersson, K.; Piao, Y.S.; Cheng, G.; Ciana, P.; Maggi, A.; Warner, M.; Gustafsson, J.A.; Nord, M. Regulation of postnatal lung development and homeostasis by estrogen receptor beta. Mol. Cell Biol. 2003, 23, 8542–8552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, H. Testosterone Regulation of Sex Differences in Fetal Lung Development. Exp. Biol. Med. 1992, 199, 446–452. [Google Scholar] [CrossRef]
- Kimura, Y.; Suzuki, T.; Kaneko, C.; Darnel, A.D.; Akahira, J.; Ebina, M.; Nukiwa, T.; Sasano, H. Expression of androgen receptor and 5α-reductase types 1 and 2 in early gestation fetal lung: A possible correlation with branching morphogenesis. Clin. Sci. 2003, 105, 709–713. [Google Scholar] [CrossRef]
- Townsend, E.A.; Miller, V.M.; Prakash, Y.S. Sex Differences and Sex Steroids in Lung Health and Disease. Endocr. Rev. 2012, 33, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Provost, P.; Blomquist, C.; Godin, C.; Huang, X.; Flamand, N.; Luu-The, V.; Nadeau, D.; Tremblay, Y. Androgen Formation and Metabolism in the Pulmonary Epithelial Cell Line A549: Expression of 17β-Hydroxysteroid Dehydrogenase Type 5 and 3α—Hydroxysteroid Dehydrogenase Type 3. Endocrinology 2000, 141, 2786–2794. [Google Scholar] [CrossRef]
- Maasberg, M.; Rotsch, M.; Jaques, G.; Enderle-Schmidt, U.; Weehle, R.; Havemann, K. Androgen Receptors, Androgen-Dependent Proliferation, And 5α-Reductase Activity of Small-Cell Lung Cancer Cell Lines. Int. J. Cancer 1989, 43, 685–691. [Google Scholar] [CrossRef]
- Chan, Y.X.; Alfonso, H.; Chubb, S.A.; Handelsman, D.J.; Fegan, P.G.; Hankey, G.J.; Golledge, J.; Flicker, L.; Yeap, B.B. Higher Dihydrotestosterone Is Associated with the Incidence of Lung Cancer in Older Men. Horm. Cancer 2017, 8, 119–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Beyer, H.; Brennan, S.; Stiles, S.; Wiese, D.; Buehler, D.; Saeed, A.; Baschnagel, A.M.; Iyer, G. Abstract 3946: Androgen receptor drives differential gene expression in KRAS-mediated non-small cell lung cancer. Cancer Res. 2018, 78, 3946. [Google Scholar] [CrossRef]
- Tian, T.; Hong, F.; Wang, Z.; Hu, J.; Chen, N.; Lv, L.; Yi, Q. HSD17B6 downregulation predicts poor prognosis and drives tumor progression via activating Akt signaling pathway in lung adenocarcinoma. Cell Death Discov. 2021, 7, 341. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Zhao, Y.; Wei, Q.; Zhao, Y.; Yi, Q. Downexpression of HSD17B6 correlates with clinical prognosis and tumor immune infiltrates in hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 210. [Google Scholar] [CrossRef]
- Auboeuf, D.; Batsché, E.; Dutertre, M.; Muchardt, C.; O’Malley, B.W. Coregulators: Transducing signal from transcription to alternative splicing. Trends Endocrinol. Metab. 2007, 18, 122–129. [Google Scholar] [CrossRef]
- Orsted, D.D.; Nordestgaard, B.G.; Bojesen, S.E. Plasma testosterone in the general population, cancer prognosis and cancer risk: A prospective cohort study. Ann. Oncol. 2014, 25, 712–718. [Google Scholar] [CrossRef]
- Mikkonen, L.; Pihlajamaa, P.; Sahu, B.; Zhang, F.P.; Janne, O.A. Androgen receptor and androgen-dependent gene expression in lung. Mol. Cell Endocrinol. 2010, 317, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Jancik, S.; Drabek, J.; Radzioch, D.; Hajduch, M. Clinical relevance of KRAS in human cancers. J. Biomed Biotechnol. 2010, 2010, 150960. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Yeh, S.; Chong, Y.; Tsai, Y.; Chang, C.; Wu, C. Abstract 2126: Androgen receptor regulates lung cancer progress through modulation of OCT-4 expression. Cell Mol. Biol. 2011, 71, 963–974. [Google Scholar] [CrossRef]
- Yeh, S.-D.; Yang, P.-C.; Lu, H.-H.; Chang, C.; Wu, C.-W. Targeting androgen receptor as a new potential therapeutic approach to battle tobacco carcinogens-induced non-small cell lung cancer. J. Transl. Med. 2012, 10, A8. [Google Scholar] [CrossRef] [Green Version]
- Noronha, R.; Goodall, C. Enhancement by testosterone of dimethylnitrosamine carcinogenesis in lung, liver and kidney of inbred NZR/Gd female rats. Carcinogenesis 1989, 4, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Yoshida, A.; Yoshida, M.; Takahashi, M.; Tsuta, K. Evaluation of androgen receptor and GATA binding protein 3 as immunohistochemical markers in the diagnosis of metastatic breast carcinoma to the lung. Pathol. Int. 2015, 65, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Berardi, R.; Morgese, F.; Santinelli, A.; Onofri, A.; Biscotti, T.; Brunelli, A.; Caramanti, M.; Savini, A.; De Lisa, M.; Ballatore, Z.; et al. Hormonal receptors in lung adenocarcinoma: Expression and difference in outcome by sex. Oncotarget 2016, 7, 82648–82657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skjefstad, K.; Grindstad, T.; Khanehkenari, M.R.; Richardsen, E.; Donnem, T.; Kilvaer, T.; Andersen, S.; Bremnes, R.M.; Busund, L.T.; Al-Saad, S. Prognostic relevance of estrogen receptor alpha, beta and aromatase expression in non-small cell lung cancer. Steroids 2016, 113, 5–13. [Google Scholar] [CrossRef]
- Tomeva, E.; Switzeny, O.J.; Heitzinger, C.; Hippe, B.; Haslberger, A.G. Comprehensive Approach to Distinguish Patients with Solid Tumors from Healthy Controls by Combining Androgen Receptor Mutation p. H875Y with Cell-Free DNA Methylation and Circulating miRNAs. Cancers 2022, 14, 462. [Google Scholar] [CrossRef]
- Helsen, C.; Van den Broeck, T.; Voet, A.; Prekovic, S.; Van Poppel, H.; Joniau, S.; Claessens, F. Androgen receptor antagonists for prostate cancer therapy. Endocr.-Relat. Cancer 2014, 21, T105–T118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, S.; van der Poel, H.G.; Bergman, A.M.; Zwart, W.; Prekovic, S. Enzalutamide therapy for advanced prostate cancer: Efficacy, resistance and beyond. Endocr.-Relat. Cancer 2019, 26, R31–R52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, H.; Sun, Q.; Liu, X.; Wu, Z.; Wang, X.; Fang, W.; Ma, Z. miR-224-5p-enriched exosomes promote tumorigenesis by directly targeting androgen receptor in non-small cell lung cancer. Mol. Nucleic Acids 2021, 23, 1217–1228. [Google Scholar] [CrossRef]
- Gockel, L.M.; Pfeifer, V.; Baltes, F.; Bachmaier, R.D.; Wagner, K.G.; Bendas, G.; Gütschow, M.; Sosič, I.; Steinebach, C. Design, synthesis, and characterization of PROTACs targeting the androgen receptor in prostate and lung cancer models. Arch. Der Pharm. 2022, 355, 2100467. [Google Scholar] [CrossRef]
- English, K.; Pugh, P.; Parry, H.; Scutt, N.; Channer, K.; Jones, T. Effect of cigarette smoking on levels of bioavailable testosterone in healthy men. Clin. Sci. 2001, 100, 661–665. [Google Scholar] [CrossRef]
- Slowikowski, B.K.; Lianeri, M.; Jagodzinski, P.P. Exploring estrogenic activity in lung cancer. Mol. Biol. Rep. 2017, 44, 35–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, A.G.; Prysak, G.M.; Murphy, V.; Lonardo, F.; Pass, H.; Schwartz, J.; Brooks, S. Nuclear estrogen receptor beta in lung cancer: Expression and survival differences by sex. Clin. Cancer. Res. 2005, 11, 7280–7287. [Google Scholar] [CrossRef] [Green Version]
- Olivo-Marston, S.E.; Mechanic, L.E.; Mollerup, S.; Bowman, E.D.; Remaley, A.T.; Forman, M.R.; Skaug, V.; Zheng, Y.L.; Haugen, A.; Harris, C.C. Serum estrogen and tumor-positive estrogen receptor-alpha are strong prognostic classifiers of non-small-cell lung cancer survival in both men and women. Carcinogenesis 2010, 31, 1778–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diver, M.J.; Imtiaz, K.E.; Ahmad, A.M.; Vora, J.P.; Fraser, W.D. Diurnal rhythms of serum total, free and bioavailable testosterone and of SHBG in middle-aged men compared with those in young men. Clin. Endocrinol. 2003, 58, 710–717. [Google Scholar] [CrossRef]
- Brambilla, D.J.; Matsumoto, A.M.; Araujo, A.B.; McKinlay, J.B. The effect of diurnal variation on clinical measurement of serum testosterone and other sex hormone levels in men. J. Clin. Endocrinol. Metab. 2009, 94, 907–913. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durovski, D.; Jankovic, M.; Prekovic, S. Insights into Androgen Receptor Action in Lung Cancer. Endocrines 2023, 4, 269-280. https://doi.org/10.3390/endocrines4020022
Durovski D, Jankovic M, Prekovic S. Insights into Androgen Receptor Action in Lung Cancer. Endocrines. 2023; 4(2):269-280. https://doi.org/10.3390/endocrines4020022
Chicago/Turabian StyleDurovski, Darko, Milica Jankovic, and Stefan Prekovic. 2023. "Insights into Androgen Receptor Action in Lung Cancer" Endocrines 4, no. 2: 269-280. https://doi.org/10.3390/endocrines4020022
APA StyleDurovski, D., Jankovic, M., & Prekovic, S. (2023). Insights into Androgen Receptor Action in Lung Cancer. Endocrines, 4(2), 269-280. https://doi.org/10.3390/endocrines4020022