
Detecting Square Grid Structure in an Animal Neuronal Network

Abstract
:1. Introduction
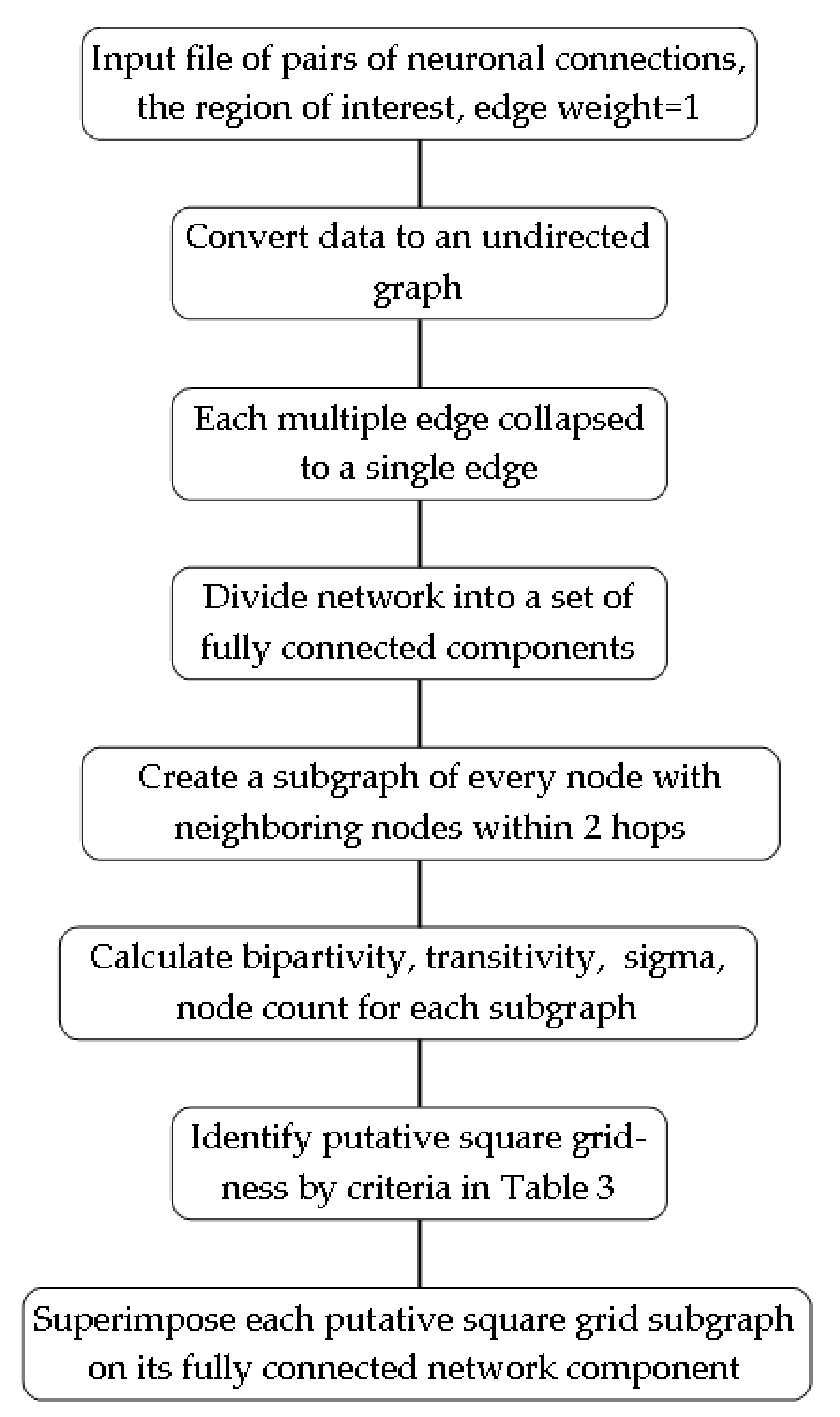
2. Methods
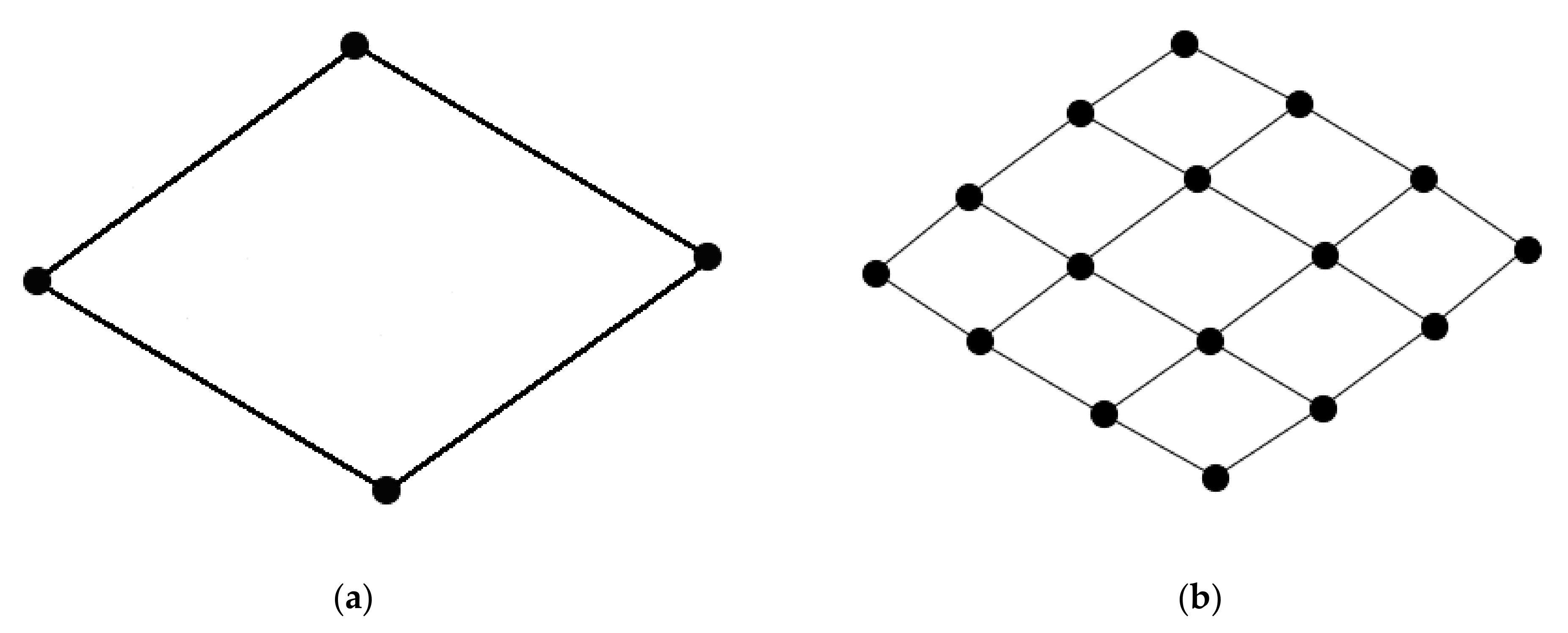
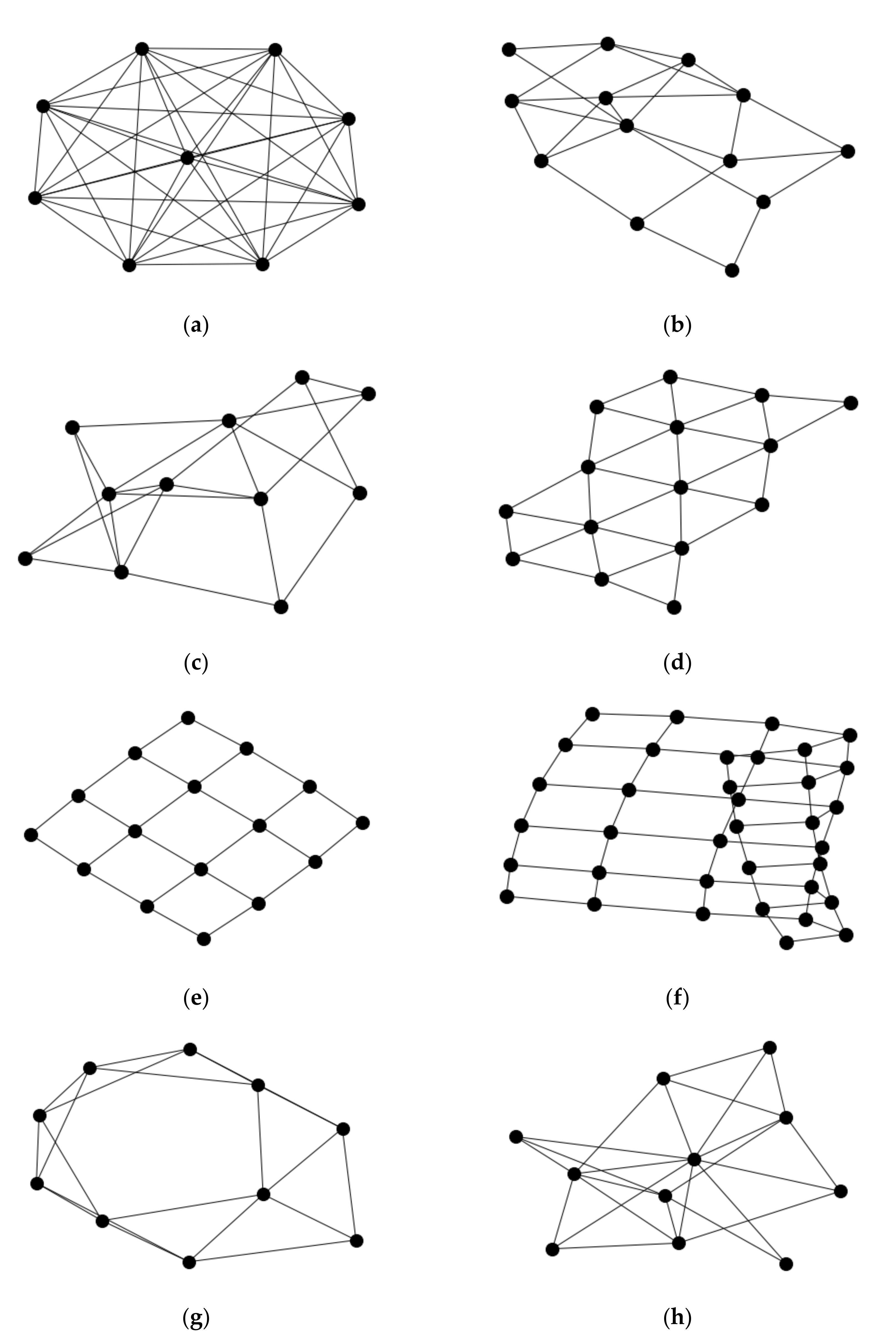
2.1. Simulating Neural Network Structure
2.2. Fruit Fly Hemibrain Data
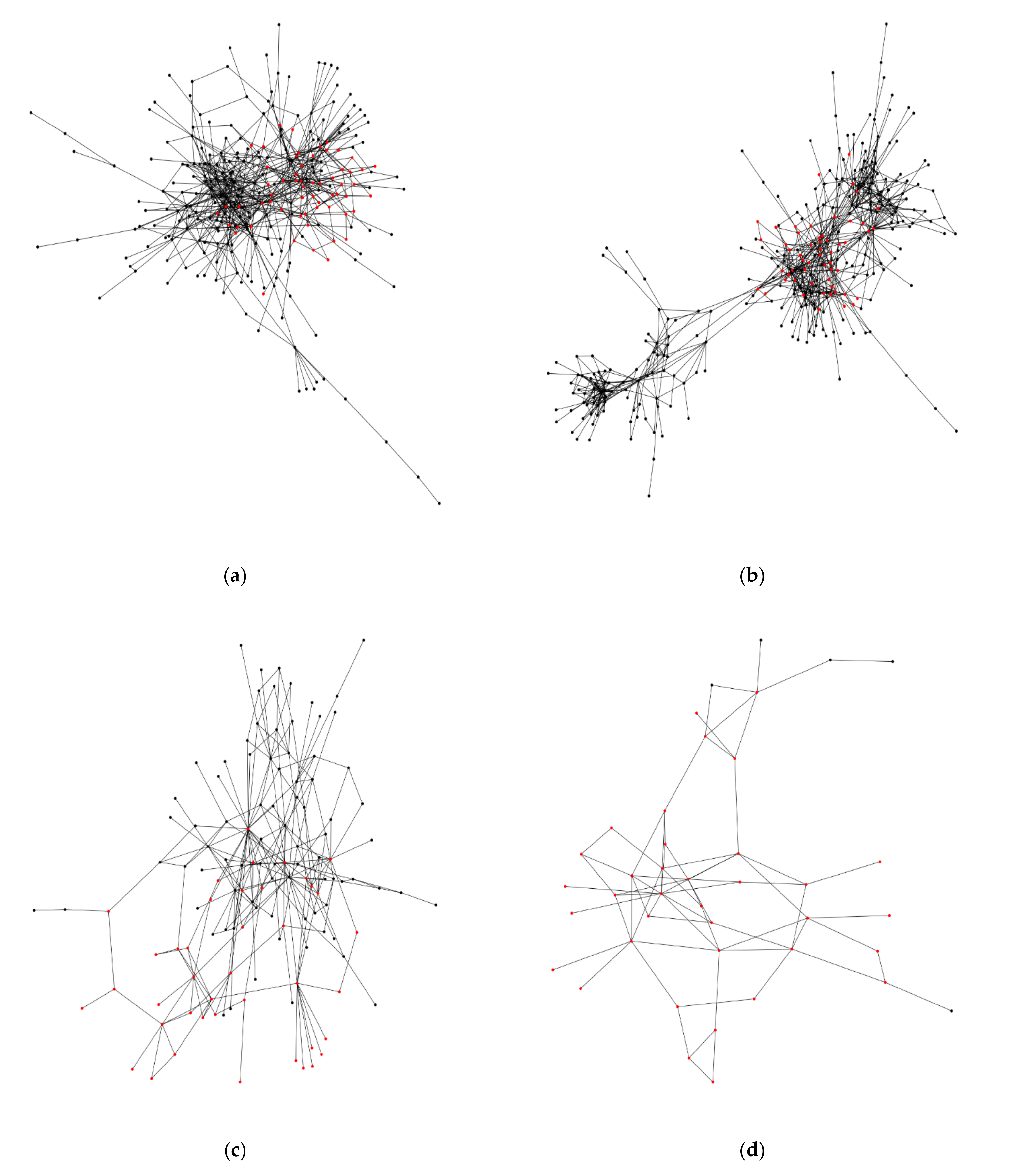
2.3. Analysis of the Hemibrain Data
2.4. Computational Resources
3. Results
3.1. Assessing Parameters of the Simulated Networks
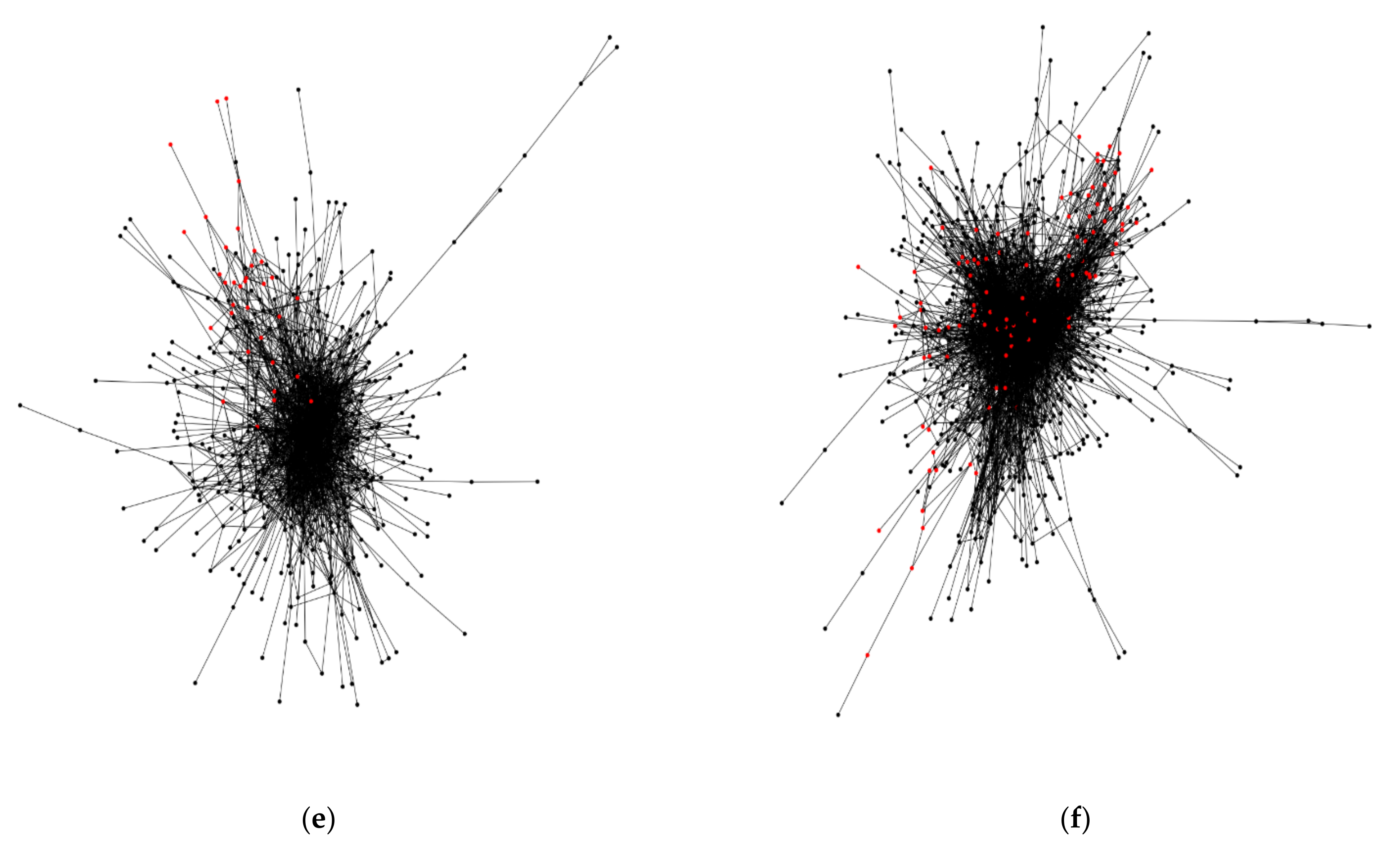
3.2. Square Grid Arrangement in the Fruit Fly Brain
4. Discussion
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bassett, D.S.; Bullmore, E.D. Small-world brain networks. Neuroscientist 2006, 12, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Telesford, Q.K.; Joyce, K.E.; Hayasaka, S.; Burdette, J.H.; Laurienti, P.J. The ubiquity of small-world networks. Brain Connect. 2011, 1, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzsaki, G.; Moser, E.I. Memory, navigation and theta rhythm in the hippocampalentorhinal system. Nat. Neurosci. 2013, 16, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, M.B.; Rowland, D.C.; Moser, E.I. Place cells, grid cells, and memory. Cold Spring Harb. Perspect. Biol. 2015, 7, a021808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantinescu, A.O.; O’Reilly, J.X.; Behrens, T.E. Organizing conceptual knowledge in humans with a gridlike code. Science 2016, 352, 1464–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monsalve-Mercado, M.M.; Leibold, C. Hippocampal spike-timing correlations lead to hexagonal grid fields. Phys. Rev. Lett. 2017, 119, 038101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada, E.; Rodriguez-Velazquez, J.A. Spectral measures of bipartivity in complex networks. Phys. Rev. E 2005, 72, 046105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, S.; Cunningham, J.P. Towards the neural population doctrine. Curr. Opin. Neurobiol. 2019, 55, 103–111. [Google Scholar] [CrossRef]
- Shannon, C. The lattice theory of information. Trans. IRE Prof. Group Inf. Theory 1953, 1, 105–107. [Google Scholar] [CrossRef]
- Goni, J.; Avena-Koenigsberger, A.; Velez de Mendizabal, N.; van den Heuvel, M.P.; Betzel, R.F.; Sporns, O. Exploring the morphospace of communication efficiency in complex networks. PLoS ONE 2013, 8, e58070. [Google Scholar]
- Eytan, D.; Marom, S. Dynamics and effective topology underlying synchronization in networks of cortical neurons. J. Neurosci. 2006, 26, 8465–8476. [Google Scholar] [CrossRef] [Green Version]
- Paraskevov, A.V.; Zendrikov, D.K. A spatially resolved network spike in model neuronal cultures reveals nucleation centers, circular traveling waves and drifting spiral waves. Phys. Biol. 2017, 14, 026003. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, L.K.; Xu, C.S.; Januszewski, M.; Lu, Z.; Takemura, S.-Y.; Hayworth, K.J.; Huang, G.B.; Shinomiya, K.; Maitlin-Shepard, J.; Berg, S.; et al. A connectome and analysis of the adult Drosophila central brain. eLife 2020, 9, e57443. [Google Scholar] [CrossRef]
- Albert, R.; Barabasi, A.L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47. [Google Scholar] [CrossRef] [Green Version]
- Hagberg, A.A.; Schult, D.A.; Swart, P.J. Exploring network structure, dynamics, and function using NetworkX. In Proceedings of the 7th Python in Science Conference, SciPy2008, Pasadena, CA, USA, 19–24 August 2008; Varoquaux, G., Vaught, T., Millman, J., Eds.; pp. 11–15. [Google Scholar]
- Lind, P.G.; Gonzalez, M.C.; Herrmann, H.J. Cycles and clustering in bipartite networks. Phys. Rev. E 2005, 72, 056127. [Google Scholar] [CrossRef] [Green Version]
- Humphries, M.D.; Gurney, K. Network ‘small-world-ness’: A quantitative method for determining canonical network equivalence. PLoS ONE 2008, 3, e0002051. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Shinomiya, K.; Ito, M.; Armstrong, D.; Boyan, G.; Hartenstein, V.; Harzsch, S.; Heisenberg, M.; Homberg, U.; Jenett, A.; et al. A systematic nomenclature for the insect brain. Neuron 2014, 81, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Bisong, E. Google Colaboratory. In Building Machine Learning and Deep Learning Models on Google Cloud Platform; Apress: Berkeley, CA, USA, 2019. [Google Scholar]
- Kluyver, T.; Ragan-Kelley, B.; Perez, F.; Granger, B.E.; Bussonnier, M.; Frederic, J.; Kelley, K.; Hamrick, J.B.; Grout, J.; Corlay, S.; et al. Jupyter Notebooks—A Publishing Format for Reproducible Computational Workflows; IOS Press: Amsterdam, Netherlands, 2016; pp. 87–90. [Google Scholar]
- Hafting, T.; Fyhn, M.; Molden, S.; Moser, M.B.; Moser, E.I. Microstructure of a spatial map in the entorhinal cortex. Nature 2005, 436, 801–806. [Google Scholar] [CrossRef]
- Scheffer, L.K.; Meinertzhagen, I.A. A connectome is not enough–what is still needed to understand the brain of Drosophila? J. Exp. Biol. 2021, 224, jeb242740. [Google Scholar] [CrossRef]
- Menzel, R. The honeybee as a model for understanding the basis of cognition. Nat. Rev. Neurosci. 2012, 13, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Eichler, K.; Li, F.; Litwin-Kumar, A.; Park, Y.; Andrade, I.; Schneider-Mizell, C.M.; Saumweber, T.; Huser, A.; Eschbach, C.; Gerber, B.; et al. The complete connectome of a learning and memory centre in an insect brain. Nature 2017, 548, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lindsey, J.W.; Marin, E.C.; Otto, N.; Dreher, M.; Dempsey, G.; Stark, I.; Bates, A.S.; Pleijzier, M.W.; Schlegel, P.; et al. The connectome of the adult Drosophila mushroom body provides insights into function. eLife 2020, 9, e62576. [Google Scholar] [CrossRef] [PubMed]
- Givon, L.E.; Lazar, A.A.; Yeh, C.H. Generating executable models of the Drosophila central complex. Front. Behav. Neurosci. 2017, 11, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neriec, N.; Desplan, C. From the eye to the brain: Development of the Drosophila visual system. Curr. Top. Dev. Biol. 2016, 116, 247–271. [Google Scholar] [PubMed] [Green Version]
- Borst, A.; Helmstaedter, M. Common circuit design in fly and mammalian motion vision. Nat. Neurosci. 2015, 18, 1067–1076. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neuropil | Brain Region | Abb. | Neuropil | Brain Region | Abb. |
|---|---|---|---|---|---|
| Optic Lobe | Medulla | ME | Lateral Horn | Lateral Horn | LH |
| Accessory Medulla | AME | Superior Neuropils | Superior Lateral Protocerebrum | SLP | |
| Lobula | LO | Superior Intermediate Protocerebrum | SIP | ||
| Lobula Plate | LOP | Superior Medial Protocerebrum | SMP | ||
| Mushroom Body | Calyx | CA | Inferior Neuropils | Crepine | CRE |
| Pedunculus | PED | Superior Clamp | SCL | ||
| Alpha Lobe | aL | Inferior Clamp | ICL | ||
| Alpha Prime Lobe | a’L | Inferior Bridge | IB | ||
| Beta Lobe | bL | Antler | ATL | ||
| Beta Prime Lobe | b’L | Antennal Lobe | Antennal Lobe | AL/AL2 | |
| Gamma Lobe | gL | Ventromedial Neuropils | Ves | VES | |
| Central Complex | Fan Shaped Body | FB | Epaulette | EPA | |
| Asymmetric Body | AB | Gorget | GOR | ||
| Epsilloid Body | EB | Superior Posterior Slope | SPS | ||
| Protocerebral Bridge | PB | Inferior Posterior Slope | IPS | ||
| Noduli | NO | Periesophageal Neuropils | Saddle | SAD | |
| Lateral Complex | Bulb | BU | Flange | FLA | |
| Lateral Accessory Lobe | LAL | Cantle | CAN | ||
| Ventrolateral Neuropils | Anterior Optic Tubercle | AOTU | Gnathal Ganglia | Gnathal Ganglia | GNG |
| Anterior Ventrolateral Protocerebrum | AVLP | ||||
| Posterior Ventrolateral Protocerebrum | PVLP | ||||
| Posteriorlateral Protocerebrum | PLP | ||||
| Wedge | WED |
| Network Model | Node Count | Model Parameters | Clustering Coefficient (Min, Max) | Transitivity (Min, Max) | Bipartivity (Min, Max) | Sigma (Min, Max) |
|---|---|---|---|---|---|---|
| Complete | 9 to 27 | - | 1.0, 1.0 | 1.0, 1.0 | 0.5, 0.5 | 1.0, 1.0 |
| Square grid | 9 to 81 | - | 0.16, 0.28 | 0.0, 0.0 | 1.0, 1.0 | 0.0, 0.0 |
| Triangular grid | 9 to 81 | - | 0.13, 2.0 | 0.42, 0.49 | 0.63, 0.70 | 1.26, 3.96 |
| Random | 9 to 27 | P = 0.4, 0.5 | 0.07, 0.40 | 0.14, 0.60 | 0.50, 0.93 | 0.89, 1.23 |
| Preferential attach | 9 to 27 | M = 3 to 6 | 0.15, 0.56 | 0.20, 0.67 | 0.50, 0.65 | 0.75, 1.08 |
| Small world | 9 to 27 | k = 4 to 6 P = 0.1, 0.5 | 0.05, 0.55 | 0.08, 0.72 | 0.54, 0.89 | 0.60, 4.88 |
| Parameter | Utility of Parameter | Parameter Values |
|---|---|---|
| Node Count | 36 to ∞ | |
| Transitivity | Filter for triangular grid | 0.0 to 0.20 |
| Bipartivity | Filter for square grid | 0.80 to 1.0 |
| Sigma | Filter for non-square grid | 0.0 to 0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedman, R. Detecting Square Grid Structure in an Animal Neuronal Network. NeuroSci 2022, 3, 91-103. https://doi.org/10.3390/neurosci3010007
Friedman R. Detecting Square Grid Structure in an Animal Neuronal Network. NeuroSci. 2022; 3(1):91-103. https://doi.org/10.3390/neurosci3010007
Chicago/Turabian StyleFriedman, Robert. 2022. "Detecting Square Grid Structure in an Animal Neuronal Network" NeuroSci 3, no. 1: 91-103. https://doi.org/10.3390/neurosci3010007
APA StyleFriedman, R. (2022). Detecting Square Grid Structure in an Animal Neuronal Network. NeuroSci, 3(1), 91-103. https://doi.org/10.3390/neurosci3010007