
The Consciousness of Pain: A Thalamocortical Perspective

{kind=link}
Abstract
:1. Introduction
2. Inducible Unconsciousness
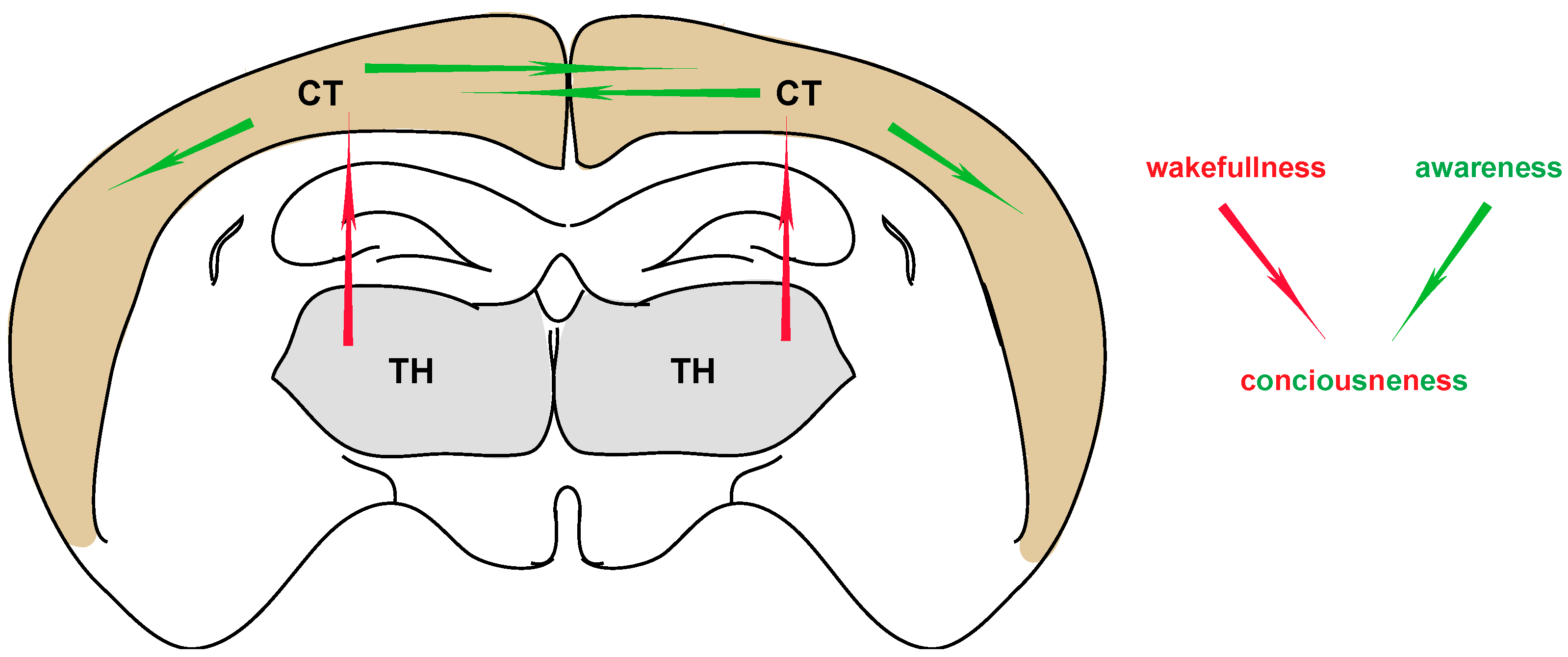
2.1. Loss of Consciousness and Anesthesia
2.2. Thalamocortical Connections in Anesthesia
3. The Percept of Pain
3.1. Pain and Consciousness
3.2. Thalamocortical Connectivity in Pain
4. Conclusions and Perspective
4.1. Pain: Evolution, Adaptation and Control
4.2. Pain as a Conscious Experience
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Voss, U.; Schermelleh-Engel, K.; Windt, J.; Frenzel, C.; Hobson, A. Measuring consciousness in dreams: The lucidity and consciousness in dreams scale. Conscious Cogn. 2013, 22, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Laureys, S.; Boly, M. Unresponsive wakefulness syndrome. Arch. Ital. Biol. 2012, 150, 31–35. [Google Scholar] [PubMed]
- Le, A.; Stojanoski, B.B.; Khan, S.; Keough, M.; Niemeier, M. A toggle switch of visual awareness? Cortex 2015, 64, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.K. Consciousness: The last 50 years (and the next). Brain Neurosci. Adv. 2018, 2, 2398212818816019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelazo, P.D. The development of conscious control in childhood. Trends Cogn. Sci. 2004, 8, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Searle, J.R. Consciousness. Annu. Rev. Neurosci. 2000, 23, 557–578. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Vanneste, S.; Smith, M.; Adhia, D. Pain and the Triple Network Model. Front. Neurol. 2022, 13, 757241. [Google Scholar] [CrossRef]
- Perl, E.R. Ideas about pain, a historical view. Nat. Rev. Neurosci. 2007, 8, 71–80. [Google Scholar] [CrossRef]
- Hippocrates. On the articulations. The genuine works of Hippocrates. Clin. Orthop. Relat. Res. 2002, 400, 19–25. [Google Scholar]
- Tashani, O.A.; Johnson, M.I. Avicenna′s concept of pain. Libyan J. Med. 2010, 5, 5253. [Google Scholar] [CrossRef]
- Benini, A.; DeLeo, J.A. Rene Descartes′ physiology of pain. Spine (Phila Pa 1976) 1999, 24, 2115–2119. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.D.; Flor, H.; Greely, H.T.; Iannetti, G.D.; Mackey, S.; Ploner, M.; Pustilnik, A.; Tracey, I.; Treede, R.D.; Wager, T.D. Brain imaging tests for chronic pain: Medical, legal and ethical issues and recommendations. Nat. Rev. Neurol. 2017, 13, 624–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meacham, K.; Shepherd, A.; Mohapatra, D.P.; Haroutounian, S. Neuropathic Pain: Central vs. Peripheral Mechanisms. Curr. Pain Headache Rep. 2017, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- St John Smith, E. Advances in understanding nociception and neuropathic pain. J. Neurol. 2018, 265, 231–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni Mhuircheartaigh, R.; Warnaby, C.; Rogers, R.; Jbabdi, S.; Tracey, I. Slow-wave activity saturation and thalamocortical isolation during propofol anesthesia in humans. Sci. Transl. Med. 2013, 5, 208ra148. [Google Scholar] [CrossRef]
- Zasler, N.D.; Formisano, R.; Aloisi, M. Pain in Persons with Disorders of Consciousness. Brain Sci. 2022, 12, 300. [Google Scholar] [CrossRef]
- Clark, J.A.; Brown, C.A.; Jones, A.K.; El-Deredy, W. Dissociating nociceptive modulation by the duration of pain anticipation from unpredictability in the timing of pain. Clin. Neurophysiol. 2008, 119, 2870–2878. [Google Scholar] [CrossRef]
- Wiech, K.; Kalisch, R.; Weiskopf, N.; Pleger, B.; Stephan, K.E.; Dolan, R.J. Anterolateral prefrontal cortex mediates the analgesic effect of expected and perceived control over pain. J. Neurosci. 2006, 26, 11501–11509. [Google Scholar] [CrossRef] [Green Version]
- Bingel, U.; Wanigasekera, V.; Wiech, K.; Ni Mhuircheartaigh, R.; Lee, M.C.; Ploner, M.; Tracey, I. The effect of treatment expectation on drug efficacy: Imaging the analgesic benefit of the opioid remifentanil. Sci. Transl. Med. 2011, 3, 70ra14. [Google Scholar] [CrossRef] [Green Version]
- Mashour, G.A. Top-down mechanisms of anesthetic-induced unconsciousness. Front. Syst. Neurosci. 2014, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K. The role of dendritic signaling in the anesthetic suppression of consciousness. Anesthesiology 2015, 122, 1415–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkire, M.T.; Hudetz, A.G.; Tononi, G. Consciousness and anesthesia. Science 2008, 322, 876–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashour, G.A.; Hudetz, A.G. Bottom-Up and Top-Down Mechanisms of General Anesthetics Modulate Different Dimensions of Consciousness. Front. Neural Circuits 2017, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, L.M. The thalamic dynamic core theory of conscious experience. Conscious Cogn. 2011, 20, 464–486. [Google Scholar] [CrossRef] [PubMed]
- Hudetz, A.G. General anesthesia and human brain connectivity. Brain Connect. 2012, 2, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Tai, S.K.; Leung, L.S. Ketamine-induced deficit of auditory gating in the hippocampus of rats is alleviated by medial septal inactivation and antipsychotic drugs. Psychopharmacology (Berl) 2009, 206, 457–467. [Google Scholar] [CrossRef]
- Krystal, J.H.; Karper, L.P.; Seibyl, J.P.; Freeman, G.K.; Delaney, R.; Bremner, J.D.; Heninger, G.R.; Bowers, M.B.; Charney, D.S. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch. Gen. Psychiatry 1994, 51, 199–214. [Google Scholar] [CrossRef]
- Brohan, J.; Goudra, B.G. The Role of GABA Receptor Agonists in Anesthesia and Sedation. CNS Drugs 2017, 31, 845–856. [Google Scholar] [CrossRef]
- Antkowiak, B.; Helfrich-Forster, C. Effects of small concentrations of volatile anesthetics on action potential firing of neocortical neurons in vitro. Anesthesiology 1998, 88, 1592–1605. [Google Scholar] [CrossRef]
- Antkowiak, B. Different actions of general anesthetics on the firing patterns of neocortical neurons mediated by the GABA(A) receptor. Anesthesiology 1999, 91, 500–511. [Google Scholar] [CrossRef] [Green Version]
- Ali, F.; Gerhard, D.M.; Sweasy, K.; Pothula, S.; Pittenger, C.; Duman, R.S.; Kwan, A.C. Ketamine disinhibits dendrites and enhances calcium signals in prefrontal dendritic spines. Nat. Commun. 2020, 11, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, G.; Han, F.; Shi, W.X. Effects of Low Doses of Ketamine on Pyramidal Neurons in Rat Prefrontal Cortex. Neuroscience 2018, 384, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.D.; Voigts, J.; Flores, F.J.; Schmitt, L.I.; Wilson, M.A.; Halassa, M.M.; Brown, E.N. Thalamic reticular nucleus induces fast and local modulation of arousal state. Elife 2015, 4, e08760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, C.; Zhang, L.; Zhou, W.; Yu, S.; Yi, R.; Luo, D.; Fu, X. Effects of Propofol on Electrical Synaptic Strength in Coupling Reticular Thalamic GABAergic Parvalbumin-Expressing Neurons. Front. Neurosci. 2020, 14, 364. [Google Scholar] [CrossRef]
- Lu, J.; Nelson, L.E.; Franks, N.; Maze, M.; Chamberlin, N.L.; Saper, C.B. Role of endogenous sleep-wake and analgesic systems in anesthesia. J. Comp. Neurol. 2008, 508, 648–662. [Google Scholar] [CrossRef] [Green Version]
- Moody, O.A.; Zhang, E.R.; Vincent, K.F.; Kato, R.; Melonakos, E.D.; Nehs, C.J.; Solt, K. The Neural Circuits Underlying General Anesthesia and Sleep. Anesth. Analg. 2021, 132, 1254–1264. [Google Scholar] [CrossRef]
- Akeju, O.; Loggia, M.L.; Catana, C.; Pavone, K.J.; Vazquez, R.; Rhee, J.; Ramirez, V.C.; Chonde, D.B.; Izquierdo-Garcia, D.; Arabasz, G.; et al. Disruption of thalamic functional connectivity is a neural correlate of dexmedetomidine-induced unconsciousness. Elife 2014, 3, e04499. [Google Scholar] [CrossRef]
- Mashour, G.A. Consciousness, anesthesia, and neural synchrony. Anesthesiology 2013, 119, 7–9. [Google Scholar] [CrossRef]
- Baker, R.; Gent, T.C.; Yang, Q.; Parker, S.; Vyssotski, A.L.; Wisden, W.; Brickley, S.G.; Franks, N.P. Altered activity in the central medial thalamus precedes changes in the neocortex during transitions into both sleep and propofol anesthesia. J. Neurosci. 2014, 34, 13326–13335. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhu, X.H.; Zhang, Y.; Chen, W. The change of functional connectivity specificity in rats under various anesthesia levels and its neural origin. Brain Topogr. 2013, 26, 363–377. [Google Scholar] [CrossRef] [Green Version]
- Redinbaugh, M.J.; Phillips, J.M.; Kambi, N.A.; Mohanta, S.; Andryk, S.; Dooley, G.L.; Afrasiabi, M.; Raz, A.; Saalmann, Y.B. Thalamus Modulates Consciousness via Layer-Specific Control of Cortex. Neuron 2020, 106, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Malekmohammadi, M.; Price, C.M.; Hudson, A.E.; Di Cesare, J.A.T.; Pouratian, N. Propofol-induced loss of consciousness is associated with a decrease in thalamocortical connectivity in humans. Brain 2019, 142, 2288–2302. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Larkum, M.E. General Anesthesia Decouples Cortical Pyramidal Neurons. Cell 2020, 180, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G. An information integration theory of consciousness. BMC Neurosci. 2004, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Tononi, G.; Boly, M.; Massimini, M.; Koch, C. Integrated information theory: From consciousness to its physical substrate. Nat. Rev. Neurosci. 2016, 17, 450–461. [Google Scholar] [CrossRef]
- Jensen, K.; Kirsch, I.; Odmalm, S.; Kaptchuk, T.J.; Ingvar, M. Classical conditioning of analgesic and hyperalgesic pain responses without conscious awareness. Proc. Natl. Acad. Sci. USA 2015, 112, 7863–7867. [Google Scholar] [CrossRef] [Green Version]
- Fardo, F.; Allen, M.; Jegindo, E.M.; Angrilli, A.; Roepstorff, A. Neurocognitive evidence for mental imagery-driven hypoalgesic and hyperalgesic pain regulation. Neuroimage 2015, 120, 350–361. [Google Scholar] [CrossRef]
- Bastuji, H.; Frot, M.; Perchet, C.; Magnin, M.; Garcia-Larrea, L. Pain networks from the inside: Spatiotemporal analysis of brain responses leading from nociception to conscious perception. Hum. Brain Mapp. 2016, 37, 4301–4315. [Google Scholar] [CrossRef]
- Grant, J.A.; Zeidan, F. Employing pain and mindfulness to understand consciousness: A symbiotic relationship. Curr. Opin. Psychol. 2019, 28, 192–197. [Google Scholar] [CrossRef]
- Zuo, N.; Song, M.; Fan, L.; Eickhoff, S.B.; Jiang, T. Different interaction modes for the default mode network revealed by resting state functional magnetic resonance imaging. Eur. J. Neurosci. 2016, 43, 78–88. [Google Scholar] [CrossRef]
- Vatansever, D.; Menon, D.K.; Manktelow, A.E.; Sahakian, B.J.; Stamatakis, E.A. Default Mode Dynamics for Global Functional Integration. J. Neurosci. 2015, 35, 15254–15262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demertzi, A.; Soddu, A.; Laureys, S. Consciousness supporting networks. Curr. Opin. Neurobiol. 2013, 23, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponten, M.; Fust, J.; D’Onofrio, P.; Dorp, R.V.; Sunnergård, L.; Ingre, M.; Axelsson, J.; Jensen, K. The pain alarm response—An example of how conscious awareness shapes pain perception. Sci. Rep. 2019, 9, 12478. [Google Scholar] [CrossRef] [PubMed]
- Kolodny, O.; Moyal, R.; Edelman, S. A possible evolutionary function of phenomenal conscious experience of pain. Neurosci. Conscious 2021, 2021, niab012. [Google Scholar] [CrossRef]
- Wager, T.D.; Atlas, L.Y.; Lindquist, M.A.; Roy, M.; Woo, C.W.; Kross, E. An fMRI-based neurologic signature of physical pain. N. Engl. J. Med. 2013, 368, 1388–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, A.; Larsen, B.; Baller, E.B.; Scott, J.C.; Sharma, V.; Adebimpe, A.; Basbaum, A.I.; Dworkin, R.H.; Edwards, R.R.; Woolf, C.J.; et al. Convergent neural representations of experimentally-induced acute pain in healthy volunteers: A large-scale fMRI meta-analysis. Neurosci. Biobehav. Rev. 2020, 112, 300–323. [Google Scholar] [CrossRef] [PubMed]
- Kuner, R.; Kuner, T. Cellular Circuits in the Brain and Their Modulation in Acute and Chronic Pain. Physiol. Rev. 2021, 101, 213–258. [Google Scholar] [CrossRef]
- Craig, A.D.; Bushnell, M.C.; Zhang, E.T.; Blomqvist, A. A thalamic nucleus specific for pain and temperature sensation. Nature 1994, 372, 770–773. [Google Scholar] [CrossRef]
- Blomqvist, A.; Zhang, E.T.; Craig, A.D. Cytoarchitectonic and immunohistochemical characterization of a specific pain and temperature relay, the posterior portion of the ventral medial nucleus, in the human thalamus. Brain 2000, 123, 601–619. [Google Scholar] [CrossRef] [Green Version]
- Dum, R.P.; Levinthal, D.J.; Strick, P.L. The spinothalamic system targets motor and sensory areas in the cerebral cortex of monkeys. J. Neurosci. 2009, 29, 14223–14235. [Google Scholar] [CrossRef] [Green Version]
- Sumser, A.; Mease, R.A.; Sakmann, B.; Groh, A. Organization and somatotopy of corticothalamic projections from L5B in mouse barrel cortex. Proc. Natl. Acad. Sci. USA 2017, 114, 8853–8858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, S.M. Thalamocortical interactions. Curr. Opin. Neurobiol. 2012, 22, 575–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, S.M. Thalamus plays a central role in ongoing cortical functioning. Nat. Neurosci. 2016, 19, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Chang, J.Y.; Woodward, D.J.; Baccala, L.A.; Han, J.S.; Luo, F. Corticofugal influences on thalamic neurons during nociceptive transmission in awake rats. Synapse 2007, 61, 335–342. [Google Scholar] [CrossRef]
- Meda, K.S.; Patel, T.; Braz, J.M.; Malik, R.; Turner, M.L.; Seifikar, H.; Basbaum, A.I.; Sohal, V.S. Microcircuit Mechanisms through which Mediodorsal Thalamic Input to Anterior Cingulate Cortex Exacerbates Pain-Related Aversion. Neuron 2019, 102, 944–959. [Google Scholar] [CrossRef]
- Garcia-Larrea, L.; Bastuji, H. Pain and consciousness. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 87, 193–199. [Google Scholar] [CrossRef]
- Nagasako, E.M.; Oaklander, A.L.; Dworkin, R.H. Congenital insensitivity to pain: An update. Pain 2003, 101, 213–219. [Google Scholar] [CrossRef]
- Huskisson, E.C.; Berry, H.; Gishen, P.; Jubb, R.W.; Whitehead, J. Effects of antiinflammatory drugs on the progression of osteoarthritis of the knee. LINK Study Group. Longitudinal Investigation of Nonsteroidal Antiinflammatory Drugs in Knee Osteoarthritis. J. Rheumatol. 1995, 22, 1941–1946. [Google Scholar]
- Nesse, R.M.; Schulkin, J. An evolutionary medicine perspective on pain and its disorders. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190288. [Google Scholar] [CrossRef]
- Eippert, F.; Bingel, U.; Schoell, E.D.; Yacubian, J.; Klinger, R.; Lorenz, J.; Büchel, C. Activation of the opioidergic descending pain control system underlies placebo analgesia. Neuron 2009, 63, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Price, D.D.; Harkins, S.W.; Baker, C. Sensory-affective relationships among different types of clinical and experimental pain. Pain 1987, 28, 297–307. [Google Scholar] [CrossRef]
- Chen, T.; Wang, J.; Wang, Y.Q.; Chu, Y.X. Current Understanding of the Neural Circuitry in the Comorbidity of Chronic Pain and Anxiety. Neural Plast. 2022, 2022, 4217593. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Koch, C. Consciousness: Here, there and everywhere? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, C. What Is Consciousness? Sci. Am. 2018, 318, 60–64. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sgourdou, P. The Consciousness of Pain: A Thalamocortical Perspective. NeuroSci 2022, 3, 311-320. https://doi.org/10.3390/neurosci3020022
Sgourdou P. The Consciousness of Pain: A Thalamocortical Perspective. NeuroSci. 2022; 3(2):311-320. https://doi.org/10.3390/neurosci3020022
Chicago/Turabian StyleSgourdou, Paraskevi. 2022. "The Consciousness of Pain: A Thalamocortical Perspective" NeuroSci 3, no. 2: 311-320. https://doi.org/10.3390/neurosci3020022
APA StyleSgourdou, P. (2022). The Consciousness of Pain: A Thalamocortical Perspective. NeuroSci, 3(2), 311-320. https://doi.org/10.3390/neurosci3020022