


Contribution of smFRET to Chromatin Research


Abstract
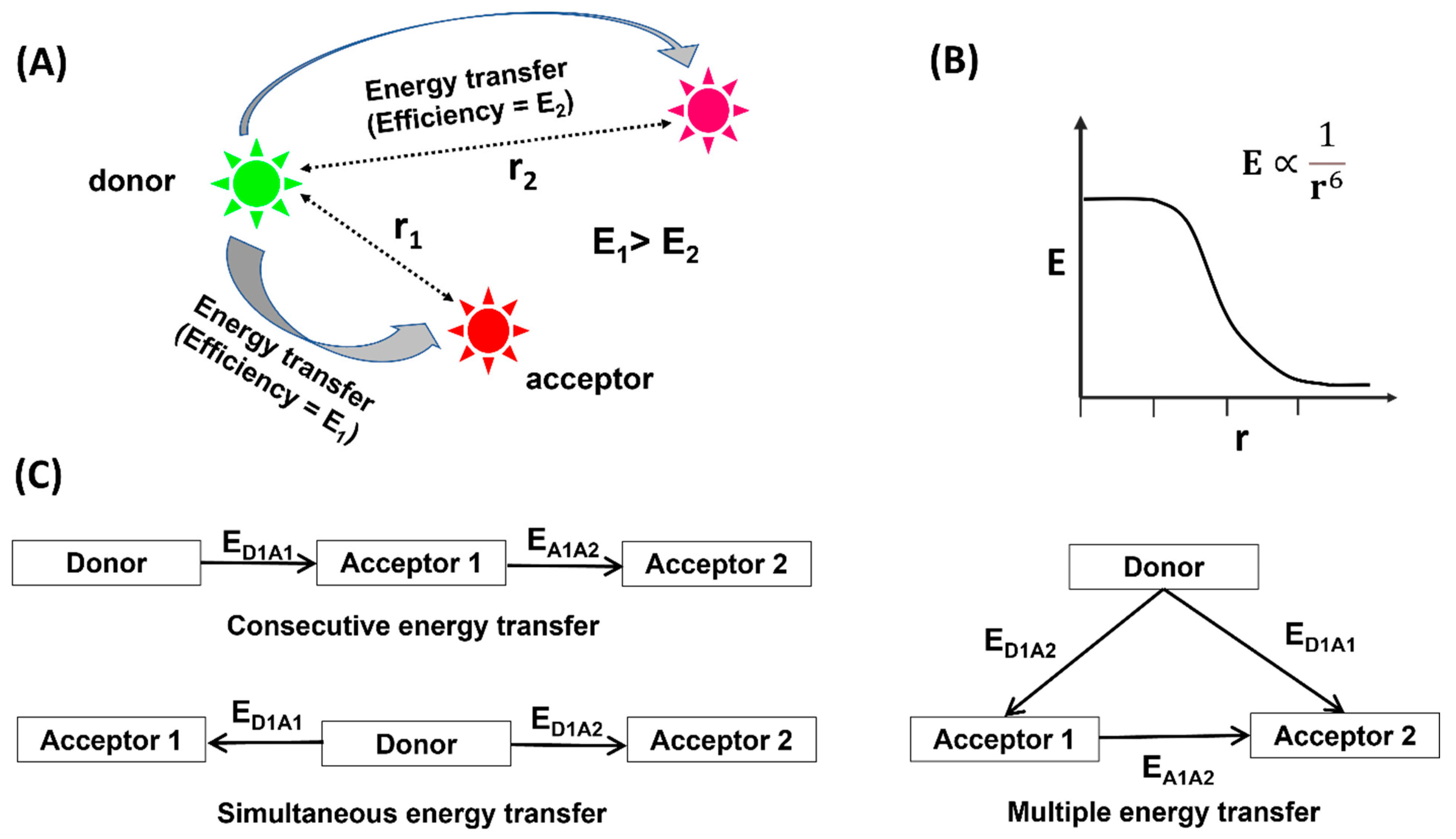
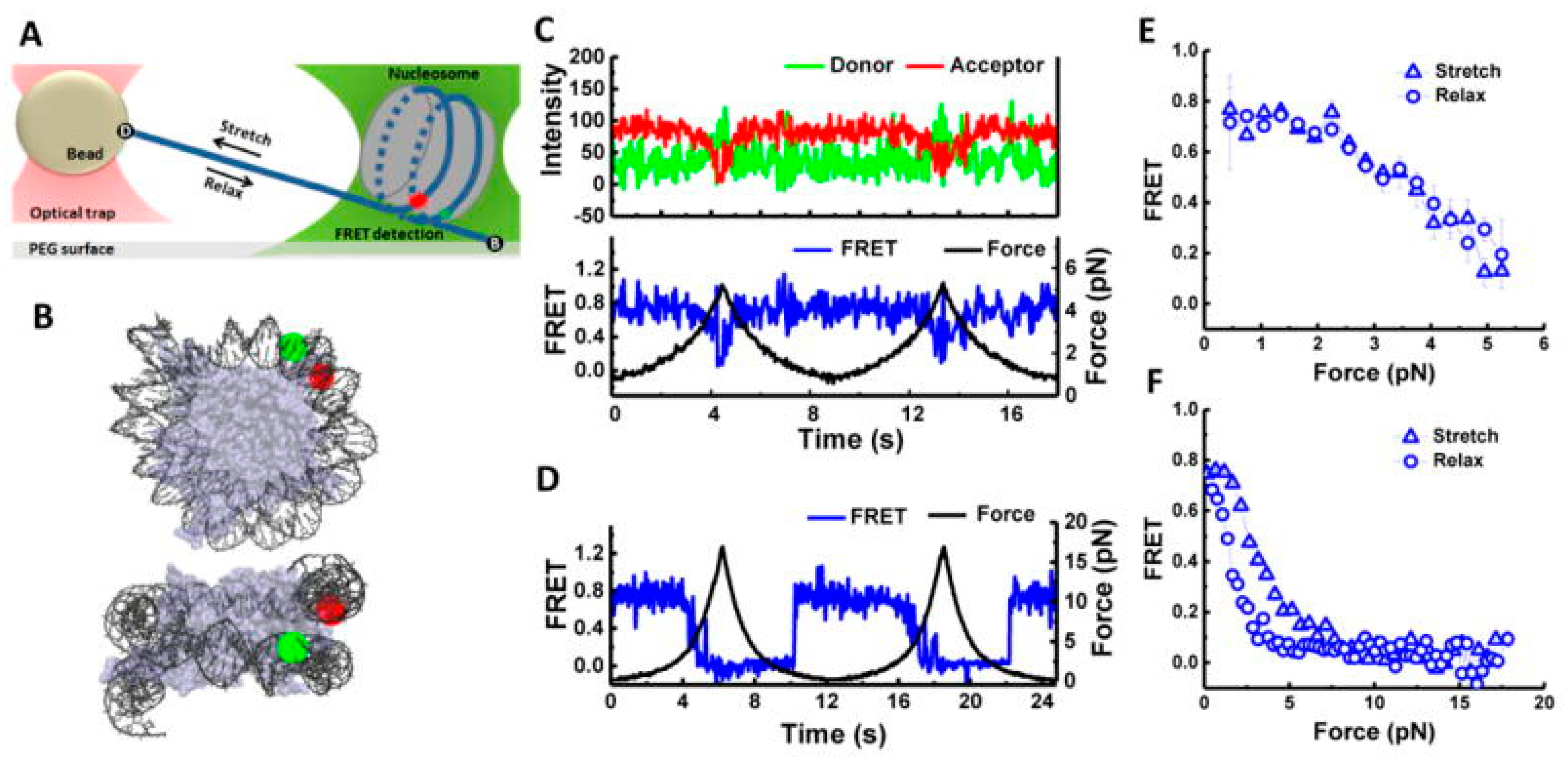
:1. Introduction
- Chromatin dynamics and compaction;
- Interaction with chaperones and other protein machinery.
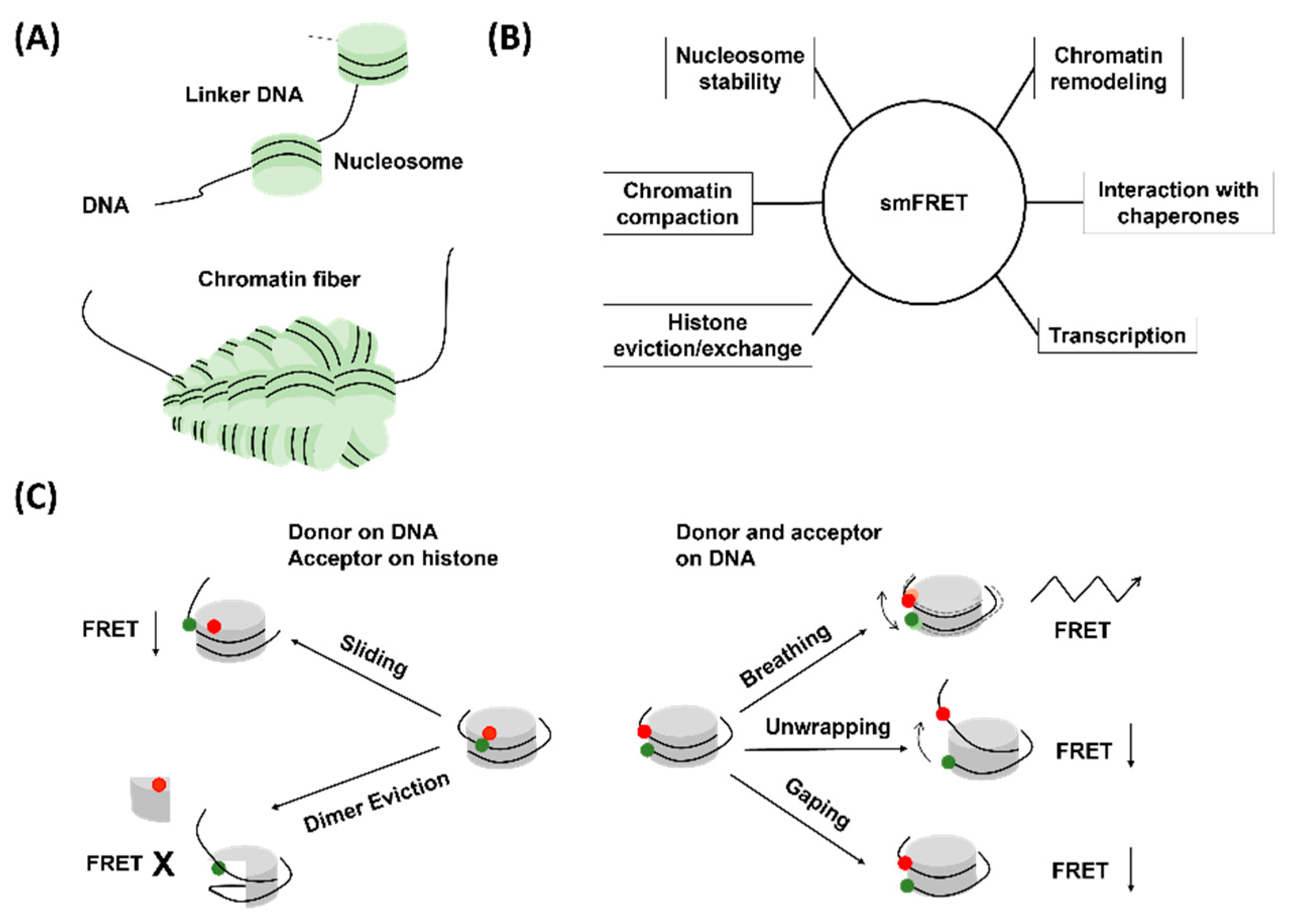
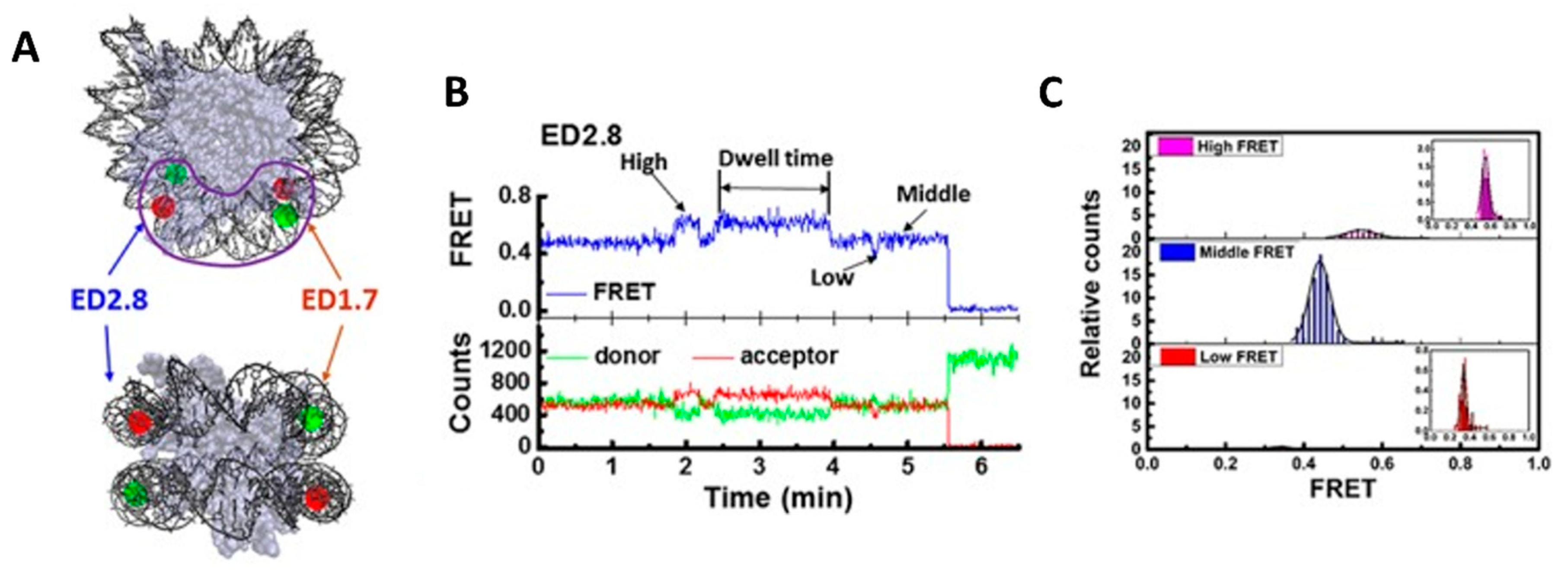
2. Chromatin Structures and Dynamics
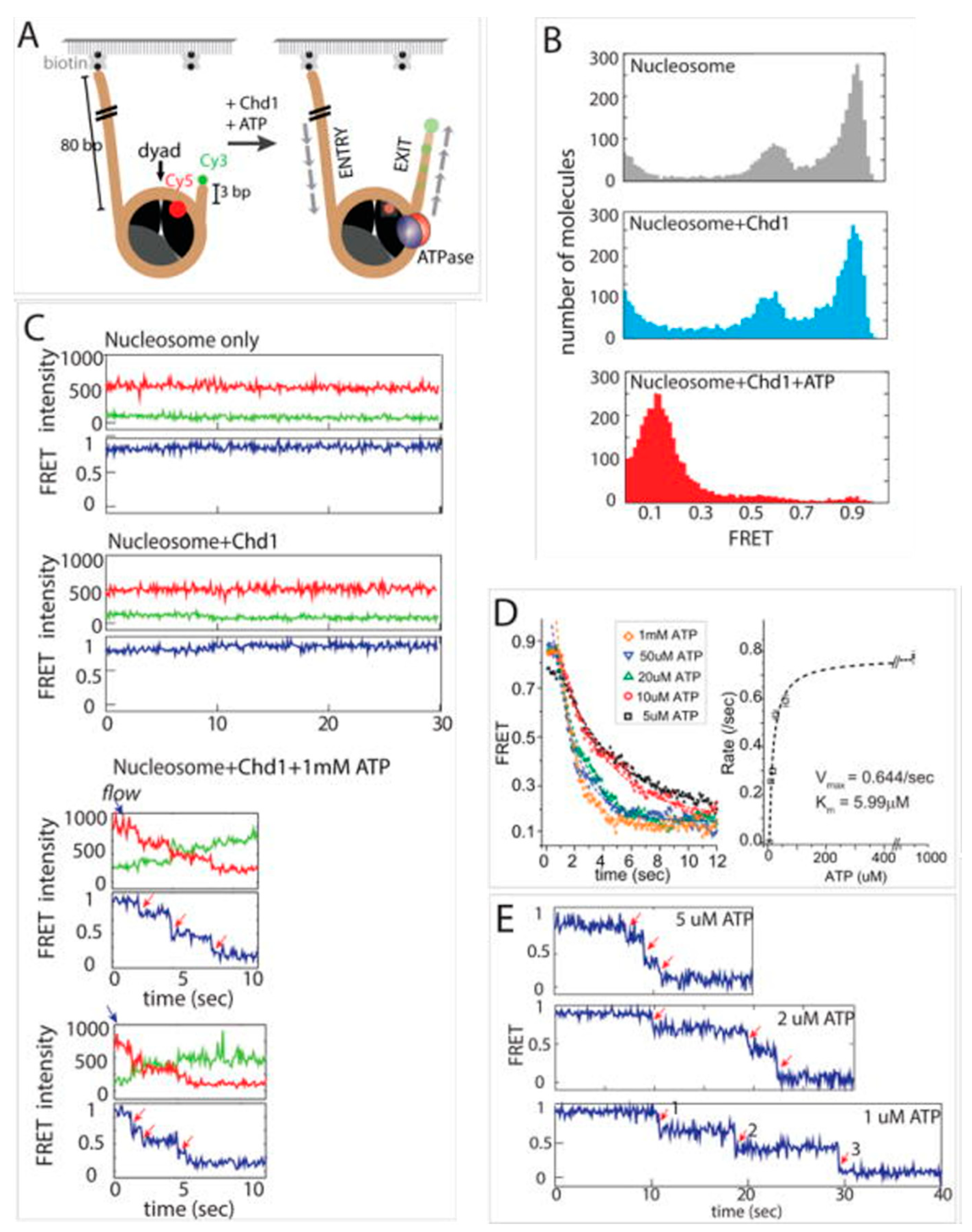
3. Interaction of Chromatins with Protein Machineries
4. Summary and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Clegg, R. Reviews in Fluorescence. 2006. Available online: https://link.springer.com/book/10.1007/0-387-33016-X (accessed on 20 November 2022).
- Sun, Y.; Wallrabe, H.; Seo, S.-A.; Periasamy, A. FRET microscopy in 2010: The legacy of Theodor Förster on the 100th anniversary of his birth. Chemphyschem: A Eur. J. Chem. Phys. Phys. Chem. 2011, 12, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, A.; Kay, S.; Day, R. Fluorescence Resonance Energy Transfer (FRET) Imaging of a Single Living Cell Using Green Fluorescent Protein; SPIE: San Jose, CA, USA, 1997; Volume 2983. [Google Scholar]
- Chirio-Lebrun, M.-C.; Prats, M. Fluorescence resonance energy transfer (FRET): Theory and experiments. Biochem. Educ. 1998, 26, 320–323. [Google Scholar] [CrossRef]
- Selvin, P.R. The renaissance of fluorescence resonance energy transfer. Nat. Struct. Biol. 2000, 7, 730–734. [Google Scholar] [CrossRef]
- Selvin, P.R. Fluorescence resonance energy transfer. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1995; Volume 246, pp. 300–334. [Google Scholar]
- Szöllosi, J.; Damjanovich, S.; Mátyus, L. Application of fluorescence resonance energy transfer in the clinical laboratory: Routine and research. Cytometry 1998, 34, 159–179. [Google Scholar] [CrossRef]
- Ju Youn, H.; Terpetschnig, E.; Szmacinski, H.; Lakowicz, J.R. Fluorescence Energy Transfer Immunoassay Based on a Long-Lifetime Luminescent Metal–Ligand Complex. Anal. Biochem. 1995, 232, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Censullo, M.L.; Kim, H.G.; Knutson, J.R.; Han, M.K. Characterization of Endonucleolytic Activity of HIV-1 Integrase Using a Fluorogenic Substrate. Anal. Biochem. 1995, 227, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Jovin, T.M.; Arndt-Jovin, D.J. 5—FRET Microscopy: Digital Imaging of Fluorescence Resonance Energy Transfer. Application in Cell Biology. In Cell Structure and Function by Microspectrofluorometry; Kohen, E., Ed.; Academic Press: Cambridge, MA, USA, 1989; pp. 99–117. [Google Scholar]
- Pearce, D.A.; Walkup, G.K.; Imperiali, B. Peptidyl chemosensors incorporating a fret mechanism for detection of Ni(II). Bioorganic Med. Chem. Lett. 1998, 8, 1963–1968. [Google Scholar] [CrossRef]
- Aladekomo, J.; Geddes, C.D.; Rolinski, O.J.; Birch, D.J.S. Metal Ion Detection Using Fluorescence Lifetime Sensors. In Proceedings of the European Quantum Electronics Conference, Glasgow, Scotland, 14 September 1998; p. QTuG61. [Google Scholar]
- Olaf, J.R.; David, J.S.B. A fluorescence lifetime sensor for Cu(I) ions. Meas. Sci. Technol. 1999, 10, 127. [Google Scholar] [CrossRef]
- Brown, M.P.; Royer, C. Fluorescence spectroscopy as a tool to investigate protein interactions. Curr. Opin. Biotechnol. 1997, 8, 45–49. [Google Scholar] [CrossRef]
- Heyduk, T. Measuring protein conformational changes by FRET/LRET. Curr. Opin. Biotechnol. 2002, 13, 292–296. [Google Scholar] [CrossRef]
- Quan, K.; Yi, C.; Yang, X.; He, X.; Huang, J.; Wang, K. FRET-based nucleic acid probes: Basic designs and applications in bioimaging. TrAC Trends Anal. Chem. 2020, 124, 115784. [Google Scholar] [CrossRef]
- Füchtbauer, A.F.; Wranne, M.S.; Bood, M.; Weis, E.; Pfeiffer, P.; Nilsson, J.R.; Dahlén, A.; Grøtli, M.; Wilhelmsson, L.M. Interbase FRET in RNA: From A to Z. Nucleic Acids Res. 2019, 47, 9990–9997. [Google Scholar] [CrossRef] [PubMed]
- Clegg, R.M. Fluorescence resonance energy transfer and nucleic acids. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1992; Volume 211, pp. 353–388. [Google Scholar]
- Imani, M.; Mohajeri, N.; Rastegar, M.; Zarghami, N. Recent advances in FRET-Based biosensors for biomedical applications. Anal. Biochem. 2021, 630, 114323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, Y.; Yang, X.; Tang, Y.; Han, S.; Kang, A.; Deng, H.; Chi, Y.; Zhu, D.; Lu, Y. FÖrster resonance energy transfer (FRET)-based biosensors for biological applications. Biosens. Bioelectron. 2019, 138, 111314. [Google Scholar] [CrossRef] [PubMed]
- Pollok, B.A.; Heim, R. Using GFP in FRET-based applications. Trends Cell Biol. 1999, 9, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.; Enderle, T.; Ogletree, D.F.; Chemla, D.S.; Selvin, P.R.; Weiss, S. Probing the interaction between two single molecules: Fluorescence resonance energy transfer between a single donor and a single acceptor. Proc. Natl. Acad. Sci. USA 1996, 93, 6264–6268. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.C.; Hildebrandt, L.L.; Kragh, S.L.; Birkedal, V. Structural dynamics of nucleic acids by single-molecule FRET. Methods Cell Biol. 2013, 113, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, R.; Solem, A.; Black, W.; Rueda, D. Single-molecule FRET of protein-nucleic acid and protein-protein complexes: Surface passivation and immobilization. Methods 2010, 52, 192–200. [Google Scholar] [CrossRef]
- Kaur, A.; Sapkota, K.; Dhakal, S. Multiplexed Nucleic Acid Sensing with Single-Molecule FRET. ACS Sens. 2019, 4, 623–633. [Google Scholar] [CrossRef]
- Mazal, H.; Haran, G. Single-molecule FRET methods to study the dynamics of proteins at work. Curr. Opin. Biomed. Eng. 2019, 12, 8–17. [Google Scholar] [CrossRef]
- Tan, Y.W.; Hanson, J.A.; Chu, J.W.; Yang, H. Confocal single-molecule FRET for protein conformational dynamics. Methods Mol. Biol. 2014, 1084, 51–62. [Google Scholar] [CrossRef]
- Metskas, L.A.; Rhoades, E. Single-Molecule FRET of Intrinsically Disordered Proteins. Annu. Rev. Phys. Chem. 2020, 71, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Gauer, J.W.; LeBlanc, S.; Hao, P.; Qiu, R.; Case, B.C.; Sakato, M.; Hingorani, M.M.; Erie, D.A.; Weninger, K.R. Single-Molecule FRET to Measure Conformational Dynamics of DNA Mismatch Repair Proteins. Methods Enzymol. 2016, 581, 285–315. [Google Scholar] [CrossRef]
- Wu, S.; Liu, J.; Wang, W. Dissecting the Conformational Dynamics-Modulated Enzyme Catalysis with Single-Molecule FRET. J. Phys. Chem. B 2018, 122, 6179–6187. [Google Scholar] [CrossRef] [PubMed]
- Huynh, M.; Sengupta, B. Analysis of Enzyme Conformation Dynamics Using Single-Molecule Förster Resonance Energy Transfer (smFRET). Biophysica 2022, 2, 123–134. [Google Scholar] [CrossRef]
- Deindl, S.; Zhuang, X. Monitoring conformational dynamics with single-molecule fluorescence energy transfer: Applications in nucleosome remodeling. Methods Enzymol. 2012, 513, 59–86. [Google Scholar] [CrossRef]
- Deichmann, U. Chromatin: Its history, current research, and the seminal researchers and their philosophy. Perspect Biol. Med. 2015, 58, 143–164. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D. Chromatin Structure: A Repeating Unit of Histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef]
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef]
- Kornberg, R.D.; Lorch, Y. Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell 1999, 98, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Sajan, S.A.; Hawkins, R.D. Methods for identifying higher-order chromatin structure. Annu. Rev. Genom. Hum. Genet. 2012, 13, 59–82. [Google Scholar] [CrossRef]
- Zhou, K.; Gaullier, G.; Luger, K. Nucleosome structure and dynamics are coming of age. Nat. Struct. Mol. Biol. 2019, 26, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Mansisidor, A.R.; Risca, V.I. Chromatin accessibility: Methods, mechanisms, and biological insights. Nucleus 2022, 13, 236–276. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, C. Joseph R. Lakowicz: Principles of fluorescence spectroscopy. Anal. Bioanal. Chem. 2008, 390, 1223–1224. [Google Scholar] [CrossRef]
- Hohng, S.; Joo, C.; Ha, T. Single-molecule three-color FRET. Biophys. J. 2004, 87, 1328–1337. [Google Scholar] [CrossRef]
- Yoo, J.; Louis, J.M.; Gopich, I.V.; Chung, H.S. Three-Color Single-Molecule FRET and Fluorescence Lifetime Analysis of Fast Protein Folding. J. Phys. Chem. B 2018, 122, 11702–11720. [Google Scholar] [CrossRef]
- Voith von Voithenberg, L.; Barth, A.; Trauschke, V.; Demarco, B.; Tyagi, S.; Koehler, C.; Lemke, E.A.; Lamb, D.C. Comparative analysis of the coordinated motion of Hsp70s from different organelles observed by single-molecule three-color FRET. Proc. Natl. Acad. Sci. 2021, 118, e2025578118. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Ragunathan, K.; Joo, C.; Ha, T.; Hohng, S. Single-molecule four-color FRET. Angew Chem. Int. Ed. Engl. 2010, 49, 9922–9925. [Google Scholar] [CrossRef]
- Four-color FRET follows Hsp90 states. Nat. Methods 2014, 11, 791. [CrossRef]
- Stein, I.H.; Steinhauer, C.; Tinnefeld, P. Single-Molecule Four-Color FRET Visualizes Energy-Transfer Paths on DNA Origami. J. Am. Chem. Soc. 2011, 133, 4193–4195. [Google Scholar] [CrossRef]
- Li, G.; Levitus, M.; Bustamante, C.; Widom, J. Rapid spontaneous accessibility of nucleosomal DNA. Nat. Struct. Mol. Biol. 2005, 12, 46–53. [Google Scholar] [CrossRef]
- Tomschik, M.; Zheng, H.; van Holde, K.; Zlatanova, J.; Leuba, S.H. Fast, long-range, reversible conformational fluctuations in nucleosomes revealed by single-pair fluorescence resonance energy transfer. Proc. Natl. Acad. Sci. USA 2005, 102, 3278–3283. [Google Scholar] [CrossRef]
- Gansen, A.; Valeri, A.; Hauger, F.; Felekyan, S.; Kalinin, S.; Tóth, K.; Langowski, J.; Seidel, C.A.M. Nucleosome disassembly intermediates characterized by single-molecule FRET. Proc. Natl. Acad. Sci. USA 2009, 106, 15308–15313. [Google Scholar] [CrossRef]
- Koopmans, W.J.A.; Buning, R.; Schmidt, T.; van Noort, J. spFRET Using Alternating Excitation and FCS Reveals Progressive DNA Unwrapping in Nucleosomes. Biophys. J. 2009, 97, 195–204. [Google Scholar] [CrossRef]
- Ngo, T.T.; Ha, T. Nucleosomes undergo slow spontaneous gaping. Nucleic Acids Res. 2015, 43, 3964–3971. [Google Scholar] [CrossRef]
- Böhm, V.; Hieb, A.R.; Andrews, A.J.; Gansen, A.; Rocker, A.; Tóth, K.; Luger, K.; Langowski, J. Nucleosome accessibility governed by the dimer/tetramer interface. Nucleic Acids Res. 2011, 39, 3093–3102. [Google Scholar] [CrossRef]
- Gansen, A.; Hieb, A.R.; Böhm, V.; Tóth, K.; Langowski, J. Closing the gap between single molecule and bulk FRET analysis of nucleosomes. PLoS ONE 2013, 8, e57018. [Google Scholar] [CrossRef]
- Lee, J.; Lee, T.H. Single-Molecule Investigations on Histone H2A-H2B Dynamics in the Nucleosome. Biochemistry 2017, 56, 977–985. [Google Scholar] [CrossRef]
- Kilic, S.; Felekyan, S.; Doroshenko, O.; Boichenko, I.; Dimura, M.; Vardanyan, H.; Bryan, L.C.; Arya, G.; Seidel, C.A.M.; Fierz, B. Single-molecule FRET reveals multiscale chromatin dynamics modulated by HP1α. Nat. Commun. 2018, 9, 235. [Google Scholar] [CrossRef]
- Lehmann, K.; Felekyan, S.; Kühnemuth, R.; Dimura, M.; Tóth, K.; Seidel, C.A.M.; Langowski, J. Dynamics of the nucleosomal histone H3 N-terminal tail revealed by high precision single-molecule FRET. Nucleic Acids Res. 2020, 48, 1551–1571. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Lee, J.; Yue, H.; Lee, T.H. Dynamics of nucleosome assembly and effects of DNA methylation. J. Biol. Chem. 2015, 290, 4291–4303. [Google Scholar] [CrossRef] [PubMed]
- Neumann, H.; Hancock, S.M.; Buning, R.; Routh, A.; Chapman, L.; Somers, J.; Owen-Hughes, T.; van Noort, J.; Rhodes, D.; Chin, J.W. A method for genetically installing site-specific acetylation in recombinant histones defines the effects of H3 K56 acetylation. Mol. Cell 2009, 36, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Zorro Shahidian, L.; Haas, M.; Le Gras, S.; Nitsch, S.; Mourão, A.; Geerlof, A.; Margueron, R.; Michaelis, J.; Daujat, S.; Schneider, R. Succinylation of H3K122 destabilizes nucleosomes and enhances transcription. EMBO Rep. 2021, 22, e51009. [Google Scholar] [CrossRef]
- Sengupta, B.; Huynh, M.; Smith, C.B.; McGinty, R.K.; Krajewski, W.; Lee, T.H. The Effects of Histone H2B Ubiquitylations on the Nucleosome Structure and Internucleosomal Interactions. Biochemistry 2022, 61, 2198–2205. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Wei, S.; Lee, T.H.; Hayes, J.J. Chromatin structure-dependent conformations of the H1 CTD. Nucleic Acids Res. 2016, 44, 9131–9141. [Google Scholar] [CrossRef]
- Blosser, T.R.; Yang, J.G.; Stone, M.D.; Narlikar, G.J.; Zhuang, X. Dynamics of nucleosome remodelling by individual ACF complexes. Nature 2009, 462, 1022–1027. [Google Scholar] [CrossRef]
- Deindl, S.; Hwang, W.L.; Hota, S.K.; Blosser, T.R.; Prasad, P.; Bartholomew, B.; Zhuang, X. ISWI remodelers slide nucleosomes with coordinated multi-base-pair entry steps and single-base-pair exit steps. Cell 2013, 152, 442–452. [Google Scholar] [CrossRef]
- Sabantsev, A.; Levendosky, R.F.; Zhuang, X.; Bowman, G.D.; Deindl, S. Direct observation of coordinated DNA movements on the nucleosome during chromatin remodelling. Nat. Commun. 2019, 10, 1720. [Google Scholar] [CrossRef]
- Harada, B.T.; Hwang, W.L.; Deindl, S.; Chatterjee, N.; Bartholomew, B.; Zhuang, X. Stepwise nucleosome translocation by RSC remodeling complexes. Elife 2016, 5. [Google Scholar] [CrossRef]
- Qiu, Y.; Levendosky, R.F.; Chakravarthy, S.; Patel, A.; Bowman, G.D.; Myong, S. The Chd1 Chromatin Remodeler Shifts Nucleosomal DNA Bidirectionally as a Monomer. Molecular Cell 2017, 68, 76–88.e76. [Google Scholar] [CrossRef] [Green Version]
- Gamarra, N.; Johnson, S.L.; Trnka, M.J.; Burlingame, A.L.; Narlikar, G.J. The nucleosomal acidic patch relieves auto-inhibition by the ISWI remodeler SNF2h. Elife 2018, 7. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Johnson, S.L.; Lee, L.J.; Longhurst, A.D.; Beckwith, S.L.; Johnson, M.J.; Morrison, A.J.; Narlikar, G.J. The Yeast INO80 Complex Operates as a Tunable DNA Length-Sensitive Switch to Regulate Nucleosome Sliding. Mol. Cell 2018, 69, 677–688.e679. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Narlikar, G.J. ATP Hydrolysis Coordinates the Activities of Two Motors in a Dimeric Chromatin Remodeling Enzyme. J. Mol. Biol. 2022, 434, 167653. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Schall, K.; Kallis, E.; Eustermann, S.; Guariento, M.; Moldt, M.; Hopfner, K.P.; Michaelis, J. Single-molecule nucleosome remodeling by INO80 and effects of histone tails. FEBS Lett. 2018, 592, 318–331. [Google Scholar] [CrossRef]
- Poyton, M.F.; Feng, X.A.; Ranjan, A.; Lei, Q.; Wang, F.; Zarb, J.S.; Louder, R.K.; Park, G.; Jo, M.H.; Ye, J.; et al. Coordinated DNA and histone dynamics drive accurate histone H2A.Z exchange. Sci. Adv. 2022, 8, eabj5509. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; North, J.A.; Rose, S.D.; Poirier, M.G. Nucleosomes accelerate transcription factor dissociation. Nucleic Acids Res. 2014, 42, 3017–3027. [Google Scholar] [CrossRef]
- Luo, Y.; North, J.A.; Poirier, M.G. Single molecule fluorescence methodologies for investigating transcription factor binding kinetics to nucleosomes and DNA. Methods 2014, 70, 108–118. [Google Scholar] [CrossRef]
- Crickard, J.B.; Lee, J.; Lee, T.H.; Reese, J.C. The elongation factor Spt4/5 regulates RNA polymerase II transcription through the nucleosome. Nucleic Acids Res. 2017, 45, 6362–6374. [Google Scholar] [CrossRef]
- Senavirathne, G.; Mahto, S.K.; Hanne, J.; O’Brian, D.; Fishel, R. Dynamic unwrapping of nucleosomes by HsRAD51 that includes sliding and rotational motion of histone octamers. Nucleic Acids Res. 2016, 45, 685–698. [Google Scholar] [CrossRef]
- Jang, S.; Kang, C.; Yang, H.S.; Jung, T.; Hebert, H.; Chung, K.Y.; Kim, S.J.; Hohng, S.; Song, J.J. Structural basis of recognition and destabilization of the histone H2B ubiquitinated nucleosome by the DOT1L histone H3 Lys79 methyltransferase. Genes Dev. 2019, 33, 620–625. [Google Scholar] [CrossRef] [Green Version]
- Spangler, C.J.; Yadav, S.P.; Li, D.; Geil, C.N.; Smith, C.B.; Wang, G.G.; Lee, T.-H.; McGinty, R.K. DOT1L activity in leukemia cells requires interaction with ubiquitylated H2B that promotes productive nucleosome binding. Cell Rep. 2022, 38, 110369. [Google Scholar] [CrossRef] [PubMed]
- Mivelaz, M.; Cao, A.M.; Kubik, S.; Zencir, S.; Hovius, R.; Boichenko, I.; Stachowicz, A.M.; Kurat, C.F.; Shore, D.; Fierz, B. Chromatin Fiber Invasion and Nucleosome Displacement by the Rap1 Transcription Factor. Mol. Cell 2020, 77, 488–500.e489. [Google Scholar] [CrossRef]
- Huynh, M.T.; Yadav, S.P.; Reese, J.C.; Lee, T.-H. Nucleosome Dynamics during Transcription Elongation. ACS Chem. Biol. 2020, 15, 3133–3142. [Google Scholar] [CrossRef] [PubMed]
- Safaric, B.; Chacin, E.; Scherr, M.J.; Rajappa, L.; Gebhardt, C.; Kurat, C.F.; Cordes, T.; Duderstadt, K.E. The fork protection complex recruits FACT to reorganize nucleosomes during replication. Nucleic Acids Res. 2022, 50, 1317–1334. [Google Scholar] [CrossRef] [PubMed]
- Tomschik, M.; van Holde, K.; Zlatanova, J. Nucleosome dynamics as studied by single-pair fluorescence resonance energy transfer: A reevaluation. J. Fluoresc. 2009, 19, 53–62. [Google Scholar] [CrossRef]
- Bacic, L.; Sabantsev, A.; Deindl, S. Recent advances in single-molecule fluorescence microscopy render structural biology dynamic. Curr. Opin. Struct Biol. 2020, 65, 61–68. [Google Scholar] [CrossRef]
- Gansen, A.; Felekyan, S.; Kühnemuth, R.; Lehmann, K.; Tóth, K.; Seidel, C.A.M.; Langowski, J. High precision FRET studies reveal reversible transitions in nucleosomes between microseconds and minutes. Nat. Commun. 2018, 9, 4628. [Google Scholar] [CrossRef]
- Llères, D.; James, J.; Swift, S.; Norman, D.G.; Lamond, A.I. Quantitative analysis of chromatin compaction in living cells using FLIM-FRET. J. Cell. Biol. 2009, 187, 481–496. [Google Scholar] [CrossRef]
- Torres, T.; Levitus, M. Measuring conformational dynamics: A new FCS-FRET approach. J. Phys. Chem. B 2007, 111, 7392–7400. [Google Scholar] [CrossRef]
- Ngo, T.T.; Zhang, Q.; Zhou, R.; Yodh, J.G.; Ha, T. Asymmetric unwrapping of nucleosomes under tension directed by DNA local flexibility. Cell 2015, 160, 1135–1144. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.A.; Shundrovsky, A.; Bai, L.; Fulbright, R.M.; Lis, J.T.; Wang, M.D. High-resolution dynamic mapping of histone-DNA interactions in a nucleosome. Nat. Struct Mol. Biol. 2009, 16, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Bustamante, C. Pulling a single chromatin fiber reveals the forces that maintain its higher-order structure. Proc. Natl. Acad. Sci. USA 2000, 97, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Topic | Subtopic | Details | Reference |
|---|---|---|---|
| Chromatin structure and dynamics | Native state | Spontaneous confirmational dynamics between closed and open state | [47,48] |
| Conformational dynamics between three states: closed, partially open, and open | [49] | ||
| Equilibrium constants of DNA unwrapping | [50] | ||
| Conformational switching time is different for different DNA ends | [51] | ||
| Stepwise spontaneous DNA unwrapping resulting in histone eviction | [52] | ||
| Effect of salt | Increasing salt concentration triggers DNA unwrapping | [49] | |
| Salt-induced DNA unwrapping is reversible | [51] | ||
| Nucleosomes containing acetylated histones reflect additional conformational states as compared to native | [53] | ||
| DNA needs to unwrap before dimer eviction | [54] | ||
| Higher-order nucleosome structure | Transient formation of tetrameric structure, interaction of HP1α with methylated histone tails | [55] | |
| Motion of H3 N-terminal tail | [56] | ||
| Effect of DNA modification | CpG-methylation assists nucleosome assembly, formation of stable tetrasomes | [57] | |
| Effect of histone post translational modification | Acetylation results in increased nucleosome breathing | [58] | |
| Succinylation decreases nucleosome stability | [59] | ||
| Ubiquitylation destabilizes nucleosome, facilitates dinucleosome formation | [60] | ||
| Effect of H1 linker histone | H1 influences higher-order nucleosome structure | [61] | |
| Interaction of nucleosome with protein machineries | Interaction with remodelers | Nucleosome remodeling comprises two steps: ACF binding and ACF translocation | [62] |
| Stepwise ISWF-directed translocation of nucleosome | [63] | ||
| Entry and exit side time lag of nucleosomeal DNA translocation during Chd1- and SNF2h-induced remodeling | [64] | ||
| SWI/SNF directed remodeling is bidirectional and sequence independent, and histone octamer is retained with DNA throughout the process | [65] | ||
| Stable remodeled nucleosomes are generated via unstable intermediates for Chd1 controlled remodeling | [66] | ||
| Nucleosomal acidic patch plays crucial role in ISWI and INO80 controlled remodeling. | [67] | ||
| DNA sliding by INO80 depends on the length of flanking DNA | [68] | ||
| Directionality of nucleosome sliding is governed by the coordination of ATP hydrolyzing motors of two units of the dimeric remodeler. | [69] | ||
| Effect of histone tails on the remodeling process | [70] | ||
| Histone dimer exchange happens in seconds timescale. | [71] | ||
| Interaction with other protein machineries | Transcription factors binds to duplex DNA more effectively than mononucleosome or nucleosome array. | [72] | |
| Single-molecule measurements can accurately measure the binding constant for the interaction between the nucleosome and transcription factors | [73] | ||
| Spt4/5 helps polymerase to escape the pause during transcription and facilitates DNA rewrapping behind it | [74] | ||
| Rad51-mediated unwrapping of nucleosomal DNA happens stepwise | [75] | ||
| Dot1L binding destabilizes nucleosomes | [76] | ||
| Dot1L interacts directly with ubiquitin on H3K120, which leads to an effective acidic patch binding | [77] | ||
| Rap1 loosens up chromatin fiber by transient interaction with it | [78] | ||
| Acetylation at K56 of H3 greatly reduces the Pol II pause duration near the entry site of the nucleosome | [79] | ||
| FACT activity is highly dependent on the middle and C terminal domains of the subunits Pob3 and Spt6 | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sengupta, B.; Huynh, M. Contribution of smFRET to Chromatin Research. Biophysica 2023, 3, 93-108. https://doi.org/10.3390/biophysica3010007
Sengupta B, Huynh M. Contribution of smFRET to Chromatin Research. Biophysica. 2023; 3(1):93-108. https://doi.org/10.3390/biophysica3010007
Chicago/Turabian StyleSengupta, Bhaswati, and Mai Huynh. 2023. "Contribution of smFRET to Chromatin Research" Biophysica 3, no. 1: 93-108. https://doi.org/10.3390/biophysica3010007
APA StyleSengupta, B., & Huynh, M. (2023). Contribution of smFRET to Chromatin Research. Biophysica, 3(1), 93-108. https://doi.org/10.3390/biophysica3010007