The Subnival Vegetation of Moquegua, South Peru: Chasmophytes, Grasslands and Cushion Communities



Abstract
:1. Introduction
2. Study Area
3. Methods
3.1. Data Collection
3.2. Environmental Variables
3.3. Data Analysis
3.4. Classification
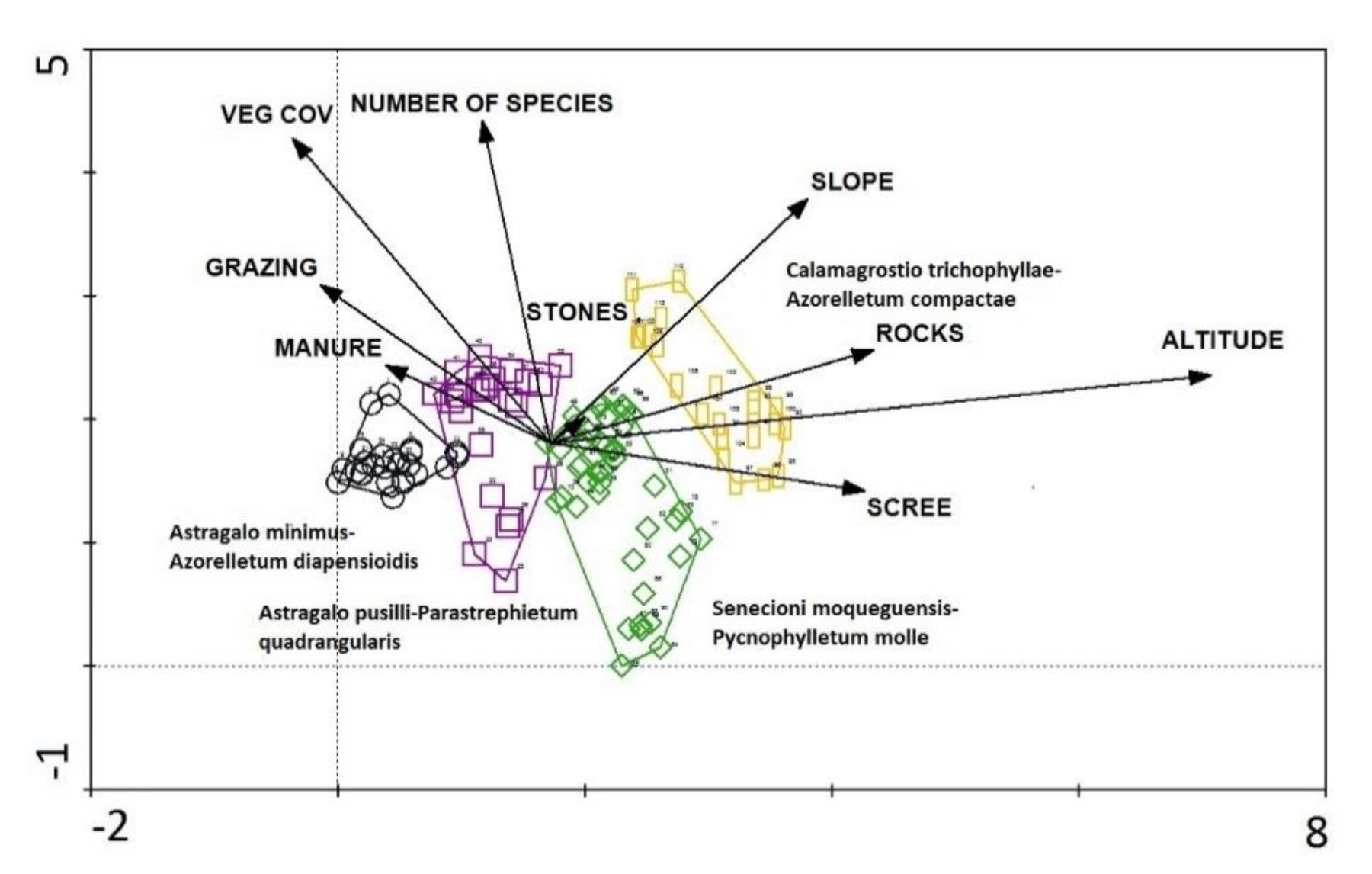
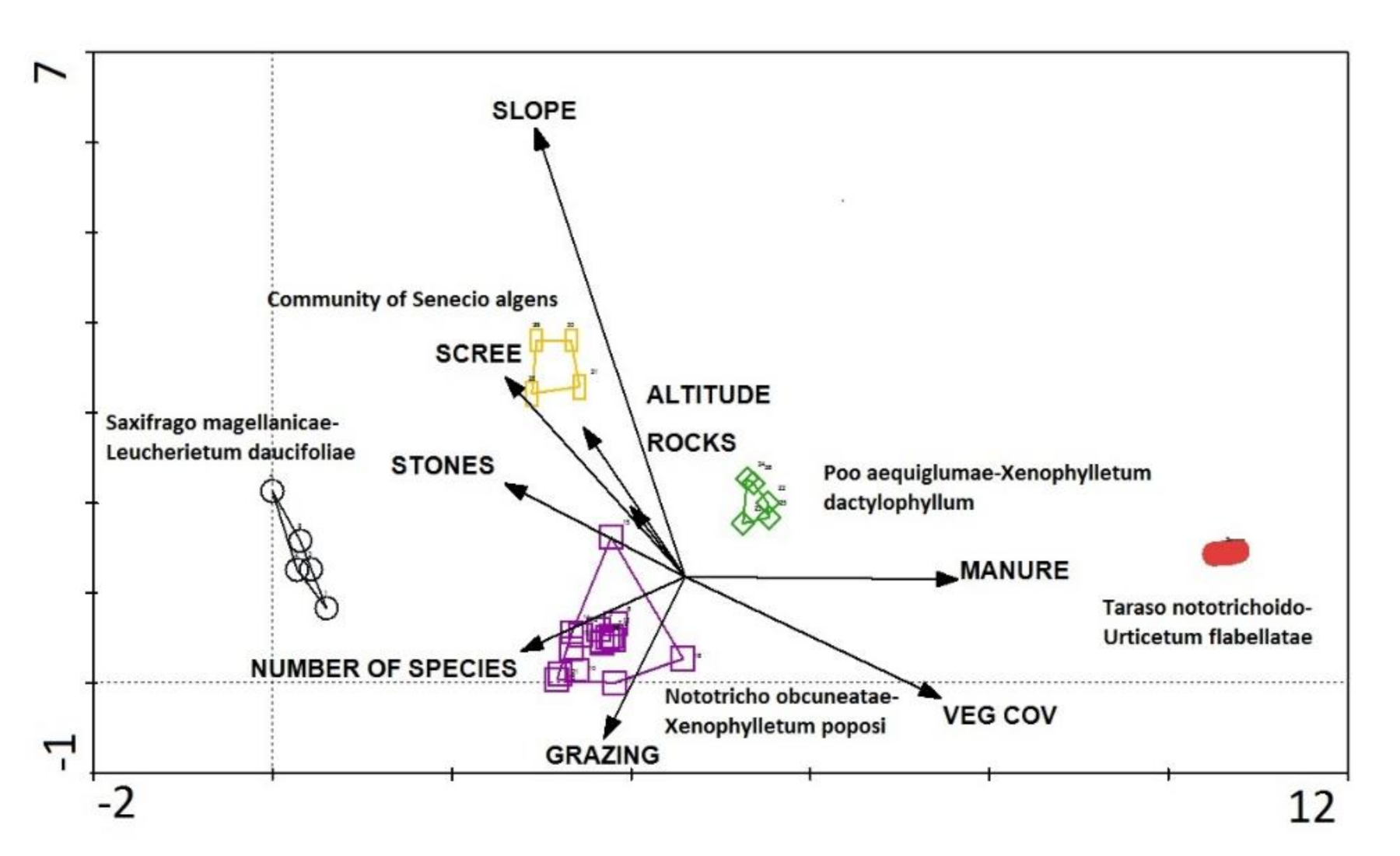
3.5. Gradient Analysis
4. Results
Classification of Relevés and Description of Vegetation Units
5. Community of Senecio algens
6. Gradients and Zonation
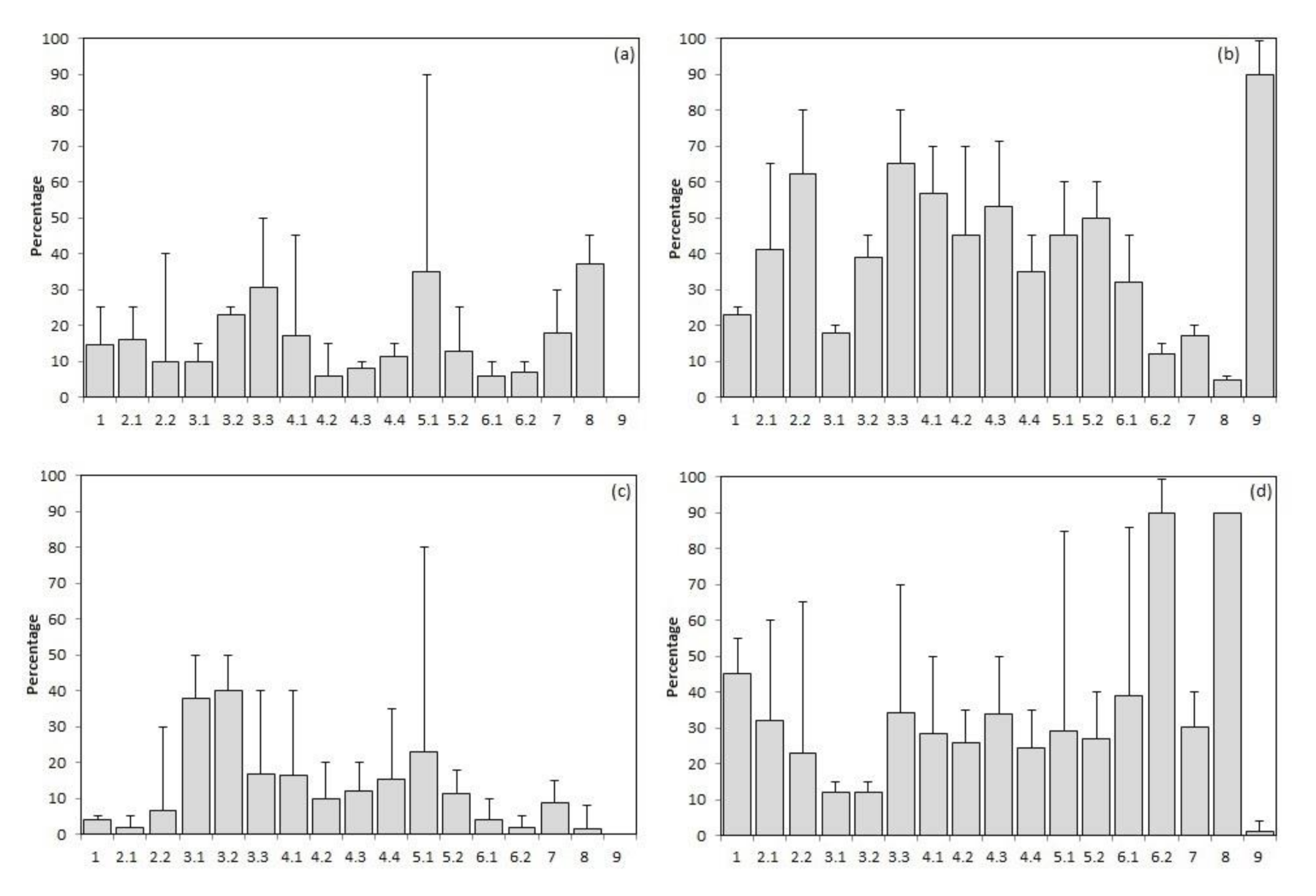
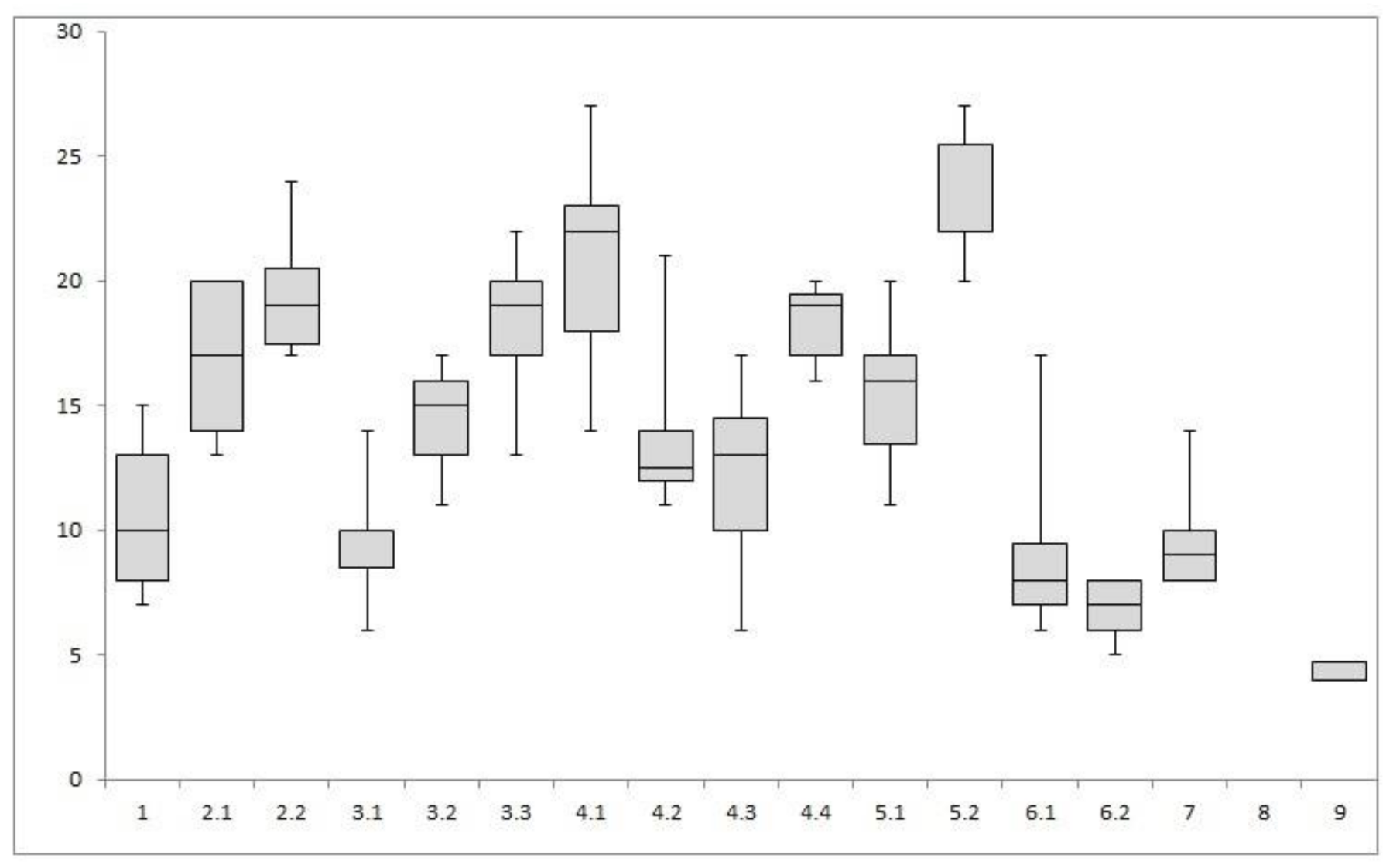
7. Floristic Composition Alfa-Diversity
8. Syntaxonomic Affinity
9. Nitrophilous Vegetation
10. Discussion
11. Conspectus of the Syntaxa
12. Grassland and Cushion Vegetation (Calamagrostietea vicunarum)
13. Cryoturbate Vegetation (Anthochloo lepidulae-Dielsiochloetea Floribundae)
14. Nitrophilous Vegetation
15. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beck, S.; García, E. Flora y Vegetación en los Diferentes Pisos Altitudinales. In Historia Natural de un Valle en Los Andes: La Paz; Forno, E., Baudoin, M., Eds.; Instituto de Ecología, Universidad Mayor de San Andrés: La Paz, Bolivia, 1991; pp. 65–108. [Google Scholar]
- Brack Egg, A.; Mendiola, C. Ecología del Perú; Editorial Bruño: Lima, Perú, 2004; p. 495. [Google Scholar]
- Brako, L.; Zarucchi, J. Catalogue of the Flowering Plants and Gymnosperms of Peru. Monogr. Syst. Bot. Mo. Bot. Gard. 1993, 45, 1–1286. [Google Scholar]
- Braun-Blanquet, J. Plant Sociology, the Study of Plant Communities; McGraw Hill: New York, NY, USA, 1979; 438p. [Google Scholar]
- Cabrera, A.L. Ecología Vegetal de la Puna. Geoecology of the Mountainous Regions of the Tropical Americas. In Colloquium Geographicum 9; Troll, C., Ed.; Dümmlers Verlag: Bonn, Germany, 1968; pp. 91–116. [Google Scholar]
- Cano, A.; Delgado, A.; Mendoza, W.; Trinidad, H.; González, P.; La Torre, M.I.; Chanco, M.; Aponte, H.; Roque, J.; Valencia, N.; et al. Flora y vegetación de suelos crioturbados y hábitats asociados en los alrededores del abra Apacheta, Ayacucho-Huancavelica (Perú). Rev. Peru. Biol. 2011, 18, 169–178. [Google Scholar] [CrossRef]
- Catorci, A.; Piermarteri, K.; Tardella, F.M. Distribution of the nurse species Pycnophyllum molle J. Rémy and P. weberbaueri Muschl. in the Andean dry Puna (Arequipa District-Southern Peru): Role of topographic/soil variability and disturbance regime. Pol. J. Ecol. 2014, 62, 385–390. [Google Scholar] [CrossRef]
- Cleef, A. The Vegetation of the Paramos of the Colombian Cordillera Oriental; Dissertationes Botanicae Series; Lubrecht & Cramer Ltd.: Vaduz, Liechtenstein, 1981; Volume 61, 316p. [Google Scholar]
- de Bello, F.; Doležal, J.; Dvorský, M.; Chlumská, Z.; Řeháková, K.; Klimešová, J.; Klimeš, L. Cushions of Thylacospermum caespitosum (Caryophyllaceae) do not facilitate other plants under extreme altitude and dry conditions in the north-west Himalayas. Ann. Bot. 2011, 108, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Estudio de Impacto Ambiental Semidetallado. Proyecto de Exploración “Chucapaca” EIAS-PEC; Compañia de Minas Buenaventura: Lima, Peru, 2009; pp. 1–32. [Google Scholar]
- Freire, S.E.; Chemisquy, M.A.; Anderberg, A.A.; Beck, S.G.; Meneses, R.L.; Loeuille, B.; Urtubey, E. The Lucilia group (Asteraceae, Gnaphalieae): Phylogenetic and taxonomic considerations based on molecular and morphological evidence. Plant Syst. Evol. 2015, 301, 1227–1248. [Google Scholar] [CrossRef]
- Funk, V.A. Xenophyllum, a new Andean genus extracted from Werneria s.l. (Compositae: Senecioneae). Novon 1997, 7, 235–241. [Google Scholar] [CrossRef]
- Galán de Mera, A. Clasificación fitosociológica de la vegetación de la región del Caribe y América del Sur. Arnaldoa 2005, 12, 86–111. [Google Scholar]
- Galán de Mera, A.; Rosa, M.; Cáceres, C. Una aproximación sintaxonómica sobre la vegetación del Perú. Clases, órdenes y alianzas. Acta Bot. Malacit. 2002, 27, 75–103. [Google Scholar] [CrossRef] [Green Version]
- Galán de Mera, A.; Cáceres, C.; González, A. La vegetación de la alta montaña andina del sur de Perú. Acta Bot. Malacit. 2003, 28, 121–147. [Google Scholar] [CrossRef]
- Galán de Mera, A.; Baldeón, S.; Beltrán, H.; Benavente, M.; Gómez, J. Datos sobre la vegetación del centro del Perú. Acta Bot. Malacit. 2004, 29, 89–115. [Google Scholar] [CrossRef] [Green Version]
- Galán de Mera, A.; Perea, E.L.; de la Cruz, J.C.; Orellana, J.V. Nuevas observaciones sobre la vegetación del sur del Perú. Del desierto pacífico al altiplano. Acta Bot. Malacit. 2009, 34, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Galán de Mera, A.; Perea, E.L.; de la Cruz, J.C.; Vera, C.T.; Villasante-Benavides, F.; Orellana, J.V. Novedades sobre la vegetación del departamento de Arequipa (Perú). Arnaldoa 2011, 18, 125–144. [Google Scholar]
- Galán de Mera, A.; del Monte, B.; Linares Perea, E.; de la Cruz, J.C.; Trujillo Vera, F.; Vicente Orellana, J.A. Las comunidades vegetales relacionadas con los ambientes humanos en el sur del Perú. Phytocoenologia 2012, 41, 265–305. [Google Scholar] [CrossRef]
- Galán de Mera, A.; Méndez, E.; Linares, E.; de la Cruz, C.; Vicente Orellana, J.A. Las comunidades vegetales relacionadas con los procesos criogénicos en los Andes peruanos. Phytocoenologia 2014, 44, 121–161. [Google Scholar] [CrossRef]
- García, E.; Beck, S.G. Puna. In Botánica Económica de los Andes Centrales; Moraes, M., Øllgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; 557p. [Google Scholar]
- Gonzáles, P.; Cano, A.; Müller, J. An unusual new record of Baccharis (Asteraceae) from the Peruvian Andes and its relation with the northern limit of the dry puna. Acta Botánica Mex. 2019, 126. [Google Scholar] [CrossRef] [Green Version]
- Gutte, P. Beitrag zur Kenntnis zentralperuanischer Pflanzengesellschaften IV. Die grasreiche Vegetation der alpine Stufe. Wissenschaftliche Zeitschrift Karl-Marx-Universität Leipzig. Math. Nat. Reihe 1985, 34, 357–401. [Google Scholar]
- Gutte, P. Beitrag zur Kenntnis zentralperuanischer Pflanzengesellschaften III. Pflanzengesellschaften der subalpinen Stufe. Feddes Repert. 1986, 97, 319–371. [Google Scholar] [CrossRef]
- Gutte, P. Beitrag zur Kenntnis zentralperuanischer Pflanzengesellschaften V. Die Vegetation der subnivalen Stufe. Feddes Repert. 1987, 98, 447–460. [Google Scholar] [CrossRef]
- Gutte, P. Segetal- und Ruderalpflanzengesellschaften im Wohngebiet der Kallawaya (Bolivianische Anden). Phytocoenologia 1995, 25, 33–67. [Google Scholar] [CrossRef]
- Hedberg, O. Evolution and speciation in a tropical high mountain flora. Biol. J. Linn. Soc. 1969, 1, 135–148. [Google Scholar] [CrossRef]
- Hill, M.O. Twinspan, a Fortran Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and the Attributes; Cornell University, Department of Ecology and Systematics: Ithaca, NY, USA, 1979. [Google Scholar]
- Holmgren, C.A.; Betancourt, J.L.; Rylander, K.A.; Roque, J.; Tovar, O.; Zeballos, H.; Linares, E.; Quade, J. Holocene vegetation history from fossil rodent middens near Arequipa, Peru. Quat. Res. 2001, 56, 242–251. [Google Scholar] [CrossRef]
- Josse, C.; Cuesta, F.; Navarro, G.; Barrena, V.; Becerra, M.T.; Cabrera, E.; Chacón-Moreno, E.; Ferreira, W.; Peralvo, M.; Saito, J.; et al. Physical Geography and Ecosystems in the Tropical Andes. In Climate Change and Biodiversity in the Tropical Andes; Herzog, S.K., Martinez, R., Jørgensen, P.M., Tiessen, H., Eds.; MacArthur Foundation: Chicago, IL, USA; IAI: San Jose dos Campos, Brazil; SCOPE: Paris, France, 2011; Chapter 10; pp. 152–169. [Google Scholar]
- JSTOR Global Plants. Available online: https://plants.jstor.org/ (accessed on 26 December 2020).
- Kleier, C.; Trenary, T.; Graham, E.A.; Stenzel, W.; Rundel, P.W. Size class structure, growth rates, and orientation of the central Andean cushion Azorella compacta. PeerJ 2015, 3, e843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, R. Considerations on Quantitative Parameters and Qualitative Attributes in Vegetation Analysis and in Phytosociological Relevés. In Sampling Methods and Taxon Analysis in Vegetation Science; Knapp, R., Ed.; Dr. W. Junk Publishers: Heidelberg, The Netherlands; Boston, MS, USA; Lancaster, UK, 1984; pp. 77–100. [Google Scholar]
- Kuentz, A.; Galán de Mera, A.; Ledru, M.P.; Thouret, J.C. Phytogeographical data and modern pollen rain of the Puna belt in southern Peru (Nevado Coropuna, Western Cordillera). J. Biogeogr. 2007, 34, 1762–1776. [Google Scholar] [CrossRef]
- Lauer, W. Zur hygrischen Höhenstufung tropischer Gebirge. Neotropische Ökosysteme. Biogeographica 1976, 7, 169–182. [Google Scholar]
- Luebert, F.; Gajardo, R. Vegetación de los Andes áridos del norte de Chile. Lazaroa 2000, 21, 111–130. [Google Scholar]
- Luebert, F.; Gajardo, R. Vegetación altoandina de Parinacota (norte de Chile) y una sinopsis de la vegetación de la puna meridional. Phytocoenologia 2005, 35, 79–128. [Google Scholar] [CrossRef]
- Martinez, S. El género Azorella (Apiaceae-Hydrocotyloideae) en la Argentina. Darwiniana 1989, 29, 139–178. [Google Scholar]
- Mendez, E. La Vegetación de los Altos Andes Centrales: Bardas Blancas-Paso Pehuenche (Malargüe, Mendoza, Argentina). Boletín Soc. Argent. Botánica 2014, 49, 257–281. [Google Scholar] [CrossRef]
- Montenegro, B.; Zúñiga, S.; Zeballos, H. Climatología de la Reserva Nacional Salinas y Aguada Blanca, Suroeste del Perú. In Diversidad Biológica de la Reserva Nacional de Salinas y Aguada Blanca; Zeballos, H., Ochoa, J.A., López, E., Eds.; LITHO & ARTE S.A.C: Lima, Peru, 2010; pp. 261–273. [Google Scholar]
- Montesinos, D.B.; Cleef, A.M.; Sýkora, K.V. Andean shrublands of Moquegua, South Peru: Prepuna plant communities. Phytocoenologia 2012, 42, 29–55. [Google Scholar] [CrossRef]
- Montesinos-Tubée, D.B. Diversidad florística de la cuenca alta del río Tambo-Ichuña (Moquegua, Perú). Rev. Peru. Biol. 2011, 18, 119–132. [Google Scholar] [CrossRef]
- Montesinos-Tubée, D.B. Lista anotada de nuevas adiciones para la flora andina de Moquegua, Perú. Rev. Peru. Biol. 2012, 19, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Montesinos-Tubée, D.B. Flora de los Andes de Moquegua. Etnobotánica de la Cuenca de los ríos Alto Tambo-Ichuña; CDH-Gold Fields. DL. 2013–06310; Inca Legacy Cultural Society: Lima, Peru, 2013; pp. 1–300. [Google Scholar]
- Montesinos-Tubée, D.B. Three new caespitose species of Senecio (Asteraceae: Senecioneae) from South Peru. Phytokeys 2014, 39, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Montesinos-Tubée, D.B. Pycnophyllum molle and its tenants in southern Peru. J. Alp. Gard. Soc. 2015, 82, 438–443. [Google Scholar]
- Montesinos-Tubée, D.B. Flora Moqueguana. Guía Práctica para la Identificación de Flora Silvestre; Anglo American: Lima, Perú, 2015; 252p. [Google Scholar]
- Montesinos-Tubée, D.B.; Kool, A. Arenaria acaulis (Caryophyllaceae): A new species from South Peru. Phytotaxa 2015, 220, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Montesinos-Tubée, D.B.; Sýkora, K.V.; Quipuscoa-Silvestre, V.; Cleef, A.M. Species composition and phytosociology of xerophytic plant communities after extreme rainfall in South Peru. Phytocoenologia 2015, 48, 203–250. [Google Scholar] [CrossRef]
- Montesinos-Tubée, D.B.; Cleef, A.M.; Sýkora, K.V. The Puna vegetation of Moquegua, South Peru: Chasmophytes, grasslands and Puya raimondii stands. Phytocoenologia 2015, 45, 365–397. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: Hoboken, NJ, USA, 1974. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Navarro, G. Vegetación de Bolivia: El Altiplano meridional. Rivasgodaya 1993, 7, 69–98. [Google Scholar]
- Navarro, G.; Ferreira, W. Zonas de vegetación potencial de Bolivia: Una base para el análisis de vacíos de conservación. Rev. Boliv. Ecol. 2004, 15, 1–40. [Google Scholar]
- Navarro, G.; Maldonado, M. Geografía Ecológica de Bolivia. Vegetación y Ambientes Acuáticos; Fundación Simón 1 ; Patiño: Santa Cruz, Bolivia, 2005; pp. 247–268, 454–499. [Google Scholar]
- PBI Solanum Project. Solanaceae Source. Available online: http://www.solanaceaesource.org (accessed on 30 November 2014).
- Pugnaire, F.I.; Morillo, J.A.; Armas, C.; Rodríguez-Echeverría, S.; Gaxiola, A. Azorella compacta: Survival champions in extreme, high-elevation environments. Ecosphere 2020, 11, e03031. [Google Scholar] [CrossRef] [Green Version]
- Quipuscoa-Silvestre, V.; Dillon, M. Tres nuevas especies de Mniodes a. Gray (Gnaphaliinae, Gnaphalieae, Asteraceae) de Bolivia y Perú y cambios nomenclaturales en el grupo Lucilia. Arnaldoa 2020, 27, 2. [Google Scholar]
- Ralph, C.P. Observations on Azorella compacta (Umbelliferae), a tropical Andean cushion plant. Biotropica 1978, 10, 62–67. [Google Scholar] [CrossRef]
- Reese, C.A.; Liu, K.B. Pollen dispersal and deposition on the Quelccaya Ice Cap, Peru. Phys. Geogr. 2002, 23, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Reese, C.A.; Liu, K.B. A modern pollen rain study from the central Andes region of South America. J. Biogeogr. 2005, 32, 709–718. [Google Scholar] [CrossRef]
- Rivas-Martínez, S. Worldwide Bioclimatic Classification System. Phytosociological Research Center. 2004. Available online: www.globalbioclimatic.org (accessed on 12 December 2010).
- Rivas-Martínez, S.; Tovar, O. Vegetatio Andinae, I. Datos sobre las comunidades vegetales altoandinas de los Andes Centrales del Perú. Lazaroa 1982, 4, 167–187. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=905920.
- Rivera, M.; Thouret, J.C.; Samaniego, P.; Le Pennec, J.L. The 2006–2009 activity of the Ubinas volcano (Peru): Petrology of the 2006 eruptive products and insights into genesis of andesite magmas, magma recharge and plumbing system. J. Volcanol. Geotherm. Res. 2014, 270, 122–141. [Google Scholar] [CrossRef]
- Ruthsatz, B. Pflanzengesellschaften und ihre Lebensbedingungen in den Andinen Halbwüsten Nordwest-Argentiniens. Diss. Bot. 1977, 39, 1–90. [Google Scholar]
- Sarmiento, G. Ecological Features of Climate in High Tropical Mountains. In High Altitude Tropical Biogeography; Monasterio, M., Ed.; Oxford University Press: Oxford, UK, 1986; pp. 11–45. [Google Scholar]
- Seibert, P.; Menhofer, X. Die Vegetation des Wohngebietes der Kallawaya und des Hochlandes von Ulla-Ulla in den bolivianischen Anden. Phytocoenologia 1991, 20, 145–276. [Google Scholar] [CrossRef]
- Seibert, P.; Menhofer, X. Die Vegetation des Wohngebietes der Kallawaya und des Hochlandes von Ulla-Ulla in den bolivianischen Anden. Phytocoenologia 1992, 20, 289–438. [Google Scholar] [CrossRef]
- Seibert, P.; Menhofer, X. Die Vegetation des Wohngebietes der Kallawaya und des Hochlandes von Ulla-Ulla in den bolivianischen Anden. Phytocoenologia 1993, 22, 275–278. [Google Scholar] [CrossRef]
- Servicio Nacional de Meteorología e Hidrología del Perú (SENAMHI). Información Meteorológica de las Estaciones de Ichuña y Ubinas (Moquegua); Servicio Nacional de Meteorología e Hidrología del Perú: Arequipa, Peru, 2013.
- Talavera, C.; Ortega, A.; Villegas, L. Flora y Vegetación de la Reserva Nacional de Salinas y Aguada Blanca, Perú. In Diversidad Biológica de la Reserva Nacional de Salinas y Aguada Blanca (Arequipa-Moquegua); Zeballos, H., Ochoa, J.A., López, E., Eds.; DESCO, PROFONANPE, SERNANP: Lima, Peru, 2010. [Google Scholar]
- Teillier, S. Flora y vegetación alto-andina del área de Collaguasi-Salar de Coposa, Andes del norte de Chile. Rev. Chil. Hist. Nat. 1998, 71, 313–329. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P.; CANOCO. Reference manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- NYBG STEERE HERBARIUM. Index Herbariorum. Available online: http://sweetgum.nybg.org/ih/ (accessed on 15 January 2019).
- Troll, C. Geo-Ecology of the Mountainous Regions of the Tropical Americas; Dümmlers Verlag: Bonn, Germany, 1968. [Google Scholar]
- Tropicos. Missouri Botanical Garden. Available online: http://www.tropicos.org (accessed on 1 January 2015).
- Weber, H.E.; Moravec, J.; Theurillat, J.P. International Code of Phytosociological Nomenclature. 3rd edition. J. Veg. Sci. 2000, 11, 739–768. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Locality | District | # rel. | Year | Elevation (m) | Slope (°) | Aspect | Environment | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Janchata slopes | Nevados | Carumas | 5 | 2015 | 4700–4710 | ′5–10 | SE | Scree plateaus |
| 2 | Cerro Santo Tomas, C. Huarmaca, Puqa Saya | Yanahuara | Ichuña | 15 | 2009, 2012 | 4530–4665 | 0–60 | N, NW, S, SE | Chasmophyte, scree slopes with copper, steppe grasslands with cushions, rocky slopes, open plateaus, manure communities |
| 3 | Coriri, Jatun Puqio, Qhaqhaskinkri | Ichuña | Ichuña | 9 | 2009 | 4460–4540 | ′3–43 | NW, S, SE, E, W | Steppe grasslands with cushions, grazed grasslands |
| 4 | Choco-Choco environs | Yunga | Yunga | 10 | 2012 | 4660–4760 | ′2–35 | N, W | Grasslands with cushions, scree slopes, open plateaus with cushions |
| 5 | Sura, Perusa-Chaquisura, Siliaca, Yanasaya | Yunga | Yunga | 28 | 2011, 2012 | 4510–4800 | ′10–50 | NW, N, SSW, SW, W | Chasmophyte, rocky slopes, steppe grasslands with cushions, grasslands, scree, open plateaus with cushions |
| 6 | Rancho-Pirhuani; Laguna Jallpacocha | Tassa/Pachamayo | Ubinas | 18 | 2014 | 4450–4565 | ′5–45 | NE, SE, E | Steppe grasslands with cushions, grazed grasslands, rocky slopes, scree slopes |
| 7 | Pirhuani | Tassa | Ubinas | 18 | 2011 | 4470–4660 | 0–17 | NNW, SSE, W | Steppe cushion communities |
| 8 | Cochapata lake environs | Tassa | Ubinas | 10 | 2013 | 4680–4700 | ′4–5 | SE | Steppe grasslands with cushions |
| 9 | Pacosani, Yaretaq, Larsepesca | Coalaque | Ubinas | 10 | 2009 | 4615–4690 | ′5–47 | N, NE, S, SW | Steppe grasslands with cushions, rock slopes |
| 10 | Janaqpampa, Gasawasi, Witopata | Querala | Ubinas | 10 | 2011 | 4500–4640 | ′4–15 | S, E, W | Steppe grasslands with cushions, open plateaus |
| 11 | Condor Sallana | Matazo | Ubinas | 20 | 2013, 2014 | 4460–4590 | 0–25 | N, S | Grazed grasslands with cushions, manure communities, rocky slopes |
| Community # | 1 | 2.1 | 2.2 | 3.1 | 3.2 | 3.3 | 4.1 | 4.2 | 4.3 | 4.4 | 5.1 | 5.2 | 6.1 | 6.2 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Relevé # | 2 | 6 | 20 | 3 | 8 | 20 | 7 | 17 | 33 | 41 | 14 | 20 | 6 | 14 | 2 | 2 | 8 |
| ALTITUDE | 4580 | 4585 | 4470 | 4450 | 4510 | 4500 | 4670 | 4610 | 4680 | 4590 | 4800 | 4590 | 4650 | 4710 | 4790 | 4745 | 4460 |
| INCLINATION | 25 | 5 | 5 | 5 | 20 | 45 | 10 | 5 | 5 | 10 | 10 | 25 | 5 | 10 | 15 | 45 | 0 |
| ORIENTATION | S | N | W | NE | N | NW | SW | W | SE | N | W | S | SSE | SE | SSW | NW | - |
| # SPP | 8 | 18 | 22 | 8 | 15 | 22 | 18 | 12 | 6 | 20 | 16 | 22 | 8 | 8 | 14 | 6 | 5 |
| VEGC % | 20 | 55 | 45 | 20 | 35 | 60 | 50 | 30 | 35 | 35 | 40 | 45 | 30 | 15 | 15 | 5 | 80 |
| ROCKS % | 5 | 5 | 10 | 40 | 40 | 10 | 0 | 20 | 10 | 5 | 20 | 10 | 0 | 5 | 8 | 8 | 0 |
| STONES % | 55 | 5 | 20 | 10 | 10 | 30 | 35 | 35 | 30 | 30 | 40 | 24 | 30 | 90 | 34 | 90 | 0 |
| GRAZING (I-III) | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 3 |
| Argyrochosmetea niveae | |||||||||||||||||
| Saxifragetalia magellanicae, Saxifragion magellanicae | |||||||||||||||||
| Saxifraga magellanica | 4 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Saxifrago magellanicae-Leucherietum daucifoliae | |||||||||||||||||
| Leucheria daucifolia | 4 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Weberbauera arequipa | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Calamagrostietea vicunarum | |||||||||||||||||
| Calamagrostis vicunarum | . | . | 1 | 1 | . | . | . | 2 | . | 1 | . | . | . | . | . | . | . |
| Parastrephietalia quadrangularis | |||||||||||||||||
| Parastrephia quadrangularis | . | . | 4 | 4 | 4 | 8 | . | . | . | . | . | . | . | . | . | . | . |
| Tetraglochin cristatum | . | 8 | 3 | . | . | 8 | . | 2 | . | . | . | . | . | . | . | . | . |
| Descurainia depressa | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Pycnophyllum molle | . | 10 | 4 | . | . | . | 20 | 10 | 15 | 7 | 8 | . | 8 | . | . | . | . |
| Hypochaeris meyeniana | . | . | 2 | . | 1 | 2 | . | . | . | . | . | . | . | . | . | . | . |
| Ephedra rupestris | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . | . | . |
| Weberbauera spathulifolia | 2 | . | 3 | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Belloa piptolepis | . | . | . | . | 1 | 1 | 1 | 2 | . | . | . | . | . | . | . | . | . |
| Senecio candollei | . | . | . | . | . | . | . | . | . | 1 | 2 | . | . | . | . | . | . |
| Senecio humillimus | . | . | . | . | 2 | . | . | . | . | . | . | 4 | . | . | . | . | . |
| Brayopsis calycina | . | 1 | 1 | . | . | . | . | . | . | 1 | 1 | . | . | . | . | . | . |
| Chaetanthera stuebelii | . | . | . | . | . | . | 1 | . | . | 1 | . | . | . | . | . | . | . |
| Nototriche anthemidifolia | . | 1 | . | . | . | . | 2 | 1 | . | 1 | 1 | . | . | . | . | . | . |
| Draba macleanii | . | 2 | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . |
| Hypochaeris eriolaena | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Plantago sericea var. lanuginosa | . | . | 1 | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Werneria aretioides | . | . | 2 | . | . | . | . | 2 | 2 | . | . | . | . | . | . | . | . |
| Senecio spinosus | . | 3 | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Werneria pectinata | . | . | . | . | 1 | . | 1 | . | . | 2 | . | . | . | . | . | . | . |
| Wahlenbergia peruviana | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Nototriche digitulifolia | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Nototriche pedicularifolia | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Lupinus ananeanus | . | . | 3 | . | . | 3 | . | . | . | . | . | . | . | . | . | . | . |
| Cerastium subspicatum | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Conyza deserticola | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Gomphrena meyeniana | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Calamagrostietea minimae | |||||||||||||||||
| Calamagrostis minima | . | 1 | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Oreomyrrhis andicola | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |
| Werneria nubigena | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Astragalo minimi-Azorelletum diapensioidis | |||||||||||||||||
| Azorella diapensioides | . | 6 | 4 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Astragalus minimus | . | 10 | 2 | . | 2 | 2 | 1 | . | . | . | . | . | . | . | . | . | . |
| Bougueria nubicola | . | 2 | 1 | . | . | 1 | 1 | . | . | . | . | . | . | . | . | . | . |
| Werneria apiculata | . | 2 | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Junellia minima | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Subassociation Typicum | |||||||||||||||||
| Werneria melanandra | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Nototriche longirostris | . | 2 | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| aciachnetosum pulvinatae | |||||||||||||||||
| Aciachne pulvinada | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Azorello-Festucion | |||||||||||||||||
| Azorella compacta | . | . | . | 8 | 10 | 15 | . | . | 12 | 4 | 4 | 4 | . | . | . | . | . |
| Festuca orthophylla | . | . | . | . | . | 2 | 5 | . | . | . | . | . | . | . | . | . | . |
| Baccharis caespitosa | . | . | . | . | 1 | . | . | . | . | . | . | 2 | . | . | . | . | . |
| Belloa sp. (# 3945) | . | . | . | . | . | 1 | . | . | . | . | 1 | . | . | . | . | . | . |
| Bowlesia tropaeolifolia | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Cumulopuntia boliviana subsp. ignescens | . | . | . | . | 3 | . | . | . | . | . | . | . | . | . | . | . | . |
| Senecio evacoides | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . |
| Stangea wandae | . | . | . | . | . | . | . | 1 | . | 1 | . | . | . | . | . | . | . |
| Astragalo pusillii-Parastrephietum quadrangularis | |||||||||||||||||
| Astragalus pusillus | . | . | . | 1 | 1 | 2 | . | . | . | . | . | . | . | . | . | . | . |
| sisyrinchietosum trinervis | |||||||||||||||||
| Sisyrinchium trinervis | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . |
| baccharidetosum tricuneatae | |||||||||||||||||
| Baccharis tricuneata | . | . | . | . | 2 | . | . | . | . | . | 3 | 4 | . | . | . | . | . |
| Descurainia sp. (# 0940) | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . |
| Subassociation Typicum | |||||||||||||||||
| Sisyrinchium brevipes | . | 1 | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Senecioni moqueguensis-Pycnophylletum molle | |||||||||||||||||
| Senecio moqueguensis | . | . | . | . | . | . | 3 | 3 | 4 | 4 | . | . | . | . | . | . | . |
| Nototriche argentea | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . |
| Perezia coerulescens var. amplibracteata | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Subassociation Typicum | |||||||||||||||||
| Mniodes sp. (# 2477) | . | . | . | . | . | . | 3 | . | . | . | . | 2 | . | . | . | . | . |
| Senecio helianthemoides | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . |
| senecionetosum tassaensis | |||||||||||||||||
| Senecio tassaensis | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . |
| Xenophyllum weddellii | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . |
| gentianelletosum primuloides | |||||||||||||||||
| Gentianella primuloides | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . |
| Senecio graveolens | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . |
| arenarietosum acaulis | |||||||||||||||||
| Arenaria acaulis | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Nototriche sepaliloba | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Cerastium behmianum | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Nototriche pusilla | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Senecio sykorae | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Poa brevis | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Calamagrostio trichophyllae-Azorelletum compactae | |||||||||||||||||
| Calamagrostis trichophylla | . | . | . | . | . | . | . | . | . | . | 1 | 2 | . | . | . | . | . |
| Nototriche mandoniana | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . |
| Silene mandonii | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Perezia pungens | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . |
| Subassociation Typicum | |||||||||||||||||
| Mniodes caespititia | . | . | . | . | . | . | . | . | . | . | 5 | . | . | . | . | . | . |
| Senecio sp.2 (# 3935) | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . | . | . | . |
| drabetosum soratensis | |||||||||||||||||
| Draba soratensis | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . |
| Bartsia sp. (# 3092) | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . |
| Poa gilgiana | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Agrostis breviculmis | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Viola granulosa | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Anthochloo-Dielsiochloetea | |||||||||||||||||
| Anthochloa lepidula | 2 | . | . | . | . | . | 1 | . | . | . | . | . | 1 | 1 | 2 | 1 | . |
| Dielsiochloa floribunda | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . |
| Nototrichion obcuneatae | |||||||||||||||||
| Nototriche obcuneata | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | 2 | . | . |
| Senecio adenophyllus | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | 1 | . |
| Senecio sp.3 (# 3931) | . | . | . | . | . | . | . | . | . | . | . | . | 1 | 1 | 2 | . | . |
| Dissanthelium calycinum | . | 1 | . | . | . | 1 | . | . | . | 1 | . | . | . | . | 1 | . | 3 |
| Nototriche sp.1 (# 3104) | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . |
| Nototricho obcuneatae-Xenophylletum poposi Galán de Mera et al. 2003 | |||||||||||||||||
| Xenophyllum poposum | . | . | . | . | . | . | . | . | . | . | . | . | 7 | 3 | . | . | . |
| Mniodes coarctata | . | . | . | . | . | . | . | . | . | 2 | . | . | 7 | 3 | . | . | . |
| nototrichietosum erinaceae | |||||||||||||||||
| Nototriche erinacea | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . |
| senecionetosum trifurcifolii | |||||||||||||||||
| Senecio trifurcifolius | . | . | . | . | . | . | . | . | . | . | . | . | . | 3 | . | . | . |
| Senecio sp.4 (# 4228b) | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . |
| Poo aequiglumae-Xenophylletum dactylophyllum | |||||||||||||||||
| Xenophyllum dactylophyllum | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 6 | . | . |
| Nototriche sp.3 (# 2447) | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . |
| Poa aequigluma | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 | . | . |
| Poa spicigera | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . | . |
| Community of Senecio algens | |||||||||||||||||
| Senecio algens | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . |
| Senecio sp.5 (# 3942) | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . |
| Class ¿? | |||||||||||||||||
| Community of Tarasa nototrichoides and Urtica flabellata | |||||||||||||||||
| Urtica flabellata | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 50 |
| Tarasa nototrichoides | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 10 |
| Lachemilla pinnata | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 10 |
| Perezia multiflora | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | 5 |
| Solanum acaule | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 2 |
| Crassuletea connatae | |||||||||||||||||
| Muhlenbergia peruviana | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Salpichroetalia glandulosae; Argyrochosmetea niveae | |||||||||||||||||
| Valeriana nivalis | . | . | . | . | . | . | . | . | . | . | 2 | . | . | . | . | . | . |
| Companions | |||||||||||||||||
| Bartsia diffusa | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Calamagrostis curvula | . | . | . | . | . | . | . | . | . | . | 1 | . | 1 | . | 2 | 1 | . |
| Cardionema ramosissimum | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . |
| Galium corymbosum | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . |
| Geranium sessiliflorum | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . |
| Lepidium meyenii | . | . | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . |
| Luzula vulcanica | . | . | 1 | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Mancoa hispida | . | . | . | . | . | 2 | . | . | . | . | . | . | . | . | . | . | . |
| Microsteris gracilis | 1 | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Oxalis nubigena | . | . | . | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . |
| Parastrephia lucida | . | . | . | . | . | . | 5 | . | . | 2 | 4 | 4 | . | . | 4 | . | . |
| Paronychia andina | . | . | 1 | . | 1 | . | . | . | . | . | . | . | . | . | . | . | . |
| Poa candamoana | 2 | . | . | . | . | . | . | . | . | . | . | 1 | . | . | . | . | . |
| Pycnophyllum glomeratum | . | . | . | . | . | . | . | 2 | . | . | . | 2 | . | . | 4 | . | . |
| Senecio nutans | . | . | . | . | . | 2 | . | . | . | . | 1 | . | . | . | 3 | . | . |
| Stangea rhizanta | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | 1 | . |
| Stipa ichu | 3 | . | 3 | 3 | 3 | 3 | . | . | . | . | . | . | . | . | . | . | . |
| Trisetum spicatum | . | . | . | . | . | . | . | . | . | . | . | 2 | . | 1 | . | . | . |
| #order | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| # rel | 5 | 24 | 23 | 43 | 22 | 16 | 5 | 5 | 10 | 23 | 8 | 8 | 6 | 5 | 2 | 5 | 2 | 1 | 7 | 18 | 8 | 5 | 8 | 21 | 12 | 11 | 9 |
| # spp | 19 | 68 | 73 | 94 | 59 | 28 | 14 | 9 | 7 | 50 | 38 | 45 | 32 | 26 | 7 | 22 | 8 | 9 | 22 | 19 | 14 | 24 | 59 | 51 | 18 | 34 | 18 |
| Argyrochosmetea niveae | _ | _ | _ | _ | _ | _ | _ | _ | _ | ||||||||||||||||||
| Saxifrago magellanicae-Leucherietum daucifoliae | _ | _ | _ | _ | _ | _ | _ | ||||||||||||||||||||
| Saxifraga magellanica | 100 | 2 | 3 | 1 | 1 | 80 | 13 | ||||||||||||||||||||
| Leucheria daucifolia | 100 | 25 | |||||||||||||||||||||||||
| Weberbauera arequipa | 60 | ||||||||||||||||||||||||||
| Gentianella sp. | 40 | ||||||||||||||||||||||||||
| Perezia pinnatifida | 40 | 5 | 25 | ||||||||||||||||||||||||
| Valeriana coarctata | 40 | 2 | 20 | ||||||||||||||||||||||||
| Calamagrostietea vicunarum | |||||||||||||||||||||||||||
| Calamagrostis vicunarum | 33 | 52 | 53 | 18 | 61 | 50 | 50 | 67 | 20 | 6 | 13 | 50 | 2 | 2 | 4 | ||||||||||||
| Parastrephietalia quadrangularis | |||||||||||||||||||||||||||
| Parastrephia quadrangularis | 13 | 83 | 2 | 65 | 100 | 88 | 100 | 40 | + | 1 | |||||||||||||||||
| Tetraglochin cristatum | 92 | 35 | 14 | 43 | 13 | 63 | 83 | 60 | 2 | ||||||||||||||||||
| Descurainia depressa | 60 | 8 | 13 | 7 | |||||||||||||||||||||||
| Pycnophyllum molle | 40 | 100 | 43 | 93 | 68 | 38 | 57 | 50 | 11 | 25 | 13 | 2 | 2 | + | |||||||||||||
| Hypochaeris meyeniana | 60 | 50 | 70 | 30 | 14 | 13 | 1 | 1 | |||||||||||||||||||
| Ephedra rupestris | 20 | 8 | 9 | 26 | 32 | 30 | 25 | 67 | 1 | 1 | |||||||||||||||||
| Weberbauera spathulifolia | 60 | 4 | 26 | 16 | 18 | 1 | 14 | 40 | |||||||||||||||||||
| Belloa piptolepis | 38 | 65 | 51 | 32 | 6 | 30 | 13 | + | 6 | 20 | + | 1 | |||||||||||||||
| Senecio candollei | 4 | 17 | 37 | 5 | 44 | 13 | 2 | ||||||||||||||||||||
| Senecio humillimus | 13 | 22 | 14 | 18 | 25 | 1 | 1 | ||||||||||||||||||||
| Brayopsis calycina | 46 | 4 | 33 | 23 | |||||||||||||||||||||||
| Chaetanthera stuebelii | 20 | 44 | 23 | ||||||||||||||||||||||||
| Nototriche anthemidifolia | 58 | 49 | 64 | ||||||||||||||||||||||||
| Draba macleanii | 17 | 28 | 18 | 63 | |||||||||||||||||||||||
| Erigeron rosulatus | 25 | 37 | 18 | ||||||||||||||||||||||||
| Hypochaeris eriolaena | 13 | 12 | 17 | ||||||||||||||||||||||||
| Plantago sericea var. lanuginosa | 8 | 13 | 23 | ||||||||||||||||||||||||
| Werneria aretioides | 38 | 42 | 13 | 4 | |||||||||||||||||||||||
| Senecio spinosus | 29 | 26 | 9 | 39 | 13 | 25 | 33 | 40 | + | ||||||||||||||||||
| Werneria pectinata | 46 | 17 | 23 | 6 | 4 | 1 | |||||||||||||||||||||
| Silene genovevae | 33 | 22 | 7 | ||||||||||||||||||||||||
| Perezia coerulescens | 13 | 13 | 9 | 4 | 13 | 43 | 6 | 25 | 38 | 1 | + | ||||||||||||||||
| Wahlenbergia peruviana | 13 | 17 | 14 | ||||||||||||||||||||||||
| Nototriche digitulifolia | 8 | 22 | 5 | ||||||||||||||||||||||||
| Nototriche pedicularifolia | 29 | 16 | 1 | ||||||||||||||||||||||||
| Lupinus ananeanus | 8 | 26 | 5 | ||||||||||||||||||||||||
| Cerastium subspicatum | 21 | 22 | 5 | 2 | |||||||||||||||||||||||
| Conyza deserticola | 8 | 9 | |||||||||||||||||||||||||
| Lobivia maximiliana | 9 | ||||||||||||||||||||||||||
| Oxalis calachaccensis | 8 | ||||||||||||||||||||||||||
| Gomphrena meyeniana | 8 | 20 | |||||||||||||||||||||||||
| Calamagrostion minimae | |||||||||||||||||||||||||||
| Calamagrostis minima | 29 | 13 | 2 | + | |||||||||||||||||||||||
| Werneria nubigena | 17 | 20 | |||||||||||||||||||||||||
| Oreomyrrhis andicola | 42 | 1 | |||||||||||||||||||||||||
| Calandrinia acaulis | 80 | 13 | 19 | + | 4 | ||||||||||||||||||||||
| Astragalo minimi-Azorelletum diapensioidis | |||||||||||||||||||||||||||
| Azorella diapensioides | 67 | 9 | 13 | 50 | 1 | + | |||||||||||||||||||||
| Astragalus minimus | 83 | 26 | 2 | ||||||||||||||||||||||||
| Bougueria nubicola | 67 | 17 | 5 | ||||||||||||||||||||||||
| Werneria apiculata | 58 | 4 | 13 | ||||||||||||||||||||||||
| Werneria melanandra | 38 | ||||||||||||||||||||||||||
| Nototriche longirostris | 38 | 13 | 1 | ||||||||||||||||||||||||
| Junellia minima | 29 | ||||||||||||||||||||||||||
| Aciachne pulvinata | 46 | 4 | 4 | ||||||||||||||||||||||||
| Nototriche turritela | 63 | 25 | 1 | 3 | |||||||||||||||||||||||
| Ourisia muscosa | 13 | ||||||||||||||||||||||||||
| Viola hillii | 4 | ||||||||||||||||||||||||||
| Azorello compactae-Festucion orthophyllae | |||||||||||||||||||||||||||
| Azorella compacta | 78 | 74 | 77 | 10 | 48 | 88 | 38 | 1 | 4 | + | |||||||||||||||||
| Festuca orthophylla | 63 | 22 | 51 | 46 | 100 | 100 | 50 | 1 | + | 2 | 4 | ||||||||||||||||
| Baccharis caespitosa | 13 | 12 | 36 | 9 | 13 | 17 | + | 1 | 43 | 20 | 38 | 3 | + | + | |||||||||||||
| Belloa sp. (# 3945) | 17 | 7 | 27 | ||||||||||||||||||||||||
| Bowlesia tropaeolifolia | 4 | 2 | 18 | + | 43 | 6 | 20 | ||||||||||||||||||||
| Baccharis tricuneata | 35 | 33 | 46 | 22 | 75 | 75 | 33 | 100 | 1 | ||||||||||||||||||
| Cumulopuntia boliviana subsp. ignescens | 22 | 14 | 43 | 25 | 20 | + | |||||||||||||||||||||
| Bartsia elongata | 9 | 14 | 25 | 40 | |||||||||||||||||||||||
| Caiophora rosulata | 9 | 18 | |||||||||||||||||||||||||
| Senecio evacoides | 2 | 32 | 20 | 14 | 25 | ||||||||||||||||||||||
| Stangea wandae | 16 | 27 | |||||||||||||||||||||||||
| Astragalo pusillii-Parastrephietum quadrangularis | |||||||||||||||||||||||||||
| Astragalus pusillus | 65 | 4 | 3 | ||||||||||||||||||||||||
| Sisyrinchium brevipes | 8 | 22 | |||||||||||||||||||||||||
| Sisyrinchium trinervis | 17 | ||||||||||||||||||||||||||
| Laennecia artemisioides | 17 | ||||||||||||||||||||||||||
| Valeriana aschersoniana | 4 | ||||||||||||||||||||||||||
| Belloa longifolia | 4 | 13 | 50 | 20 | 1 | ||||||||||||||||||||||
| Descurainia sp. (# 0940) | 4 | ||||||||||||||||||||||||||
| Junellia pappigera | 4 | ||||||||||||||||||||||||||
| Senecioni moqueguensis-Pycnophylletum molle | |||||||||||||||||||||||||||
| Senecio moqueguensis | 84 | ||||||||||||||||||||||||||
| Senecio tassaensis | 4 | 26 | 13 | ||||||||||||||||||||||||
| Nototriche argentea | 21 | 5 | 1 | ||||||||||||||||||||||||
| Arenaria acaulis | 16 | ||||||||||||||||||||||||||
| Gentianella primuloides | 16 | ||||||||||||||||||||||||||
| Nototriche sepaliloba | 16 | ||||||||||||||||||||||||||
| Cerastium behmianum | 14 | ||||||||||||||||||||||||||
| Xenophyllum weddellii | 14 | ||||||||||||||||||||||||||
| Nototriche pusilla | 12 | ||||||||||||||||||||||||||
| Oritrophium sp. (# 2194b) | 12 | ||||||||||||||||||||||||||
| Senecio sykorae | 12 | ||||||||||||||||||||||||||
| Perezia coerulescens var. amplibracteata | 9 | ||||||||||||||||||||||||||
| Werneria heteroloba | 9 | ||||||||||||||||||||||||||
| Myrosmodes sp. (# 2287) | 7 | ||||||||||||||||||||||||||
| Poa brevis | 7 | ||||||||||||||||||||||||||
| Senecio graveolens | 7 | 4 | 13 | 17 | 13 | ||||||||||||||||||||||
| Senecio sp.1 (# 4217a) | 7 | ||||||||||||||||||||||||||
| Spergularia andina | 4 | 7 | |||||||||||||||||||||||||
| Lupinus sp. (# 2424) | 5 | ||||||||||||||||||||||||||
| Senecio scorzonerifolius | 5 | ||||||||||||||||||||||||||
| Viola sp. (# 4217a) | 5 | ||||||||||||||||||||||||||
| Werneria sp. (# 3940) | 5 | ||||||||||||||||||||||||||
| Xenophyllum digitatum | 5 | ||||||||||||||||||||||||||
| Calamagrostio trichophyllae-Azorelletum compactae | |||||||||||||||||||||||||||
| Nototriche mandoniana | 77 | ||||||||||||||||||||||||||
| Calamagrostis trichophylla | 77 | ||||||||||||||||||||||||||
| Mniodes caespititia | 68 | 6 | |||||||||||||||||||||||||
| Silene mandonii | 68 | ||||||||||||||||||||||||||
| Senecio sp.2 (# 3935) | 64 | ||||||||||||||||||||||||||
| Erigeron lanceolatus | 46 | ||||||||||||||||||||||||||
| Werneria glaberrima | 9 | 41 | |||||||||||||||||||||||||
| Nototriche pedatiloba | 36 | ||||||||||||||||||||||||||
| Perezia pungens | 36 | 25 | 20 | ||||||||||||||||||||||||
| Draba soratensis | 32 | ||||||||||||||||||||||||||
| Bartsia sp. (# 3092) | 2 | 18 | |||||||||||||||||||||||||
| Poa gilgiana | 18 | 2 | 60 | ||||||||||||||||||||||||
| Agrostis breviculmis | 14 | ||||||||||||||||||||||||||
| Viola granulosa | 14 | 1 | |||||||||||||||||||||||||
| Lupinus chilensis | 5 | ||||||||||||||||||||||||||
| Anthochloo-Diesliochloetea | |||||||||||||||||||||||||||
| Anthochloa lepidula | 40 | 13 | 5 | 9 | 94 | 80 | 40 | 13 | 25 | 4 | + | + | |||||||||||||||
| Dielsiochloa floribunda | 5 | 31 | 60 | 60 | 4 | 88 | 1 | 4 | |||||||||||||||||||
| Nototrichion obcuneatae (Nototricho obcuneatae-Xenophylletum poposi) | |||||||||||||||||||||||||||
| Nototriche obcuneata | 6 | 60 | 20 | 9 | 2 | 1 | 4 | ||||||||||||||||||||
| Senecio adenophyllus | 5 | 14 | 56 | 20 | 40 | 11 | 38 | 63 | 4 | 1 | 2 | ||||||||||||||||
| Senecio sp.3 (# 3931) | 14 | 25 | 40 | 40 | |||||||||||||||||||||||
| Dissanthelium calycinum | 38 | 26 | 26 | 18 | 6 | 60 | 100 | 9 | 38 | 1 | |||||||||||||||||
| Nototriche sp.1 (# 3104) | 5 | 13 | 40 | ||||||||||||||||||||||||
| Nototricho obcuneatae-Xenophylletum poposi Galán de Mera, Cáceres & Gonzáles 2003 | |||||||||||||||||||||||||||
| Mniodes coarctata | 16 | 88 | + | 4 | |||||||||||||||||||||||
| Xenophyllum poposum | 81 | 9 | 4 | ||||||||||||||||||||||||
| Nototriche erinacea | 56 | ||||||||||||||||||||||||||
| Senecio trifurcifolius | 31 | ||||||||||||||||||||||||||
| Poa sp. (# 3099) | 25 | ||||||||||||||||||||||||||
| Nototriche sp.2 (# 4228a) | 19 | ||||||||||||||||||||||||||
| Senecio sp.4 (# 4228b) | 13 | ||||||||||||||||||||||||||
| Poo aequiglumae-Xenophylletum dactylophyllum | |||||||||||||||||||||||||||
| Nototriche sp.3 (# 2447) | 100 | ||||||||||||||||||||||||||
| Poa aequigluma | 100 | ||||||||||||||||||||||||||
| Poa spicigera | 100 | 63 | |||||||||||||||||||||||||
| Xenophyllum dactylophyllum | 80 | 1 | |||||||||||||||||||||||||
| Community of Senecio algens | |||||||||||||||||||||||||||
| Senecio algens | 2 | 80 | 2 | 17 | 13 | 13 | 1 | 1 | |||||||||||||||||||
| Senecio sp.5 (# 3942) | 100 | ||||||||||||||||||||||||||
| Class | |||||||||||||||||||||||||||
| Community of Tarasa nototrichoides and Urtica flabellata | |||||||||||||||||||||||||||
| Urtica flabellata | 100 | ||||||||||||||||||||||||||
| Lachemilla pinnata | 38 | 4 | 100 | 1 | + | ||||||||||||||||||||||
| Tarasa nototrichoides | 4 | 4 | 90 | ||||||||||||||||||||||||
| Perezia multiflora | 17 | 50 | 13 | ||||||||||||||||||||||||
| Jaborosa squarrosa | 40 | 13 | |||||||||||||||||||||||||
| Valeriana sp. (Image DSC075, 03/2014) | 10 | ||||||||||||||||||||||||||
| CRASSULETEA CONNATAE | |||||||||||||||||||||||||||
| Muhlenbergia peruviana | 25 | 13 | 2 | 13 | 83 | ||||||||||||||||||||||
| Crassula connata | 17 | ||||||||||||||||||||||||||
| ARGYROCHOSMETEA NIVEAE; Salpichroetalia glandulosae | |||||||||||||||||||||||||||
| Asplenium peruvianum | 9 | 4 | 71 | 11 | 100 | ||||||||||||||||||||||
| Belloa kunthiana | 9 | ||||||||||||||||||||||||||
| Belloa schultzii | 4 | 2 | 29 | 6 | 38 | ||||||||||||||||||||||
| Loricaria graveolens | 13 | ||||||||||||||||||||||||||
| Salpichroa glandulosa | 2 | 5 | + | 43 | 22 | 20 | |||||||||||||||||||||
| Valeriana nivalis | 9 | 5 | 2 | 1 | 14 | 94 | 25 | 13 | 2 | + | |||||||||||||||||
| Companions | |||||||||||||||||||||||||||
| Calamagrostis curvula | 17 | 44 | 36 | 56 | 100 | 80 | 39 | 13 | 100 | 13 | 4 | + | |||||||||||||||
| Trisetum spicatum | 40 | 33 | 4 | 2 | 36 | 19 | + | 20 | 38 | ||||||||||||||||||
| Luzula vulcanica | 33 | 13 | 44 | 27 | 13 | ||||||||||||||||||||||
| Parastrephia lucida | 4 | 51 | 86 | 19 | 60 | 30 | 1 | 4 | 4 | ||||||||||||||||||
| Pycnophyllum glomeratum | 8 | 4 | 44 | 23 | 44 | 20 | 13 | 1 | 4 | 4 | |||||||||||||||||
| Microsteris gracilis | 100 | 21 | 26 | 23 | |||||||||||||||||||||||
| Paronychia andina | 33 | 39 | 9 | 14 | 3 | 3 | |||||||||||||||||||||
| Senecio nutans | 30 | 46 | 25 | 60 | 17 | 100 | 67 | 1 | + | ||||||||||||||||||
| Stangea rhizanta | 60 | 12 | 32 | 60 | 20 | 13 | |||||||||||||||||||||
| Astragalus peruvianus | 17 | 17 | 2 | ||||||||||||||||||||||||
| Bartsia diffusa | 8 | 26 | 5 | 13 | 1 | 6 | 13 | ||||||||||||||||||||
| Lepidium meyenii | 13 | 9 | 5 | 1 | |||||||||||||||||||||||
| Cardionema ramosissimum | 8 | 4 | 2 | ||||||||||||||||||||||||
| Mniodes sp. (# 2477) | 26 | 9 | 13 | ||||||||||||||||||||||||
| Poa candamoana | 40 | 9 | 50 | 13 | 50 | 83 | 100 | ||||||||||||||||||||
| Stipa ichu | 60 | 29 | 78 | 26 | 38 | 75 | 100 | 20 | 1 | ||||||||||||||||||
| Astragalus uniflorus | 7 | 6 | 13 | ||||||||||||||||||||||||
| Chaetanthera peruviana | 4 | 14 | 13 | ||||||||||||||||||||||||
| Geranium sessiliflorum | 9 | 23 | 13 | 20 | + | 20 | 1 | 1 | |||||||||||||||||||
| Mancoa hispida | 13 | 4 | |||||||||||||||||||||||||
| Adesmia spinosissima | 9 | 22 | 13 | 20 | |||||||||||||||||||||||
| Cyperus seslerioides | 13 | 13 | |||||||||||||||||||||||||
| Descurainia athrocarpa | 2 | 6 | 13 | 1 | |||||||||||||||||||||||
| Galium corymbosum | 35 | 13 | 2 | 29 | 6 | 60 | |||||||||||||||||||||
| Lupinus cuzcensis | 4 | ||||||||||||||||||||||||||
| Oxalis debilis | 4 | ||||||||||||||||||||||||||
| Oxalis nubigena | 9 | 29 | 1 | ||||||||||||||||||||||||
| Weberbauera peruviana | 4 | ||||||||||||||||||||||||||
| Species from other communities | |||||||||||||||||||||||||||
| Lupinus paruroensis | 17 | 13 | 67 | ||||||||||||||||||||||||
| Chersodoma jodopappa | 4 | 13 | 33 | 20 | |||||||||||||||||||||||
| Deyeuxia heterophylla | 25 | 13 | 67 | 20 | |||||||||||||||||||||||
| Deyeuxia breviaristata | 13 | + | 3 | ||||||||||||||||||||||||
| Astragalus arequipensis | 13 | ||||||||||||||||||||||||||
| Festuca rigescens | 13 | 13 | + | ||||||||||||||||||||||||
| Lupinus saxatilis | 38 | 40 | |||||||||||||||||||||||||
| Erigeron incarum | 13 | 17 | |||||||||||||||||||||||||
| Luzula racemosa | 4 | 38 | 1 | ||||||||||||||||||||||||
| Festuca dolichophylla | 100 | 100 | 100 | 14 | 25 | ||||||||||||||||||||||
| Deyeuxia cabrerae | 17 | 6 | 13 | 13 | + | 2 | |||||||||||||||||||||
| Paranephelius ovatus | 75 | 17 | 60 | 1 | |||||||||||||||||||||||
| Coreopsis fasciculata | 75 | 60 | |||||||||||||||||||||||||
| Plantago sericea | 13 | 67 | |||||||||||||||||||||||||
| Plantago linearis | 25 | 20 | |||||||||||||||||||||||||
| Nassella pubiflora | 25 | 17 | |||||||||||||||||||||||||
| Dissanthelinum macusaniense | 17 | 25 | |||||||||||||||||||||||||
| Nassella mucronata | 38 | 83 | 60 | ||||||||||||||||||||||||
| Trifolium amabile | 63 | 17 | |||||||||||||||||||||||||
| Bromus catharticus | 38 | 20 | |||||||||||||||||||||||||
| Lupinus microphyllus | 17 | 20 | |||||||||||||||||||||||||
| Senecio modestus | 1 | 1 | 40 | 13 | |||||||||||||||||||||||
| Draba brackenridgei | + | 29 | |||||||||||||||||||||||||
| Mniodes andina | + | 88 | |||||||||||||||||||||||||
| Oxalis andina | 1 | + | 60 | 1 | |||||||||||||||||||||||
| Senecio rhizomatus | 1 | + | 20 | ||||||||||||||||||||||||
| Cystopteris fragilis | + | 20 | |||||||||||||||||||||||||
| Asplenium triphyllum | 1 | 11 | 40 | ||||||||||||||||||||||||
| Senecio culcitioides | 1 | 14 | 28 | 13 | 50 | ||||||||||||||||||||||
| Woodsia montevidensis | 1 | 14 | |||||||||||||||||||||||||
| Caiophora horrida | 1 | 1 | |||||||||||||||||||||||||
| Sisymbium peruvianum | 1 | 6 | |||||||||||||||||||||||||
| Stipa rigida | 1 | 20 | 63 | ||||||||||||||||||||||||
| Xenophyllum ciliolatum | 25 | 2 | |||||||||||||||||||||||||
| Silene andicola | 13 | 1 | + | 2 | |||||||||||||||||||||||
| Poa humilluma | 38 | 1 | |||||||||||||||||||||||||
| Adesmia patancana | 1 | ||||||||||||||||||||||||||
| Perezia ciliosa | 1 | ||||||||||||||||||||||||||
| Other species: In 10 Baccharis incarum 57; in 10 Nassella brachyphylla 13, Echinopsis pamparuizii 9, Baccharis genistelloides 9, Hypochaeris taraxacoides 9, Stipa nardoides 4, Stipa rigidiseta 4, Plantago monticola 4, Nassella asplundii 4, Polylepis tarapacana 13, Arenaria serpens 9, Dissanthelium peruvianum 9, Werneria sp. 4, Oxalis pachyrrhiza 4, Calamagrostis intermedia 4, Caiophora chuquitensis 4, Bomarea dulcis 4, Dissanthelinum breve 4; in 11 Stipa obtusa 25, Opuntia lagopus 13, Baccharis buxifolia 13, Adesmia miraflorensis 13, Polylepis rugulosa 25, Senecio adenophylloides 13, Senecio rudbeckiifolius 13, Stipa annua 13; in 12 Nassella pubiflora 25, Tunilla soehrensii 13, Quinchamalium procumbens 13, Echinopsis pampana 13, Vulpia megalura 38, Tagetes multiflora 25, Bidens pilosa 25, Gilia laciniata 13, Lepechinia meyenii 13, Nassella depauperata 13, Schkhuria multiflora 13, Chondrosum simplex 13, Hordeum muticum 13, Hypochaeris chillensis 13, Erodium cicutarium 13, Lepidium chichicara 13; in 13 Bartsia camporum 50, Mutisia arequipensis 17, Gnaphalium dombeyanum 17; in 14 Agrostis gelida 100; in 16 Asplenium castaneum +, Peperomia peruviana 2, Oxalis arenaria 1; in 17 Draba cryptantha 2; in 18 Gnaphalium sp. 1, Cerastium mucronatum 1, Phacelia pinnata 1, Lobivia caespitosa 1; in 19 Bomarea uniflora 57, Bowlesia sodiroana 14, Oxalis petrophila 14, Valeriana plectritoides var. pallida 14, Galium aparine 14, Poa horridula 14; in 20 Draba cuzcoensis 6, Ribes brachybothrys 6; in 22 Peperomia peruviana 60, Oxalis arenaria 40, Asplenium castaneum 20, Englerocharis peruviana 20; in 23 Xenophyllum decorum 88, Plettkea cryptantha 38, Xenophyllum digitatum 13, Chaetanthera cochlearifolia 63, Weberbauera rosulans 13, Niphogeton dissecta 25, Gentianella weberbaueri 13, Englerocharis peruviana 50, Nototriche aretioides 38, Nototriche pinnata 13, Plantago lamprophylla 13, Bromus villosissimus 38, Pycnophyllum weberbaueri 13, Pernettya prostrata 13, Niphogeton scabra 13, Astragalus brackenridgei 13, Deyeuxia tarmensis 13, Senecio tephrosioides 13, Deyeuxia ovata 25, Deyeuxia rigescens 13, Deyeuxia glacialis 13, Senecio canescens 13, Ephedra americana 38, Melpomene moliniformis 13, Valeriana candamoana 38; in 24 Poa gymnantha 18, Astragalus micranthellus 1, Bartsia crenata 1, Bromus lanatus 1, Chaetanthera boliviensis 1, Chaetanthera villosa 1, Deyeuxia cephalanta 1, Deyeuxia densiflora 1, Deyeuxia glaciaris 1, Gnaphalium badium 1, Niphogetom scabra 1, Poa aequigluma 1, Pycnophyllum filiforme 1, Senecio puchii 1, Valeriana interrupta 1, Viola nivalis 1; in 25 Deyeuxia deserticola 9; in 26 Nototriche sp. | |||||||||||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montesinos-Tubée, D.B.; Cleef, A.M.; Sýkora, K.V. The Subnival Vegetation of Moquegua, South Peru: Chasmophytes, Grasslands and Cushion Communities. Ecologies 2021, 2, 71-111. https://doi.org/10.3390/ecologies2010005
Montesinos-Tubée DB, Cleef AM, Sýkora KV. The Subnival Vegetation of Moquegua, South Peru: Chasmophytes, Grasslands and Cushion Communities. Ecologies. 2021; 2(1):71-111. https://doi.org/10.3390/ecologies2010005
Chicago/Turabian StyleMontesinos-Tubée, Daniel B., Antoine M. Cleef, and Karlè V. Sýkora. 2021. "The Subnival Vegetation of Moquegua, South Peru: Chasmophytes, Grasslands and Cushion Communities" Ecologies 2, no. 1: 71-111. https://doi.org/10.3390/ecologies2010005
APA StyleMontesinos-Tubée, D. B., Cleef, A. M., & Sýkora, K. V. (2021). The Subnival Vegetation of Moquegua, South Peru: Chasmophytes, Grasslands and Cushion Communities. Ecologies, 2(1), 71-111. https://doi.org/10.3390/ecologies2010005