
Hydrogen-Rich Water Prevents Dehydration-Induced Cellular Oxidative Stress and Cell Death in Human Skin Keratinocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydrogen-Rich Water Preparation
2.2. Cell Culture
2.3. Drying Experiment
2.4. Giemsa Stain
2.5. Cell Viability Assay
2.6. Cellular ROS Detection
2.7. Scanning Electron Microscope Analysis
2.8. Statistical Analysis
3. Results
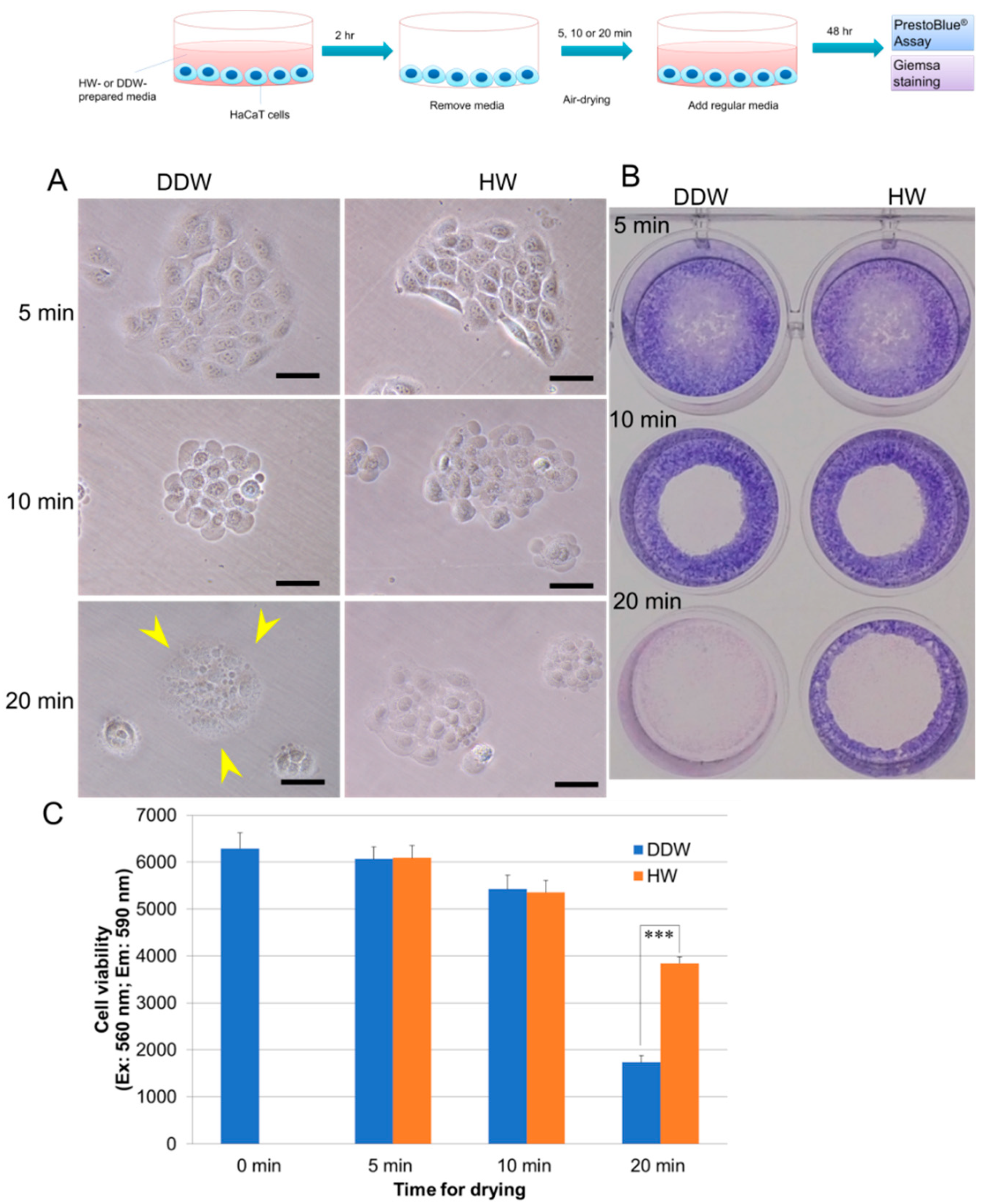
3.1. HW Prevents Dehydration-Induced Cell Death in Human Skin Keratinocytes HaCaT
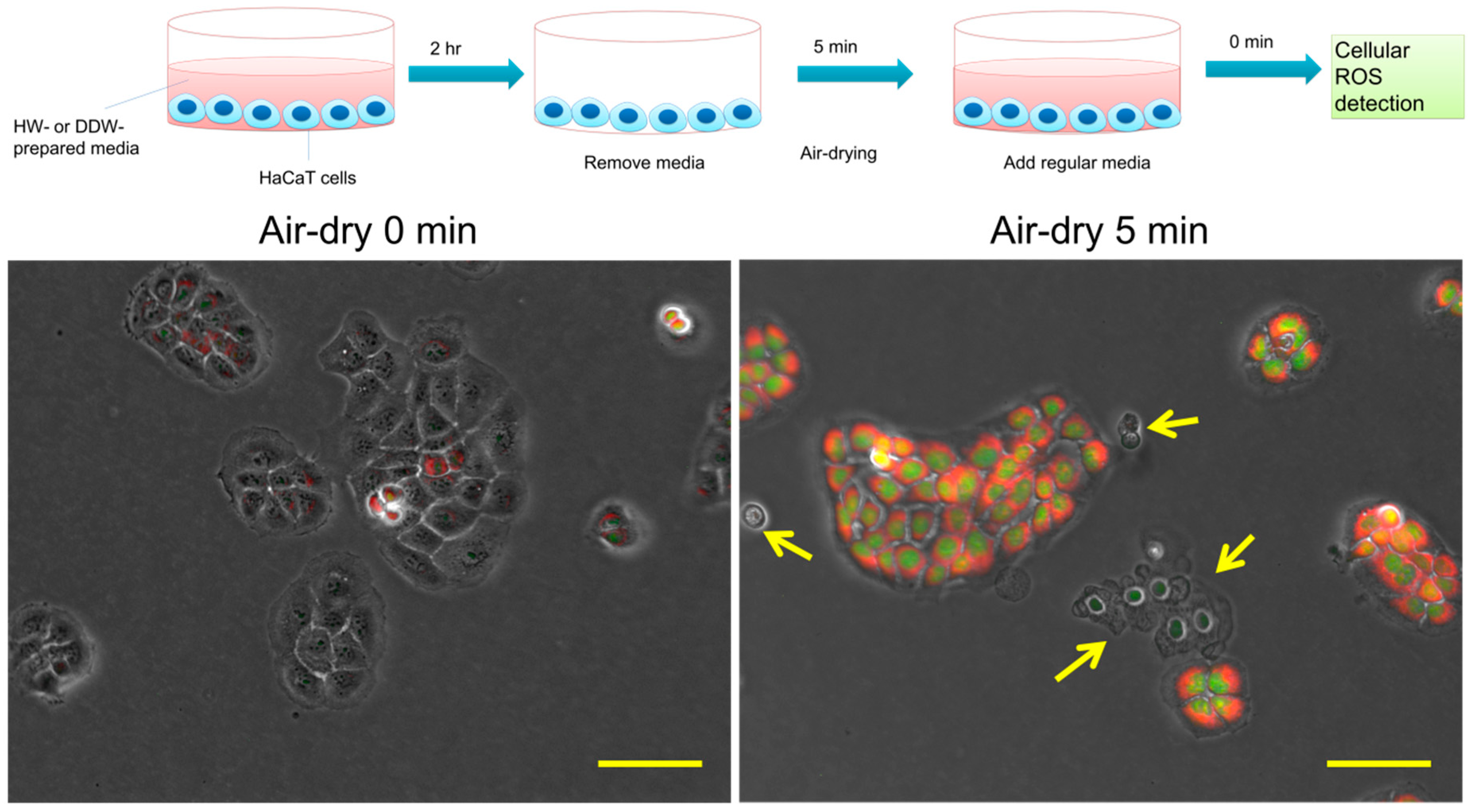
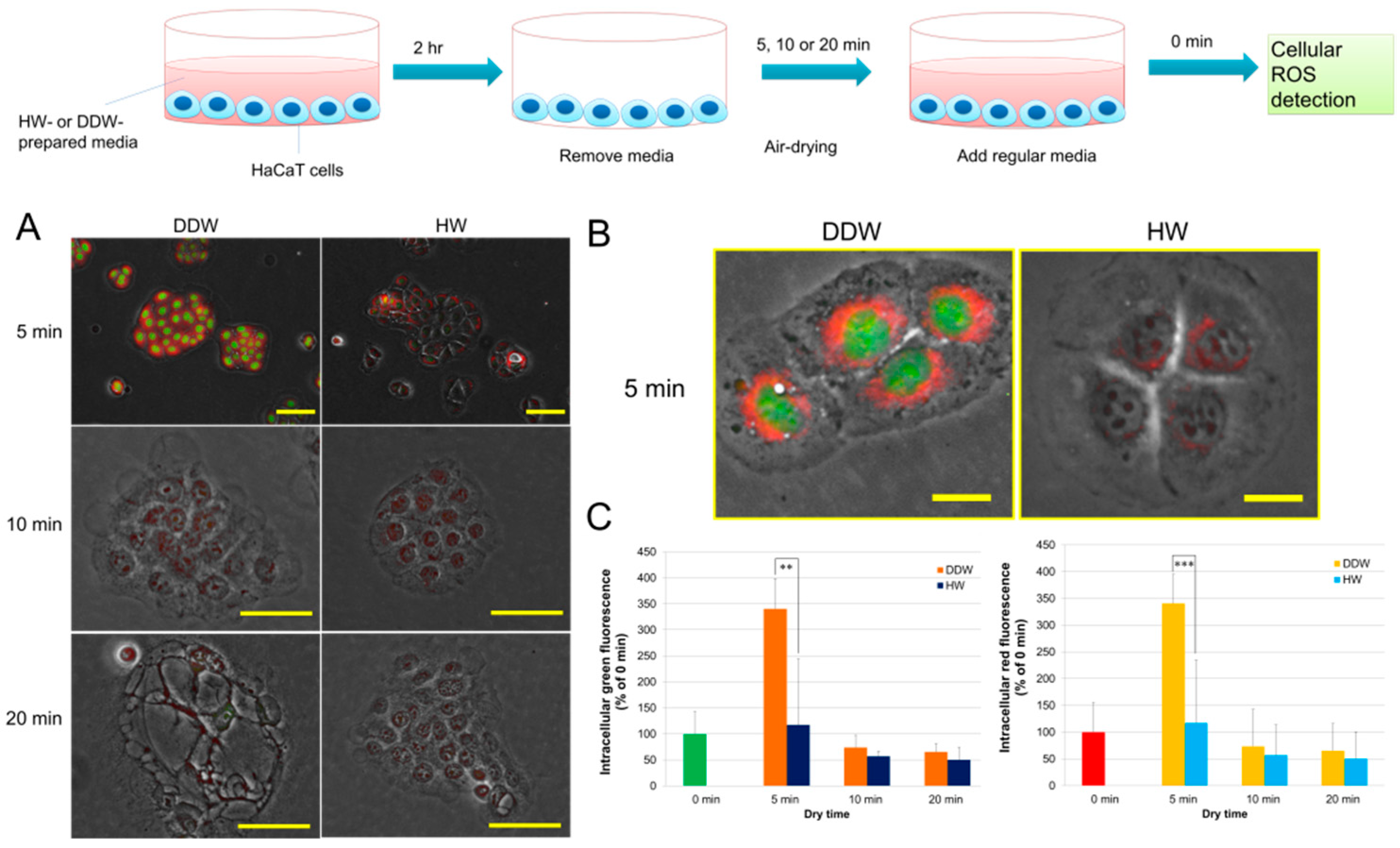
3.2. HW Diminishes 5 min Air-Drying-Induced Intracellular ROS Generation in HaCaT Cells

3.3. HW Reduces 10 min Air-Drying-Induced Apoptosis-like Cell Death and Membrane Damage in HaCaT Cells


4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | reactive oxygen species |
| HW | hydrogen-rich water |
| DDW | double distilled water |
| SEM | scanning electron microscope |
| DH | dissolved hydrogen |
| ORP | oxidation-reduction potential |
| DMEM | Dulbecco’s modified Eagle’s medium |
| AVP | arginine vasopressin |
References
- França, M.B.; Panek, A.D.; Eleutherio, E.C. Oxidative stress and its effects during dehydration. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Koster, M.I. Making an epidermis. Ann. N. Y. Acad. Sci. 2009, 1170, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Russell, G.; Nenov, A.; Kisher, H.; Hancock, J.T. Molecular Hydrogen as Medicine: An Assessment of Administration Methods. Hydrogen 2021, 2, 444–460. [Google Scholar] [CrossRef]
- Xiao, L.; Miwa, N. Hydrogen-rich water achieves cytoprotection from oxidative stress injury in human gingival fibroblasts in culture or 3D-tissue equivalents, and wound-healing promotion, together with ROS-scavenging and relief from glutathione diminishment. Hum. Cell 2017, 30, 72–87. [Google Scholar] [CrossRef]
- Xiao, L.; Mochizuki, M.; Nakahara, T.; Miwa, N. Hydrogen-Generating Silica Material Prevents UVA-ray-Induced Cellular Oxidative Stress, Cell Death, Collagen Loss and Melanogenesis in Human Cells and 3D Skin Equivalents. Antioxidants 2021, 10, 76. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Kajiyama, S.; Hasegawa, G.; Asano, M.; Hosoda, H.; Fukui, M.; Nakamura, N.; Kitawaki, J.; Imai, S.; Nakano, K.; Ohta, M.; et al. Supplementation of hydrogen-rich water improves lipid and glucose metabolism in patients with type 2 diabetes or impaired glucose tolerance. Nutr. Res. 2008, 28, 137–143. [Google Scholar] [CrossRef]
- Sim, M.; Kim, C.S.; Shon, W.J.; Lee, Y.K.; Choi, E.Y.; Shin, D.M. Hydrogen-rich water reduces inflammatory responses and prevents apoptosis of peripheral blood cells in healthy adults: A randomized, double-blind, controlled trial. Sci. Rep. 2020, 10, 12130. [Google Scholar] [CrossRef]
- Mikami, T.; Tano, K.; Lee, H.; Lee, H.; Park, J.; Ohta, F.; LeBaron, T.W.; Ohta, S. Drinking hydrogen water enhances endurance and relieves psychometric fatigue: A randomized, double-blind, placebo-controlled study 1. Can. J. Physiol. Pharmacol. 2019, 97, 857–862. [Google Scholar] [CrossRef]
- Gu, Y.; Huang, C.S.; Inoue, T.; Yamashita, T.; Ishida, T.; Kang, K.M.; Nakao, A. Drinking hydrogen water ameliorated cognitive impairment in senescence-accelerated mice. J. Clin. Biochem. Nutr. 2010, 46, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Seike, T.; Yutsudo, N.; Ohno, M.; Yamada, H.; Yamaguchi, H.; Sakumi, K.; Yamakawa, Y.; Kido, M.A.; Takaki, A.; et al. Hydrogen in drinking water reduces dopaminergic neuronal loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. PLoS ONE 2009, 4, e7247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Miwa, N. Hydrogen Nano-Bubble Water Suppresses ROS Generation, Adipogenesis, and Interleukin-6 Secretion in Hydrogen-Peroxide- or PMA-Stimulated Adipocytes and Three-Dimensional Subcutaneous Adipose Equivalents. Cells 2021, 10, 626. [Google Scholar] [CrossRef] [PubMed]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Liao, F.; Fan, Y.; Miwa, N. Enzyme-digested Colla Corii Asini (E’jiao) accelerates wound healing and prevents ultraviolet A-induced collagen synthesis decline and wrinkle formation in three-dimensional skin equivalents. Hum. Cell 2020, 33, 1056–1067, Correction in Hum. Cell 2021, 34, 291. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Liao, F.; Ide, R.; Horie, T.; Fan, Y.; Saiki, C.; Miwa, N. Enzyme-digested Colla Corii Asini (E’jiao) prevents hydrogen peroxide-induced cell death and accelerates amyloid beta clearance in neuronal-like PC12 cells. Neural Regen. Res. 2020, 15, 2270–2272. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Aoshima, H.; Saitoh, Y.; Miwa, N. The effect of squalane-dissolved fullerene-C60 on adipogenesis-accompanied oxidative stress and macrophage activation in a preadipocyte-monocyte co-culture system. Biomaterials 2010, 31, 5976–5985. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Sakagami, H.; Miwa, N. A New Method for Testing Filtration Efficiency of Mask Materials Under Sneeze-like Pressure. In Vivo 2020, 34, 1637–1644. [Google Scholar] [CrossRef]
- Lavizzo-Mourey, R.J. Dehydration in the elderly: A short review. J. Natl. Med. Assoc. 1987, 79, 1033–1038. [Google Scholar]
- Bahouth, M.N.; Gottesman, R.F.; Szanton, S.L. Primary ‘dehydration’ and acute stroke: A systematic research review. J. Neurol. 2018, 265, 2167–2181. [Google Scholar] [CrossRef]
- Watso, J.C.; Farquhar, W.B. Hydration Status and Cardiovascular Function. Nutrients 2019, 11, 1866. [Google Scholar] [CrossRef] [Green Version]
- Manz, F. Hydration and Disease. J. Am. Coll. Nutr. 2007, 26, 541S. [Google Scholar] [CrossRef]
- McDermott, B.P.; Anderson, S.A.; Armstrong, L.E.; Casa, D.J.; Cheuvront, S.N.; Cooper, L.; Kenney, W.L.; O’Connor, F.G.; Roberts, W.O. National Athletic Trainers’ Association Position Statement: Fluid Replacement for the Physically Active. J. Athl. Train. 2017, 52, 877–895. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Abu Damir, H.; Ali, O.M.; Amir, N.; Tariq, S.; Greenwood, M.P.; Lin, P.; Gillard, B.; Murphy, D.; Adem, A. The effect of long-term dehydration and subsequent rehydration on markers of inflammation, oxidative stress and apoptosis in the camel kidney. BMC Vet. Res. 2020, 16, 458. [Google Scholar] [CrossRef]
- Balasubramanian, S.K.; Wolkers, W.F.; Bischof, J.C. Membrane hydration correlates to cellular biophysics during freezing in mammalian cells. Biochim. Biophys. Acta 2009, 1788, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Pereira Ede, J.; Panek, A.D.; Eleutherio, E.C. Protection against oxidation during dehydration of yeast. Cell Stress Chaperones 2003, 8, 120–124. [Google Scholar] [CrossRef] [Green Version]
- McKersie, B.D.; Stinson, R.H. Effect of Dehydration on Leakage and Membrane Structure in Lotus corniculatus L. Seeds. Plant Physiol. 1980, 66, 316–320. [Google Scholar] [CrossRef] [Green Version]
- Cruz de Carvalho, R.; Catalá, M.; Branquinho, C.; Marques da Silva, J.; Barreno, E. Dehydration rate determines the degree of membrane damage and desiccation tolerance in bryophytes. Physiol. Plant. 2017, 159, 277–289. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Storey, K.B. Role of antioxidant defenses in the tolerance of severe dehydration by anurans. The case of the leopard frog Rana pipiens. Mol. Cell. Biochem. 1998, 189, 79–89. [Google Scholar] [CrossRef]
- Espindola, A.; Gomes, D.S.; Panek, A.D.; Eleutherio, E.C. The role of glutathione in yeast dehydration tolerance. Cryobiology 2003, 47, 236–241. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DH (24 °C) | DH (100 °C ⟹ 24 °C) 1 | ORP (24 °C) | ORP (100 °C ⟹ 24 °C) 1 | |
|---|---|---|---|---|
| HW | 880 ± 38 ppb | 420 ± 25 ppb | −341 ± 15 mv | −160 ± 10 mv |
| DDW | 46 ± 12 ppb | N/A 2 | 571 ± 26 mV | N/A 2 |
| HW-DMEM | DDW-DMEM | For 100 mL | Final Ratio (%) |
|---|---|---|---|
| HW | DDW | 70 mL | 70 |
| DMEM 10× | DMEM 10× | 10 mL | 10 |
| NaHCO3 0.23 M | NaHCO3 0.23 M | 10 mL | 10 |
| FBS | FBS | 10 mL | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Miwa, N. Hydrogen-Rich Water Prevents Dehydration-Induced Cellular Oxidative Stress and Cell Death in Human Skin Keratinocytes. Hydrogen 2022, 3, 62-71. https://doi.org/10.3390/hydrogen3010005
Xiao L, Miwa N. Hydrogen-Rich Water Prevents Dehydration-Induced Cellular Oxidative Stress and Cell Death in Human Skin Keratinocytes. Hydrogen. 2022; 3(1):62-71. https://doi.org/10.3390/hydrogen3010005
Chicago/Turabian StyleXiao, Li, and Nobuhiko Miwa. 2022. "Hydrogen-Rich Water Prevents Dehydration-Induced Cellular Oxidative Stress and Cell Death in Human Skin Keratinocytes" Hydrogen 3, no. 1: 62-71. https://doi.org/10.3390/hydrogen3010005
APA StyleXiao, L., & Miwa, N. (2022). Hydrogen-Rich Water Prevents Dehydration-Induced Cellular Oxidative Stress and Cell Death in Human Skin Keratinocytes. Hydrogen, 3(1), 62-71. https://doi.org/10.3390/hydrogen3010005