
Lactic Acid Fermentation of Carrageenan Hydrolysates from the Macroalga Kappaphycus alvarezii: Evaluating Different Bioreactor Operation Modes


Abstract
:1. Introduction
2. Material and Methods
2.1. Microorganism, Propagation and Fermentation Media
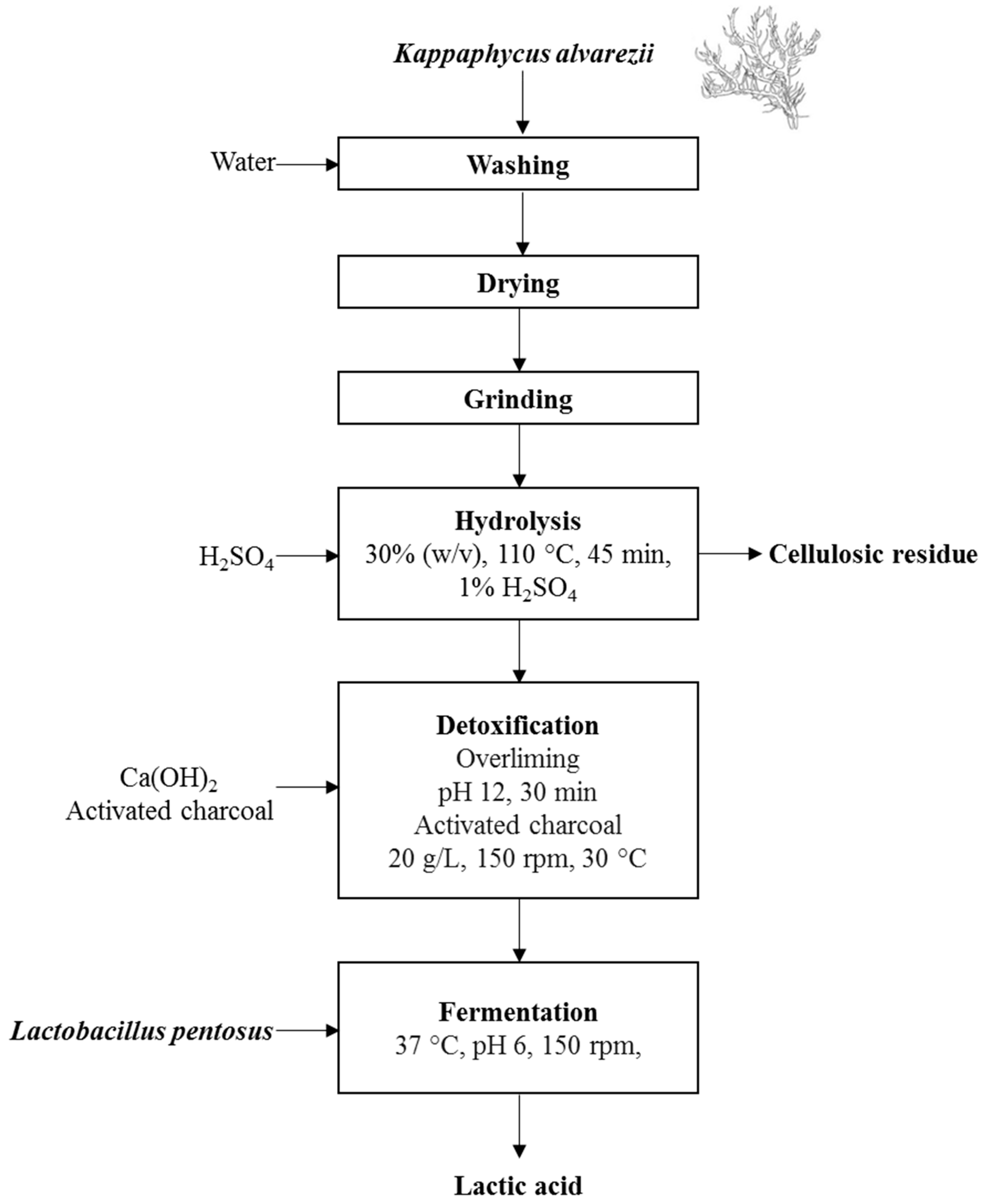
2.2. Hydrolysate and Acclimation Procedures
2.3. Bioreactor Fermentation Essays
2.4. Regeneration of Activated Charcoal
2.5. Analytical Methods
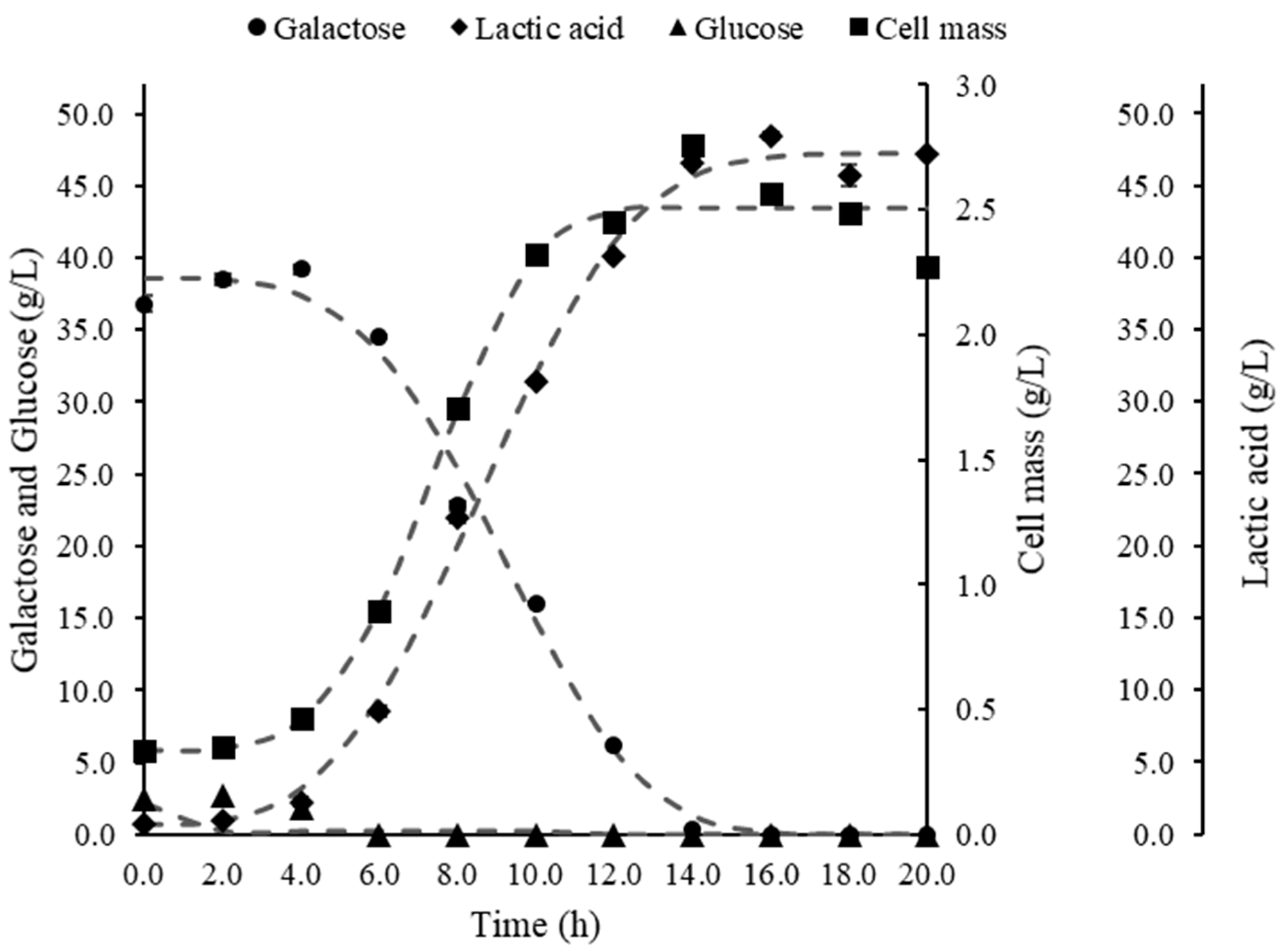
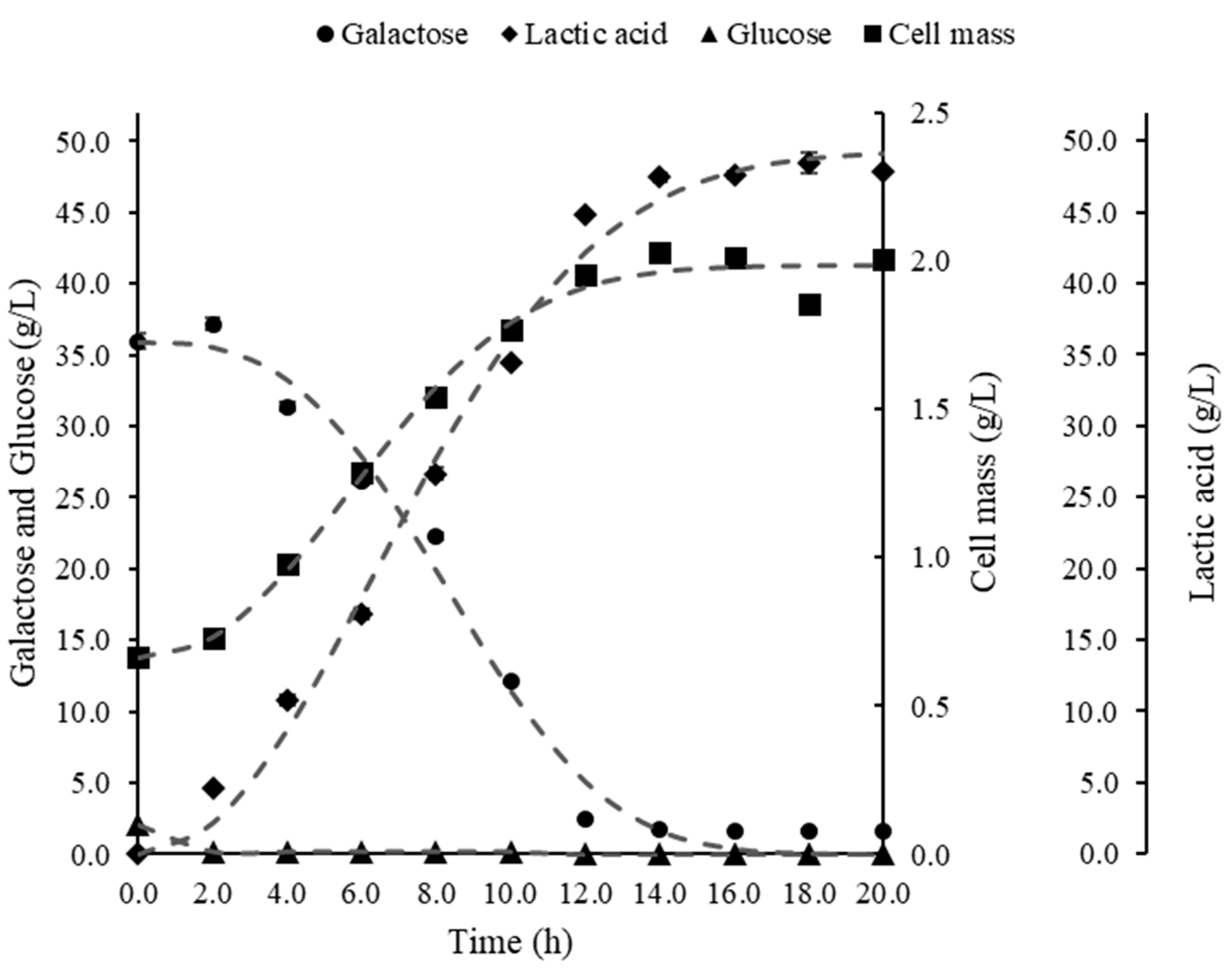
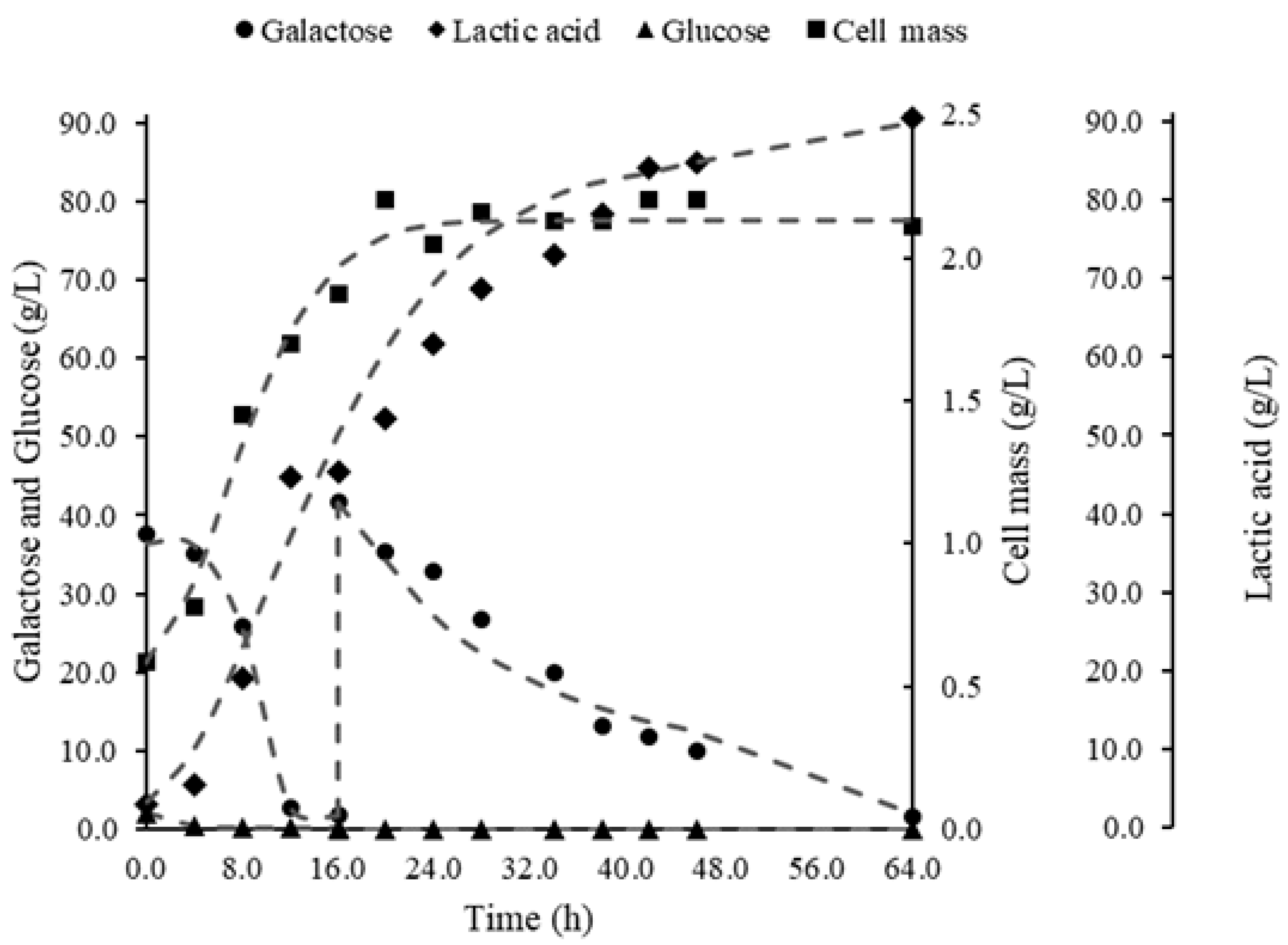
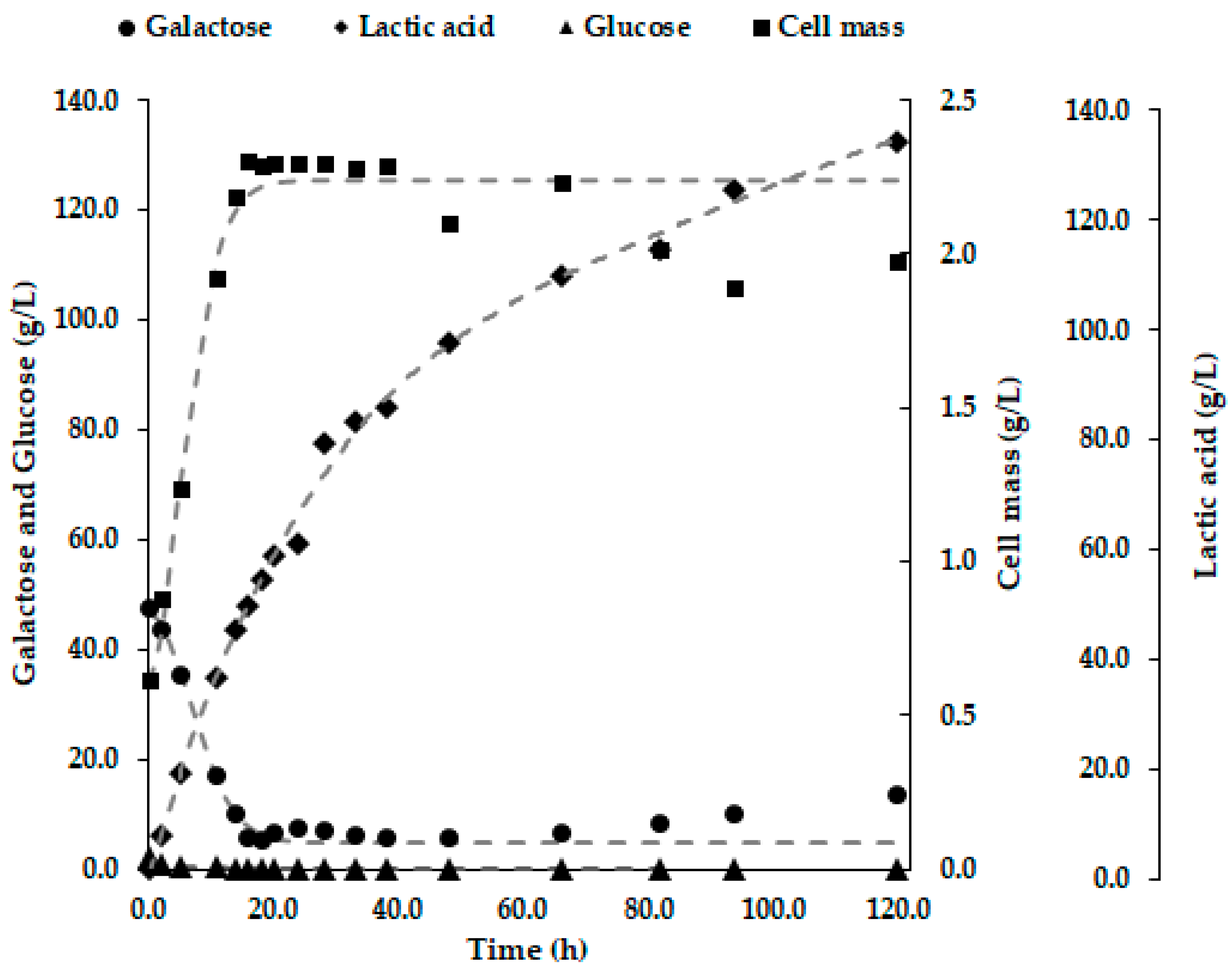
3. Results
3.1. Bioreactor Fermentations
3.2. Regenerability of Activated Charcoal
4. Discussion
4.1. Comparison of Bioreactor Operation Systems
4.2. Regeneration of Activated Charcoal
4.3. K. alvarezii Biorefinery for Lactic Acid Production Proposal
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.; Nielsen, J. Biobased Organic Acids Production by Metabolically Engineered Microorganisms. Curr. Opin. Biotechnol. 2016, 37, 165–172. [Google Scholar] [CrossRef]
- Goldberg, I.; Rokem, J.S.; Pines, O. Organic Acids: Old Metabolites, New Themes. J. Chem. Technol. Biotechnol. 2006, 81, 1601–1611. [Google Scholar] [CrossRef]
- Panda, S.K.; Sahu, L.; Behera, S.K.; Ray, R.C. Research and Production of Organic Acids and Industrial Potential. In Bioprocessing for Biomolecules Production; John Wiley & Sons, Ltd.: Chichester, UK, 2019; pp. 195–209. [Google Scholar]
- Njokweni, S.G.; Steyn, A.; Botes, M.; Viljoen-Bloom, M.; van Zyl, W.H. Potential Valorization of Organic Waste Streams to Valuable Organic Acids through Microbial Conversion: A South African Case Study. Catalysts 2021, 11, 964. [Google Scholar] [CrossRef]
- Zapaśnik, A.; Sokołowska, B.; Bryła, M. Role of Lactic Acid Bacteria in Food Preservation and Safety. Foods 2022, 11, 1283. [Google Scholar] [CrossRef]
- Komesu, A.; Oliveira, J.A.R.d.; Martins, L.H.d.S.; Wolf Maciel, M.R.; Maciel Filho, R. Lactic Acid Production to Purification: A Review. Bioresources 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Ghaffar, T.; Irshad, M.; Anwar, Z.; Aqil, T.; Zulifqar, Z.; Tariq, A.; Kamran, M.; Ehsan, N.; Mehmood, S. Recent Trends in Lactic Acid Biotechnology: A Brief Review on Production to Purification. J. Radiat. Res. Appl. Sci. 2014, 7, 222–229. [Google Scholar] [CrossRef]
- Song, L.; Yang, D.; Liu, R.; Liu, S.; Dai, L.; Dai, X. Microbial Production of Lactic Acid from Food Waste: Latest Advances, Limits, and Perspectives. Bioresour. Technol. 2022, 345, 126052. [Google Scholar] [CrossRef] [PubMed]
- Augustiniene, E.; Valanciene, E.; Matulis, P.; Syrpas, M.; Jonuskiene, I.; Malys, N. Bioproduction of L- and d-Lactic Acids: Advances Trends in Microbial Strain Application And. Crit. Rev. Biotechnol. 2022, 42, 342–360. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent Advances in Lactic Acid Production by Microbial Fermentation Processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production—Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef]
- Pot, B.; Felis, G.E.; Bruyne, K.D.; Tsakalidou, E.; Papadimitriou, K.; Leisner, J.; Vandamme, P. The Genus Lactobacillus. In Lactic Acid Bacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 249–353. [Google Scholar]
- Wischral, D.; Arias, J.M.; Modesto, L.F.; de França Passos, D.; Pereira, N. Lactic Acid Production from Sugarcane Bagasse Hydrolysates by Lactobacillus Pentosus: Integrating Xylose and Glucose Fermentation. Biotechnol. Prog. 2019, 35, e2718. [Google Scholar] [CrossRef] [PubMed]
- Tabacof, A.; Calado, V.; Pereira, N. Third Generation Lactic Acid Production by Lactobacillus pentosus from the Macroalgae Kappaphycus alvarezii Hydrolysates. Fermentation 2023, 9, 319. [Google Scholar] [CrossRef]
- Buyondo, J.P.; Lui, S. Lactic Acid Production by Lactobacillus pentosus from Wood Extract Hydrolysate. J. Sci. Technol. For. Prod. Process 2011, 1, 38–47. [Google Scholar]
- Wang, J.; Huang, J.; Laffend, H.; Jiang, S.; Zhang, J.; Ning, Y.; Fang, M.; Liu, S. Optimization of Immobilized Lactobacillus Pentosus Cell Fermentation for Lactic Acid Production. Bioresour. Bioprocess 2020, 7, 15. [Google Scholar] [CrossRef]
- Ahmad, A.; Banat, F.; Taher, H. A Review on the Lactic Acid Fermentation from Low-Cost Renewable Materials: Recent Developments and Challenges. Env. Technol. Innov. 2020, 20, 101138. [Google Scholar] [CrossRef]
- Djukić-Vuković, A.; Mladenović, D.; Ivanović, J.; Pejin, J.; Mojović, L. Towards Sustainability of Lactic Acid and Poly-Lactic Acid Polymers Production. Renew. Sustain. Energy Rev. 2019, 108, 238–252. [Google Scholar] [CrossRef]
- Nagarajan, D.; Chen, C.-Y.; Ariyadasa, T.U.; Lee, D.-J.; Chang, J.-S. Macroalgal Biomass as a Potential Resource for Lactic Acid Fermentation. Chemosphere 2022, 309, 136694. [Google Scholar] [CrossRef]
- Sudhakar, K.; Mamat, R.; Samykano, M.; Azmi, W.H.; Ishak, W.F.W.; Yusaf, T. An Overview of Marine Macroalgae as Bioresource. Renew. Sustain. Energy Rev. 2018, 91, 165–179. [Google Scholar] [CrossRef]
- Campbell, R.; Hotchkiss, S. Carrageenan Industry Market Overview. In Tropical Seaweed Farming Trends, Problems and Opportunities; Springer International Publishing: Cham, Switzerland, 2017; pp. 193–205. [Google Scholar]
- Torres, M.D.; Kraan, S.; Domínguez, H. Seaweed Biorefinery. Rev. Environ. Sci. Biotechnol. 2019, 18, 335–388. [Google Scholar] [CrossRef]
- Lange, L.; Bak, U.G.; Hansen, S.C.B.; Gregersen, O.; Harmsen, P.; Karlsson, E.N.; Meyer, A.; Mikkelsen, M.D.; Van Den Broek, L.; Hreggviðsson, G.Ó. Opportunities for Seaweed Biorefinery. In Sustainable Seaweed Technologies: Cultivation, Biorefinery, and Applications; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–31. ISBN 9780128179437. [Google Scholar]
- Álvarez-Viñas, M.; Flórez-Fernández, N.; Torres, M.D.; Domínguez, H. Successful Approaches for a Red Seaweed Biorefinery. Mar. Drugs 2019, 17, 620. [Google Scholar] [CrossRef]
- Balina, K.; Romagnoli, F.; Blumberga, D. Seaweed Biorefinery Concept for Sustainable Use of Marine Resources. Energy Procedia 2017, 128, 504–511. [Google Scholar] [CrossRef]
- Sadhukhan, J.; Gadkari, S.; Martinez-Hernandez, E.; Ng, K.S.; Shemfe, M.; Torres-Garcia, E.; Lynch, J. Novel Macroalgae (Seaweed) Biorefinery Systems for Integrated Chemical, Protein, Salt, Nutrient and Mineral Extractions and Environmental Protection by Green Synthesis and Life Cycle Sustainability Assessments. Green Chem. 2019, 21, 2635–2655. [Google Scholar] [CrossRef]
- Chung, M.R.W.Y.; Tan, I.S.; Foo, H.C.Y.; Lam, M.K.; Lim, S. Potential of Macroalgae-Based Biorefinery for Lactic Acid Production from Exergy Aspect. Biomass Convers. Biorefinery 2023, 13, 2623–2653. [Google Scholar] [CrossRef]
- Mears, L.; Stocks, S.M.; Sin, G.; Gernaey, K.V. A Review of Control Strategies for Manipulating the Feed Rate in Fed-Batch Fermentation Processes. J. Biotechnol. 2017, 245, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, S. A New Approach for Finding Smooth Optimal Feeding Profiles in Fed-Batch Fermentations. Biochem. Eng. J. 2016, 105, 177–188. [Google Scholar] [CrossRef]
- Bai, D.-M.; Wei, Q.; Yan, Z.-H.; Zhao, X.-M.; Li, X.-G.; Xu, S.-M. Fed-Batch Fermentation of Lactobacillus lactis for Hyper-Production of l-Lactic Acid. Biotechnol. Lett. 2003, 25, 1833–1835. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, P.M.; Santos, L.P.; Coelho, L.F.; Avila Neto, P.M.; Sass, D.C.; Contiero, J. Production of L (+) Lactic Acid by Lactobacillus Casei Ke11: Fed Batch Fermentation Strategies. Fermentation 2021, 7, 151. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Xiao, Y.; Tashiro, Y.; Wang, Y.; Zendo, T.; Sakai, K.; Sonomoto, K. Fed-Batch Fermentation for Enhanced Lactic Acid Production from Glucose/Xylose Mixture without Carbon Catabolite Repression. J. Biosci. Bioeng. 2015, 119, 153–158. [Google Scholar] [CrossRef]
- Ding, S.; Tan, T. L-Lactic Acid Production by Lactobacillus Casei Fermentation Using Different Fed-Batch Feeding Strategies. Process Biochem. 2006, 41, 1451–1454. [Google Scholar] [CrossRef]
- Machado, J.; Rossi, D.M.; Ayub, M.A.Z. Batch and Fed-Batch Strategies of Lactic Acid Production by Lactobacillus plantarum BL011 Using Soybean Hull Hydrolysates as Substrate. Biomass Convers. Biorefinery 2022, 1–11. [Google Scholar] [CrossRef]
- Lobeda, K.; Jin, Q.; Wu, J.; Zhang, W.; Huang, H. Lactic Acid Production from Food Waste Hydrolysate by Lactobacillus pentosus: Focus on Nitrogen Supplementation, Initial Sugar Concentration, PH, and Fed-batch Fermentation. J. Food Sci. 2022, 87, 3071–3083. [Google Scholar] [CrossRef]
- Jang, S.; Shirai, Y.; Uchida, M.; Wakisaka, M. Production of L(+)-Lactic Acid from Mixed Acid and Alkali Hydrolysate of Brown Seaweed. Food Sci. Technol. Res. 2011, 17, 155–160. [Google Scholar] [CrossRef]
- Mwiti, G.; Yeo, I.-S.; Jeong, K.-H.; Choi, H.-S.; Kim, J. Activation of Galactose Utilization by the Addition of Glucose for the Fermentation of Agar Hydrolysate Using Lactobacillus brevis ATCC 14869. Biotechnol. Lett. 2022, 44, 823–830. [Google Scholar] [CrossRef]
- Carratalá-Abril, J.; Lillo-Ródenas, M.A.; Linares-Solano, A.; Cazorla-Amorós, D. Regeneration of Activated Carbons Saturated with Benzene or Toluene Using an Oxygen-Containing Atmosphere. Chem. Eng. Sci. 2010, 65, 2190–2198. [Google Scholar] [CrossRef]
- Cubas-Cano, E.; González-Fernández, C.; Tomás-Pejó, E. Evolutionary Engineering of Lactobacillus Pentosus Improves Lactic Acid Productivity from Xylose-Rich Media at Low PH. Bioresour. Technol. 2019, 288, 121540. [Google Scholar] [CrossRef]
- Song, L.; Liu, S.; Liu, R.; Yang, D.; Dai, X. Direct Lactic Acid Production from Household Food Waste by Lactic Acid Bacteria. Sci. Total Environ. 2022, 840, 156479. [Google Scholar] [CrossRef]
- Taleghani, H.G.; Najafpour, G.D.; Ghoreyshi, A.A. A Study on the Effect of Parameters on Lactic Acid Production from Whey. Pol. J. Chem. Technol. 2016, 18, 58–63. [Google Scholar] [CrossRef]
- Wardani, S.K.; Cahyanto, M.N.; Rahayu, E.S.; Utami, T. The Effect of Inoculum Size and Incubation Temperature on Cell Growth, Acid Production and Curd Formation during Milk Fermentation by Lactobacillus plantarum Dad 13. Int. Food Res. J. 2017, 24, 921–926. [Google Scholar]
- Kuo, Y.-C.; Yuan, S.-F.; Wang, C.-A.; Huang, Y.-J.; Guo, G.-L.; Hwang, W.-S. Production of Optically Pure L-Lactic Acid from Lignocellulosic Hydrolysate by Using a Newly Isolated and D-Lactate Dehydrogenase Gene-Deficient Lactobacillus Paracasei Strain. Bioresour. Technol. 2015, 198, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Sato, Y.; Hama, S.; Tanaka, T.; Noda, H.; Kondo, A.; Honda, K. L-Lactate Oxidase-mediated Removal of L-lactic Acid Derived from Fermentation Medium for the Production of Optically Pure D-lactic Acid. Biotechnol. J. 2022, 17, 2100331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vadlani, P.V.; Kumar, A.; Hardwidge, P.R.; Govind, R.; Tanaka, T.; Kondo, A. Enhanced D-Lactic Acid Production from Renewable Resources Using Engineered Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 2016, 100, 279–288. [Google Scholar] [CrossRef]
- Yoshida, S.; Okano, K.; Tanaka, T.; Ogino, C.; Kondo, A. Homo-d-Lactic Acid Production from Mixed Sugars Using Xylose-Assimilating Operon-Integrated Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 2011, 92, 67–76. [Google Scholar] [CrossRef]
- Sun, N.; Sun, C.; Liu, J.; Liu, H.; Snape, C.E.; Li, K.; Wei, W.; Sun, Y. Surface-Modified Spherical Activated Carbon Materials for Pre-Combustion Carbon Dioxide Capture. RSC Adv. 2015, 5, 33681–33690. [Google Scholar] [CrossRef]
- Imbierowicz, M.; Troszkiewicz, M.; Piotrowska, K. Heat Effects of Wet Oxidation of Glucose: A Biomass Model Compound. Chem. Eng. J. 2015, 260, 864–874. [Google Scholar] [CrossRef]
- Pereira, É. de M.; Colmati, F.; Amorim, A.M.; Sodré, F.F.; Linares, J.J. Electrochemical Regeneration of Glyphosate-Saturated Carbon as an Alternative for Activated Carbon Reutilization. J. Solid State Electrochem. 2023, 1–8. [Google Scholar] [CrossRef]
- Ahuja, V.; Kshirsagar, S.; Ghosh, P.; Sarkar, B.; Sutar, A.; More, S.; Dasgupta, D. Process Development for Detoxification of Corncob Hydrolysate Using Activated Charcoal for Xylitol Production. J. Environ. Chem. Eng. 2022, 10, 107097. [Google Scholar] [CrossRef]
- Yücel, H.G.; Aksu, Z. Ethanol Fermentation Characteristics of Pichia Stipitis Yeast from Sugar Beet Pulp Hydrolysate: Use of New Detoxification Methods. Fuel 2015, 158, 793–799. [Google Scholar] [CrossRef]
- Fazal-ur-Rehman, M. Methodological Trends in Preparation of Activated Carbon from Local Sources and Their Impacts on Production-A Review. Curr. Trends Chem. Eng. Process Technol. 2018, 4, 109–119. [Google Scholar]
- Santos, D.H.d.S.; Xiao, Y.; Chaukura, N.; Hill, J.M.; Selvasembian, R.; Zanta, C.L.P.S.; Meili, L. Regeneration of Dye-Saturated Activated Carbon through Advanced Oxidative Processes: A Review. Heliyon 2022, 8, e10205. [Google Scholar] [CrossRef]
- Zanella, O.; Tessaro, I.C.; Féris, L.A. Desorption- and Decomposition-Based Techniques for the Regeneration of Activated Carbon. Chem. Eng. Technol. 2014, 37, 1447–1459. [Google Scholar] [CrossRef]
- Yan, X.; Jin, F.; Tohji, K.; Kishita, A.; Enomoto, H. Hydrothermal Conversion of Carbohydrate Biomass to Lactic Acid. AIChE J. 2010, 56, 2727–2733. [Google Scholar] [CrossRef]
- Hargreaves, P.I.; Barcelos, C.A.; da Costa, A.C.A.; Pereira, N. Production of Ethanol 3G from Kappaphycus alvarezii: Evaluation of Different Process Strategies. Bioresour. Technol. 2013, 134, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Abd-Rahim, F.; Wasoh, H.; Rafein, M.; Ariff, A. Food Hydrocolloids Production of High Yield Sugars from Kappaphycus alvarezii Using Combined Methods of Chemical and Enzymatic Hydrolysis. Food Hydrocoll. 2014, 42, 309–315. [Google Scholar] [CrossRef]
- Paz-Cedeno, F.R.; Solórzano-Chávez, E.G.; de Oliveira, L.E.; Gelli, V.C.; Monti, R.; de Oliveira, S.C.; Masarin, F. Sequential Enzymatic and Mild-Acid Hydrolysis of By-Product of Carrageenan Process from Kappaphycus alvarezii. Bioenergy Res. 2019, 12, 419–432. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activated Charcoal | Galactose (g/L) * | Glucose (g/L) * | HMF (g/L) * |
|---|---|---|---|
| Newly purchased activated charcoal | 41.00 ± 0.44 | 3.60 ± 0.10 | 0.70 ± 0.01 |
| After 1st regeneration | 38.83 ± 0.12 | 1.00 ± 0.15 | 0.50 ± 0.01 |
| After 2nd regeneration | 37.40 ± 0.70 | Not detected | 0.21 ± 0.01 |
| After 3rd regeneration | 33.93 ± 0.06 | Not detected | Not detected |
| Fermentation Parameter | Batch a | Batch b [14] | Batch c | Pulse Fed-Batch c | Extended Fed-Batch c |
|---|---|---|---|---|---|
| Final lactic acid solution (g/L) | 47.17 ± 0.12 | 29.39 ± 0.32 | 49.96 ± 0.15 | 90.96 ± 0.32 | 132.59 ± 0.40 |
| Percentage reduction of substrate % | 100 | 94.4 | 95.7 | 95.7 | 89.4 |
| Productivity(g/(L.h)) | 3.37 | 1.05 | 3.57 | 1.37 | 1.10 |
| Yield (g/g) | 1.28 | 1.07 | 1.37 | 1.17 | 1.04 |
| Inoculum size (g/L) | 0.4 | 0.3 | 0.6 | 0.6 | 0.6 |
| Maximum cell concentration (g/L) | 2.76 ± 0.14 | 1.65 ± 0.04 | 2.00 ± 0.01 | 2.20 ± 0.04 | 2.29 ± 0.03 |
| Overall fermentation time (h) | 14 | 28 | 14 | 64 | 120 |
| Final media volume (L) | 1 | 1 | 1 | 1.1 | 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabacof, A.; Calado, V.; Pereira, N., Jr. Lactic Acid Fermentation of Carrageenan Hydrolysates from the Macroalga Kappaphycus alvarezii: Evaluating Different Bioreactor Operation Modes. Polysaccharides 2023, 4, 256-270. https://doi.org/10.3390/polysaccharides4030017
Tabacof A, Calado V, Pereira N Jr. Lactic Acid Fermentation of Carrageenan Hydrolysates from the Macroalga Kappaphycus alvarezii: Evaluating Different Bioreactor Operation Modes. Polysaccharides. 2023; 4(3):256-270. https://doi.org/10.3390/polysaccharides4030017
Chicago/Turabian StyleTabacof, Adam, Verônica Calado, and Nei Pereira, Jr. 2023. "Lactic Acid Fermentation of Carrageenan Hydrolysates from the Macroalga Kappaphycus alvarezii: Evaluating Different Bioreactor Operation Modes" Polysaccharides 4, no. 3: 256-270. https://doi.org/10.3390/polysaccharides4030017
APA StyleTabacof, A., Calado, V., & Pereira, N., Jr. (2023). Lactic Acid Fermentation of Carrageenan Hydrolysates from the Macroalga Kappaphycus alvarezii: Evaluating Different Bioreactor Operation Modes. Polysaccharides, 4(3), 256-270. https://doi.org/10.3390/polysaccharides4030017