
Eco(toxicological) Assessment of the Neonicotinoid Formulation Actara® Using Planarian Girardia tigrina as Model Organism

 , ,
, ,  , ,
, , 

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organisms
2.2. Reagents
2.3. Acute Toxicity Test
2.4. Evaluation of Sublethal Endpoints and Regeneration
2.4.1. Post-Exposure Regeneration
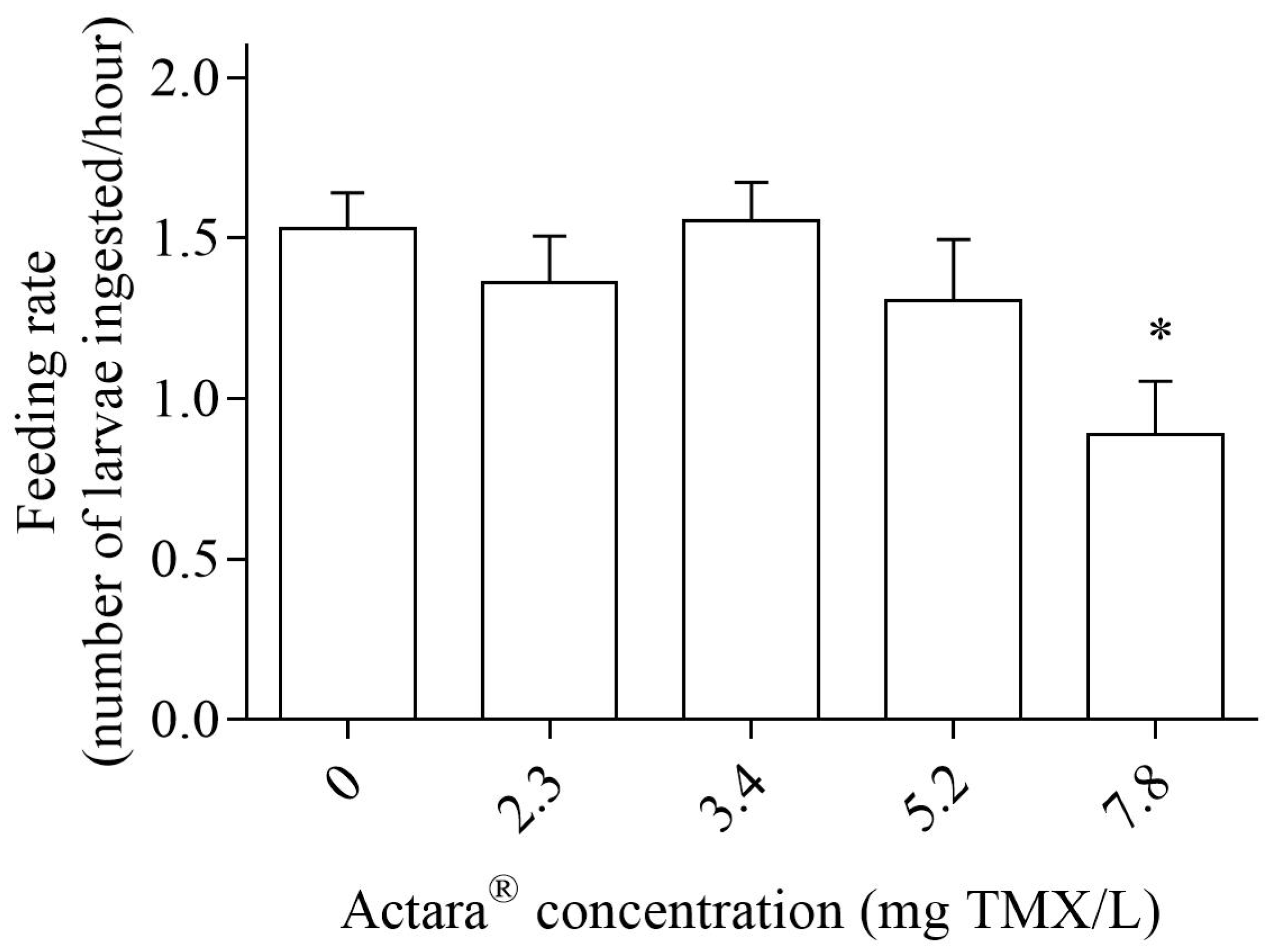
2.4.2. Post-Exposure Feeding Rate
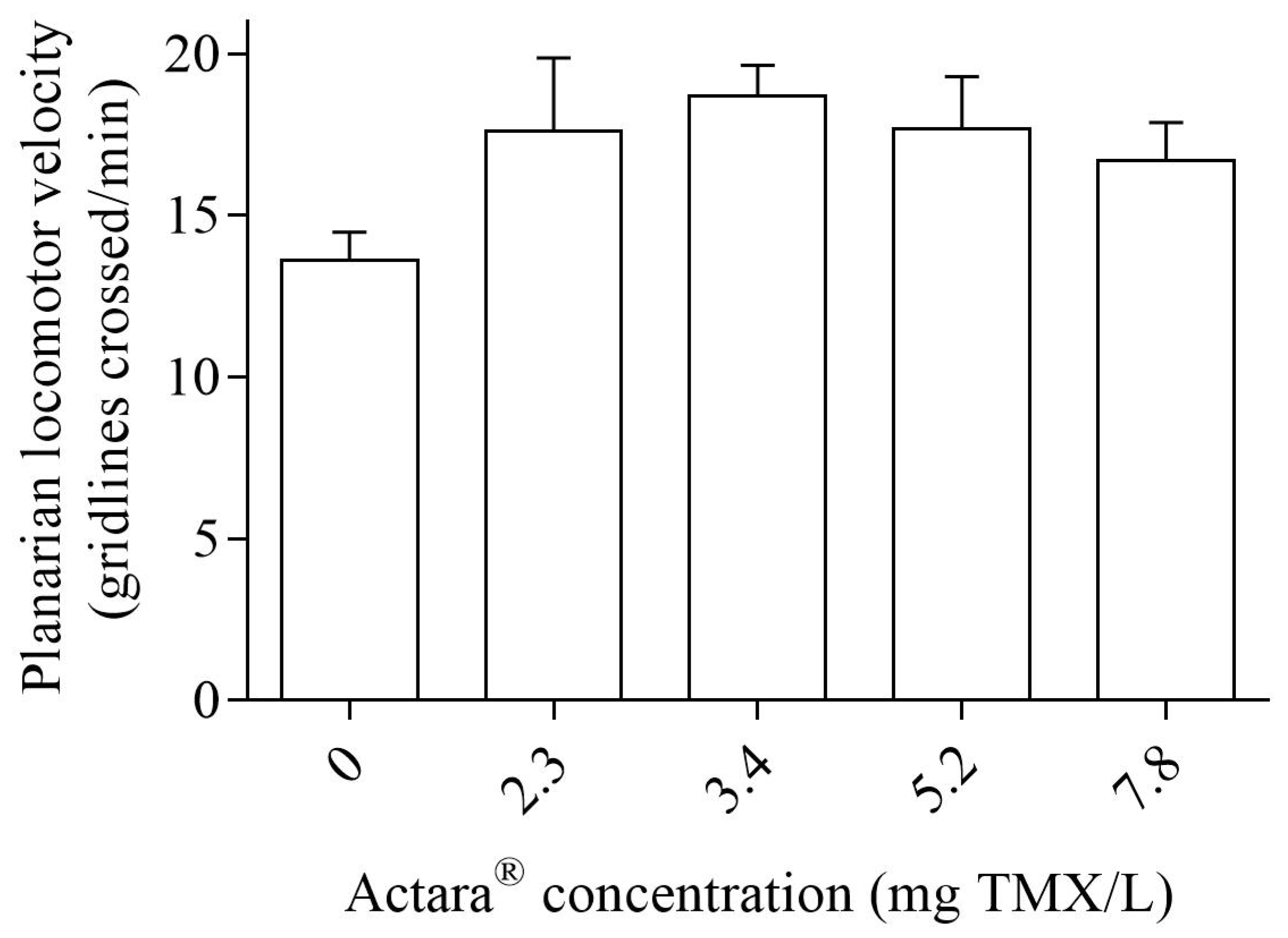
2.4.3. Post-Exposure Locomotor Velocity
2.5. Statistical Analyses
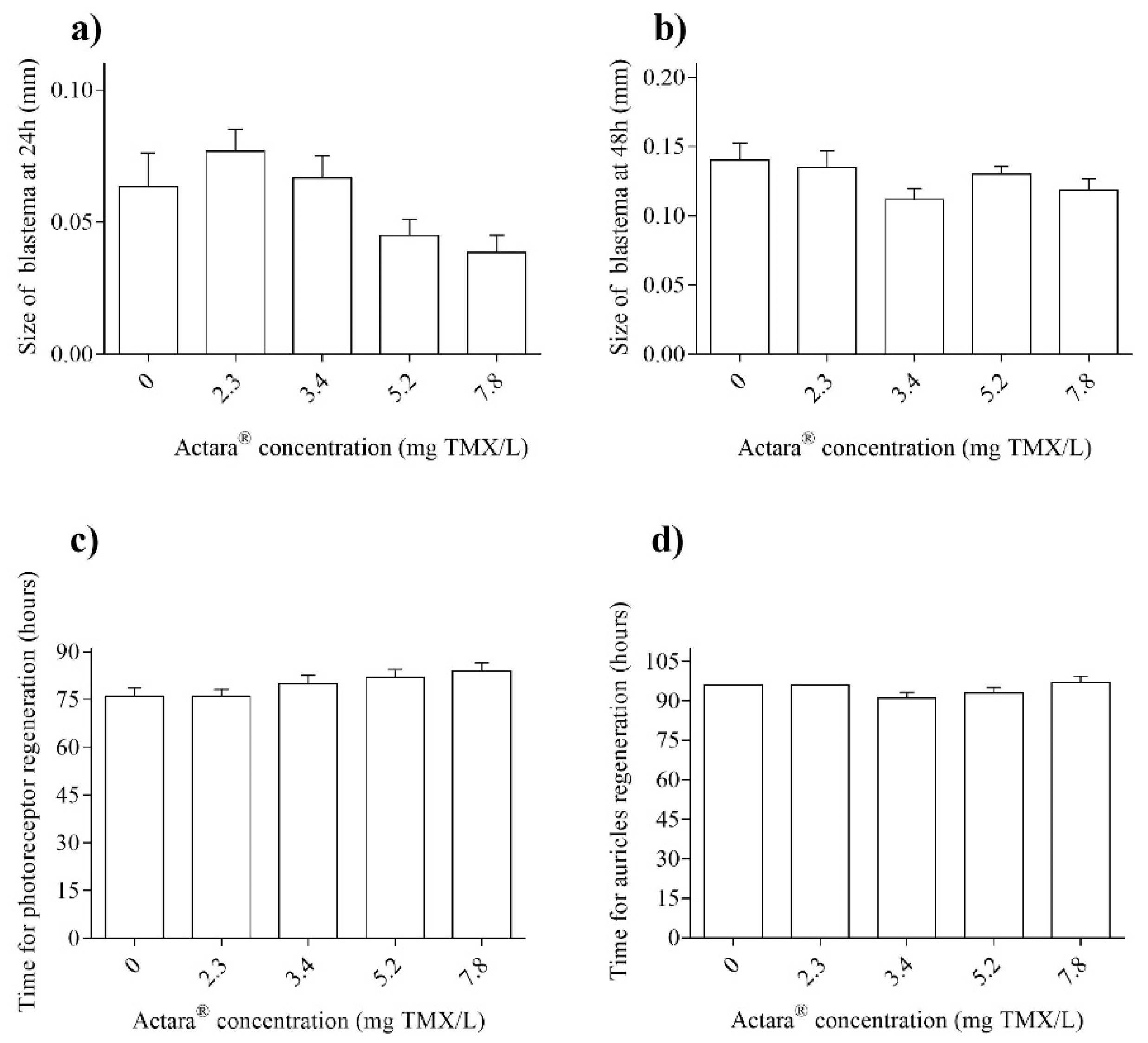
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borsuah, J.F.; Messer, T.L.; Snow, D.D.; Comfort, S.D.; Mittelstet, A.R. Literature Review: Global Neonicotinoid Insecticide Occurrence in Aquatic Environments. Water 2020, 12, 3388. [Google Scholar] [CrossRef]
- Robinson, S.A.; Gavel, M.J.; Richardson, S.D.; Chlebak, R.J.; Milotic, D.; Koprivnikar, J.; Forbes, M.R. Sub-chronic exposure to a neonicotinoid does not affect susceptibility of larval leopard frogs to infection by trematode parasites, via either depressed cercarial performance or host immunity. Parasitol. Res. 2019, 118, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, N.; Chen, K.H.-C.; Huang, J.-C.; Lai, H.-T.; Uapipatanakul, B.; Roldan, M.J.M.; Macabeo, A.P.G.; Ger, T.-R.; Hsiao, C.-D. Physiological Effects of Neonicotinoid Insecticides on Non-Target Aquatic Animals—An Updated Review. Int. J. Mol. Sci. 2021, 22, 9591. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B. Neonicotinoids: Insecticides acting on insect nicotinic acetylcholine receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Buckingham, S.D.; Freeman, J.C.; Squire, M.D.; Baylis, H.A.; Sattelle, D.B. Effects of the α subunit on imidacloprid sensitivity of recombinant nicotinic acetylcholine receptors. Br. J. Pharmacol. 1998, 123, 518–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, K.; Ihara, M.; Sattelle, D.B. Neonicotinoid Insecticides: Molecular Targets, Resistance, and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 241–255. [Google Scholar] [CrossRef]
- Cruz, B.; de Carvalho, R.F.; Silva, T.S.; Sarmento, R.D.A.; Cavallini, G.S.; Pereira, D.H. Adsorptive capacity of a g-C3N4 matrix for thiamethoxam removal: A DFT study. Comput. Theor. Chem. 2022, 1215, 113816. [Google Scholar] [CrossRef]
- de Lima e Silva, C.; de Rooij, W.; Verweij, R.A.; van Gestel, C.A. Toxicity in Neonicotinoids to Folsomia candida and Eisenia andrei. Environ. Toxicol. Chem. 2020, 39, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Simon-Delso, N.; Amaralrogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Hladik, M.L.; Main, A.R.; Goulson, D. Environmental Risks and Challenges Associated with Neonicotinoid Insecticides. Environ. Sci. Technol. 2018, 52, 3329–3335. [Google Scholar] [CrossRef] [Green Version]
- Butler, D. Scientists hail European ban on bee-harming pesticides. Nature 2018, 555, 150–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craddock, H.A.; Huang, D.; Turner, P.C.; Quirós-Alcalá, L.; Payne-Sturges, D.C. Trends in neonicotinoid pesticide residues in food and water in the United States, 1999–2015. Environ. Health 2019, 18, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodcock, B.A.; Ridding, L.; Pereira, M.G.; Sleep, D.; Newbold, L.; Oliver, A.; Shore, R.F.; Bullock, J.M.; Heard, M.S.; Gweon, H.S.; et al. Neonicotinoid use on cereals and sugar beet is linked to continued low exposure risk in honeybees. Agric. Ecosyst. Environ. 2021, 308, 107205. [Google Scholar] [CrossRef]
- Abdourahime, H.; Anastassiadou, M.; Arena, M.; Auteri, D.; Barmaz, S.; Brancato, A.; Brocca, D.; Bura, L.; Cabrera, L.C.; Chiusolo, A.; et al. Peer review of the pesticide risk assessment of the active substance thiacloprid. EFSA J. 2019, 17, e05595. [Google Scholar] [CrossRef] [Green Version]
- Melo, B.S.; Ribeiro, F.; Saraiva, A.S.; Barbosa, R.D.S.; Ferreira, J.S.D.J.; Melo, M.S.; Rosa, L.P.; Gravato, C.; Soares, A.M.; Sarmento, R.A. The multigenerational effects of clothianidin on Chironomus xanthus: Larvae exposed to this acetylcholine super agonist show no clear resistance. Ecotoxicol. Environ. Saf. 2022, 245, 114092. [Google Scholar] [CrossRef]
- Melo, B.S.; Ribeiro, F.; de Souza Saraiva, A.; da Silva Barbosa, R.; de Jesus Ferreira, J.S.; Melo, M.S.; Rosa, L.P.; Dornelas, A.S.; Gravato, C.; Soares, A.M.; et al. Is there a common mechanism of neonicotinoid resistance among insects? Preliminary results show that F1 larvae of pre-exposed Chironomus xanthus are more tolerant to imidacloprid. J. Hazard. Mater. Adv. 2022, 6, 100073. [Google Scholar] [CrossRef]
- Paunescu, A.; Soare, L.C.; Fierascu, I.; Fierascu, R.C.; Mihaescu, C.F.; Tofan, L.; Ponepal, C.M. Ecotoxicological Studies on the Action of Actara 25 WG Insecticide on Prussian Carp (Carassius gibelio) and Marsh Frog (Pelophylax ridibundus). Toxics 2022, 10, 114. [Google Scholar] [CrossRef]
- Anderson, T.A.; Salice, C.J.; Erickson, R.A.; McMurry, S.T.; Cox, S.B.; Smith, L.M. Effects of landuse and precipitation on pesticides and water quality in playa lakes of the southern high plains. Chemosphere 2013, 92, 84–90. [Google Scholar] [CrossRef]
- Ferreira-Junior, D.F.; Sarmento, R.A.; Saraiva, A.S.; Dornelas, A.S.P.; Pestana, J.L.T.; Soares, A.M.V.M. Effects of a Thiamethoxam-Based Insecticide on the Life History of Chironomus xanthus. Water Air Soil Pollut. 2018, 229, 376. [Google Scholar] [CrossRef]
- Saraiva, A.S.; Sarmento, R.A.; Rodrigues, A.C.; Campos, D.; Fedorova, G.; Žlábek, V.; Gravato, C.; Pestana, J.L.; Soares, A.M. Assessment of thiamethoxam toxicity to Chironomus riparius. Ecotoxicol. Environ. Saf. 2017, 137, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, R.d.S.; Cavallini, G.S.; Cruz, Á.B.; Pereira, D.H.; de Souza, N.L.G.D.; Carlos, T.D.; Soares, A.M.V.M.; Gravato, C.; Sarmento, R.A. Degradation of the Neonicotinoid Thiamethoxam by the Solar-Fenton Process: Theoretical Insights and Ecotoxicological Evaluation 2022. Available online: https://ssrn.com/abstract=4193674 (accessed on 30 September 2022).
- ASTM. Standard practice for acute toxicity testing with fish, macroinvertebrates and amphibians. In American Society for Testing and Materials; American Standards for Testing and Materials: Philadelphia, PA, USA, 1980; Volume 729, pp. 406–430. [Google Scholar]
- Dornelas, A.S.; Sarmento, R.; Saraiva, A.S.; Barbosa, R.S.; Vieira, M.M.; Gravato, C.; Soares, A.M. Effects of two biopesticides and salt on behaviour, regeneration and sexual reproduction of the freshwater planarian Girardia tigrina. J. Hazard. Mater. 2021, 404, 124089. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Haleem, D.R.; Genidy, N.A.; Fahmy, A.R.; Abu El Azm, F.S.M.; Ismail, N.S.M. Comparative Modelling, Toxicological and Biochemical Studies of Imidacloprid and Thiamethoxam Insecticides on the House Fly, Musca domestica L. (Diptera: Muscidae). Egypt. Acad. J. Biol. Sci. A Èntomol. 2018, 11, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Di Ilio, V.; Metwaly, N.; Saccardo, F.; Caprio, E. Adult and Egg Mortality of Rhynchophorus Ferrugineus Oliver (Coleoptera: Curculionidae) Induced By Thiamethoxam And Clothianidin. J. Agric. Vet. Sci. 2018, 11, 59–67. [Google Scholar] [CrossRef]
- Balanza, V.; Mendoza, J.E.; Bielza, P. Variation in susceptibility and selection for resistance to imidacloprid and thiamethoxam in Mediterranean populations of Orius laevigatus. Èntomol. Exp. Appl. 2019, 167, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Stuchi, A.L.P.B.; Moreira, D.R.; Galhardo, D.; dos Santos, S.A.; Ronqui, L.; Cantagalli, L.B.; Lopes, D.A.; Sinópolis-Gigliolli, A.A.; Toledo, V.D.A.A.D.; Ruvolo-Takasusuki, M.C.C. Comparative toxicity of fipronil, malathion, and thiamethoxam on the stingless bee Tetragonisca fiebrigi (Schwarz, 1938). Acta Sci. Biol. Sci. 2022, 44, e57846. [Google Scholar] [CrossRef]
- Mohamed, H.T.; Mohamed, I.A.; Abou-Elhagag, G.H.; Saba, R.M. Toxicity and field persistence of thiamethoxam and dinotefuran against cabbage aphid, Brevicoryne brassica L. (Homoptera: Aphididae) under laboratory and field conditions. J. Phyto-Pathol. Pest Manag. 2015, 2, 20–26. [Google Scholar]
- Amirzade, N.; Izadi, H.; Amin Jalali, M.; Zohdi, H. Evaluation of three neonicotinoid insecticides against the common pistachio psylla, Agonoscena pistaciae, and its natural enemies. J. Insect Sci. 2014, 14, 35. [Google Scholar] [CrossRef]
- Fouad, E.A.; Abotaleb, A.O. Sublethal Effects of Two Insecticides, Deltamethrin, Thiamethoxam and the Botanical Insecticide (Foeniculum vulgare Mill.) on Callosobruchus maculates (Fabr.) (Coleoptera: Bruchidae). Egypt. Acad. J. Biol. Sci. A Èntomol. 2021, 14, 255–269. [Google Scholar] [CrossRef]
- Riaz, M.A.; Chandor-Proust, A.; Dauphin-Villemant, C.; Poupardin, R.; Jones, C.M.; Strode, C.; Régent-Kloeckner, M.; David, J.-P.; Reynaud, S. Molecular mechanisms associated with increased tolerance to the neonicotinoid insecticide imidacloprid in the dengue vector Aedes aegypti. Aquat. Toxicol. 2013, 126, 326–337. [Google Scholar] [CrossRef]
- Raby, M.; Nowierski, M.; Perlov, D.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P. Acute toxicity of 6 neonicotinoid insecticides to freshwater invertebrates. Environ. Toxicol. Chem. 2018, 37, 1430–1445. [Google Scholar] [CrossRef] [PubMed]
- Surgan, M.; Condon, M.; Cox, C. Pesticide Risk Indicators: Unidentified Inert Ingredients Compromise Their Integrity and Utility. Environ. Manag. 2010, 45, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, J.J.; Sevilla-Morán, B.; Sandín-España, P.; López-Goti, C.; Alonso-Prados, J.L. Challenges of biopesticides under the European Regulation (EC) No. 1107/2009: An overview of new trends in residue analysis. Stud. Nat. Prod. Chem. 2014, 43, 437–482. [Google Scholar]
- Benchaâbane, S.; Ayad, A.; Loucif-Ayad, W.; Soltani, N. Multibiomarker responses after exposure to a sublethal concentration of thiamethoxam in the African honeybee (Apis mellifera intermissa). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 257, 109334. [Google Scholar] [CrossRef] [PubMed]
- Mullin, C.A. Effects of ‘inactive’ ingredients on bees. Curr. Opin. Insect Sci. 2015, 10, 194–200. [Google Scholar] [CrossRef]
- Straw, E.A. A Brief Note on Pollinator Exposure to Co-Formulants and Adjuvants. Environ. Toxicol. Chem. 2022, 41, 1584–1585. [Google Scholar] [CrossRef]
- Straw, E.A.; Brown, M.J.F. Co-formulant in a commercial fungicide product causes lethal and sub-lethal effects in bumble bees. Sci. Rep. 2021, 11, 21653. [Google Scholar] [CrossRef]
- Mullin, C.A.; Fine, J.D.; Reynolds, R.D.; Frazier, M.T. Toxicological Risks of Agrochemical Spray Adjuvants: Organosilicone Surfactants May Not Be Safe. Front. Public Health 2016, 4, 92. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Antoniou, M.N. Ignoring Adjuvant Toxicity Falsifies the Safety Profile of Commercial Pesticides. Front. Public Health 2018, 5, 361. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Bernay, B.; Séralini, G.-E. Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology 2013, 313, 122–128. [Google Scholar] [CrossRef]
- Jemec, A.; Tišler, T.; Drobne, D.; Sepčić, K.; Fournier, D.; Trebše, P. Comparative toxicity of imidacloprid, of its commercial liquid formulation and of diazinon to a non-target arthropod, the microcrustacean Daphnia magna. Chemosphere 2007, 68, 1408–1418. [Google Scholar] [CrossRef]
- Halappa, B.; Patil, R.K. Detoxifying enzyme studies on cotton leafhopper, Amrasca biguttula biguttula (Ishida), resistance to neonicotinoid insecticides in field populations in Karnataka, India. J. Plant Prot. Res. 2016, 56, 346–352. [Google Scholar] [CrossRef]
- Uğurlu, P.; Ünlü, E.; Satar, E.I. The toxicological effects of thiamethoxam on Gammarus kischineffensis (Schellenberg 1937) (Crustacea: Amphipoda). Environ. Toxicol. Pharmacol. 2015, 39, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Matsumura, F. Synergistic Actions of Formamidine Insecticides on the Activity of Pyrethroids and Neonicotinoids Against Aedes aegypti (Diptera: Culicidae). J. Med. Èntomol. 2012, 49, 1405–1410. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, A.J.; Hedges, A.M.; Intini, K.D.; Brown, L.R.; Maisonneuve, F.J.; Robinson, S.A.; Gillis, P.L.; de Solla, S.R. Acute and chronic toxicity of neonicotinoid and butenolide insecticides to the freshwater amphipod, Hyalella azteca. Ecotoxicol. Environ. Saf. 2019, 175, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, A.J.; Hedges, A.M.; Intini, K.D.; Brown, L.R.; Maisonneuve, F.J.; Robinson, S.A.; Gillis, P.L.; de Solla, S.R. Lethal and sublethal toxicity of neonicotinoid and butenolide insecticides to the mayfly, Hexagenia spp. Environ. Pollut. 2018, 238, 63–75. [Google Scholar] [CrossRef]
- de M Ribeiro, M.G.P.; Hunt, T.E.; Siegfried, B.D. Acute-Contact and Chronic-Systemic In Vivo Bioassays: Regional Monitoring of Susceptibility to Thiamethoxam in Soybean Aphid (Hemiptera: Aphididae) Populations from the North Central United States. J. Econ. Èntomol. 2018, 111, 337–347. [Google Scholar] [CrossRef]
- Fu, Z.; Han, F.; Huang, K.; Zhang, J.; Qin, J.G.; Chen, L.; Li, E. Combined toxic effects of thiamethoxam on intestinal flora, transcriptome and physiology of Pacific white shrimp Litopenaeus vannamei. Sci. Total. Environ. 2022, 830, 154799. [Google Scholar] [CrossRef]
- Cavallaro, M.C.; Morrissey, C.A.; Headley, J.V.; Peru, K.M.; Liber, K. Comparative chronic toxicity of imidacloprid, clothianidin, and thiamethoxam to Chironomus dilutus and estimation of toxic equivalency factors. Environ. Toxicol. Chem. 2016, 36, 372–382. [Google Scholar] [CrossRef]
- Prosser, R.; de Solla, S.; Holman, E.; Osborne, R.; Robinson, S.; Bartlett, A.; Maisonneuve, F.; Gillis, P. Sensitivity of the early-life stages of freshwater mollusks to neonicotinoid and butenolide insecticides. Environ. Pollut. 2016, 218, 428–435. [Google Scholar] [CrossRef]
- Stamm, M.D.; Baxendale, F.P.; Heng-Moss, T.M.; Siegfried, B.D.; Blankenship, E.E.; Gaussoin, R.E. Dose-response relationships of clothianidin, imidacloprid, and thiamethoxam to Blissus occiduus (Hemiptera: Blissidae). J. Econ. Èntomol. 2011, 104, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Thiamethoxam induces transgenerational hormesis effects and alteration of genes expression in Aphis gossypii. Pestic. Biochem. Physiol. 2020, 165, 104557. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbosa, R.S.; Rotili, E.A.; Santos, A.M.A.; Ribeiro, F.; Dornelas, A.S.P.; Pereira, D.H.; Cavallini, G.S.; Soares, A.M.V.M.; Sarmento, R.A.; Gravato, C. Eco(toxicological) Assessment of the Neonicotinoid Formulation Actara® Using Planarian Girardia tigrina as Model Organism. Pollutants 2022, 2, 546-555. https://doi.org/10.3390/pollutants2040035
Barbosa RS, Rotili EA, Santos AMA, Ribeiro F, Dornelas ASP, Pereira DH, Cavallini GS, Soares AMVM, Sarmento RA, Gravato C. Eco(toxicological) Assessment of the Neonicotinoid Formulation Actara® Using Planarian Girardia tigrina as Model Organism. Pollutants. 2022; 2(4):546-555. https://doi.org/10.3390/pollutants2040035
Chicago/Turabian StyleBarbosa, Rone S., Eliane Aparecida Rotili, Amanda Magda Almeida Santos, Fabianne Ribeiro, Aline Silvestre Pereira Dornelas, Douglas Henrique Pereira, Grasiele Soares Cavallini, Amadeu M. V. M. Soares, Renato Almeida Sarmento, and Carlos Gravato. 2022. "Eco(toxicological) Assessment of the Neonicotinoid Formulation Actara® Using Planarian Girardia tigrina as Model Organism" Pollutants 2, no. 4: 546-555. https://doi.org/10.3390/pollutants2040035
APA StyleBarbosa, R. S., Rotili, E. A., Santos, A. M. A., Ribeiro, F., Dornelas, A. S. P., Pereira, D. H., Cavallini, G. S., Soares, A. M. V. M., Sarmento, R. A., & Gravato, C. (2022). Eco(toxicological) Assessment of the Neonicotinoid Formulation Actara® Using Planarian Girardia tigrina as Model Organism. Pollutants, 2(4), 546-555. https://doi.org/10.3390/pollutants2040035