
Phytoplankton Community in Relation to Environmental Variables in the Tidal Mangrove Creeks of the Pasur River Estuary, Bangladesh

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
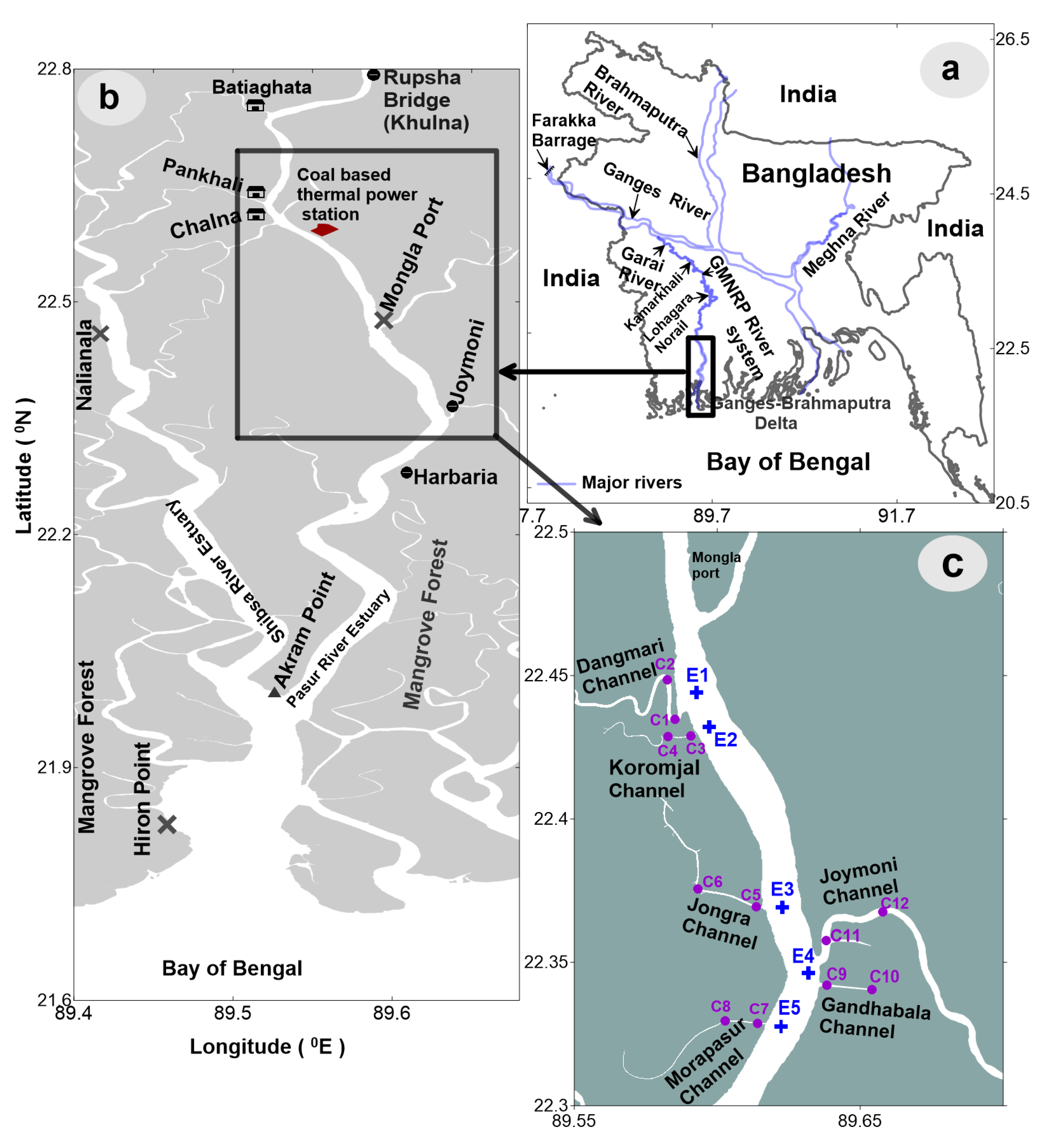
2.1. Study Area
2.2. Sample Collection and Laboratory Analysis
2.3. Statistical Analysis
3. Results
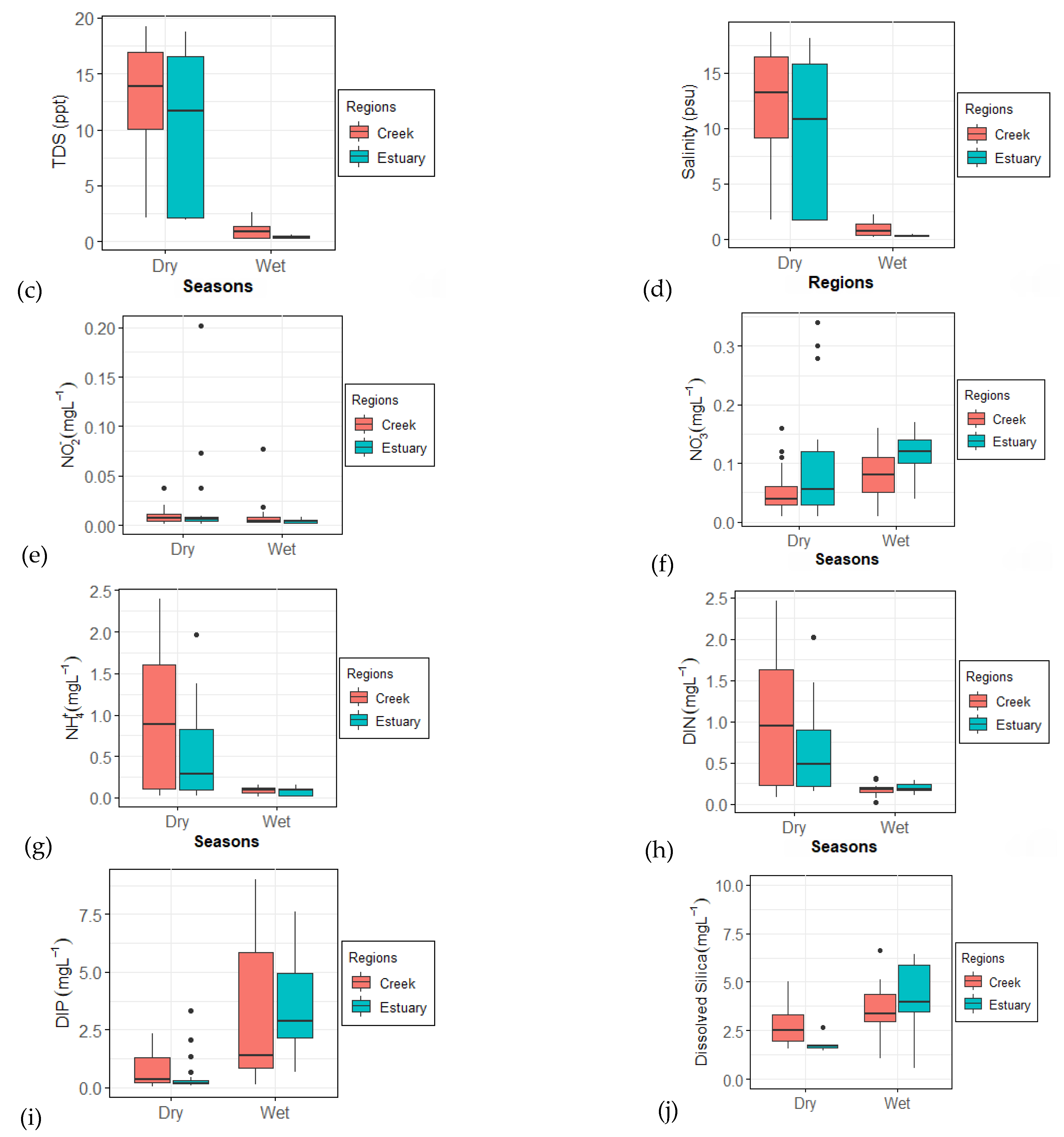
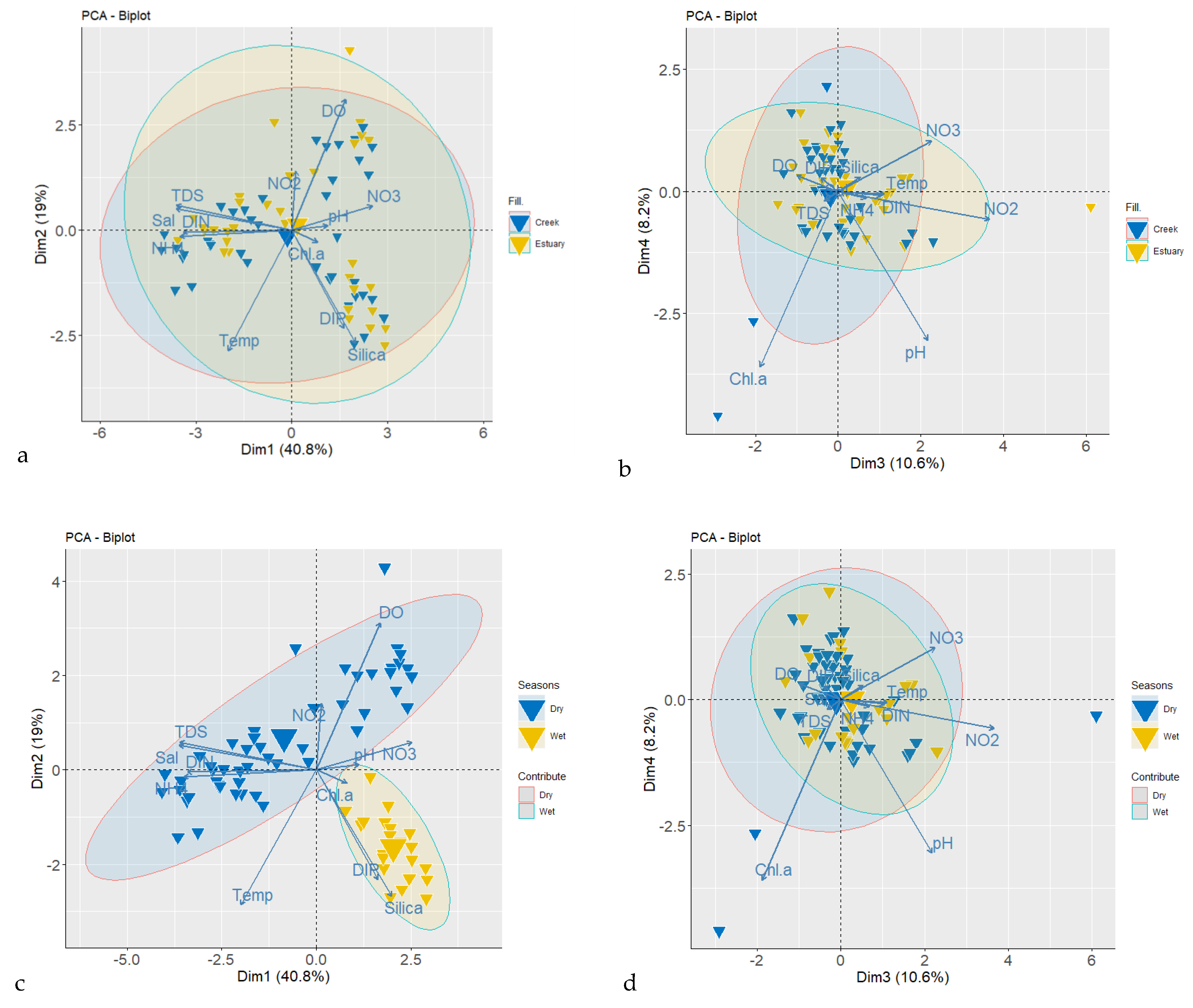
3.1. Physicochemical Parameters
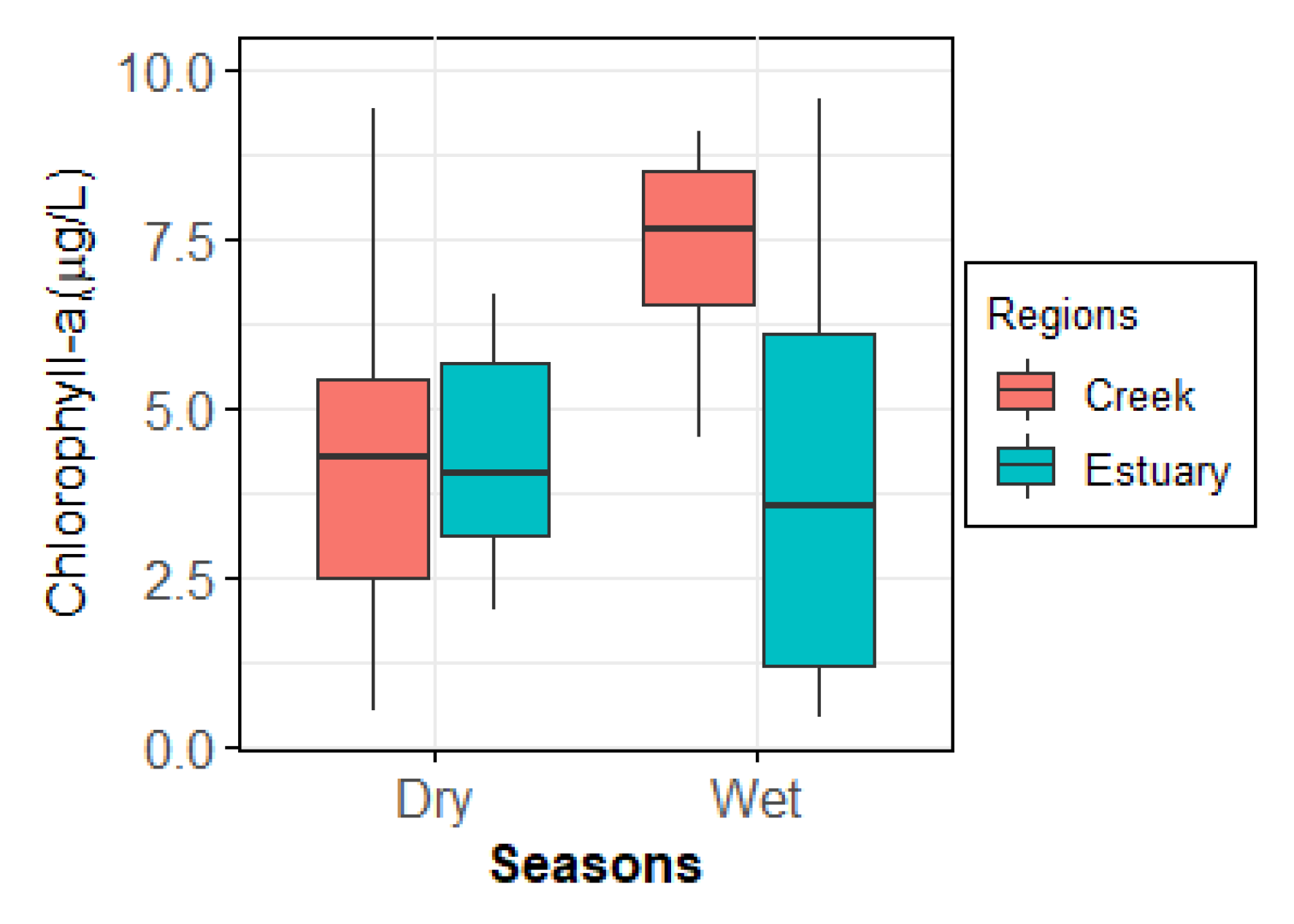
3.2. Chlorophyll a
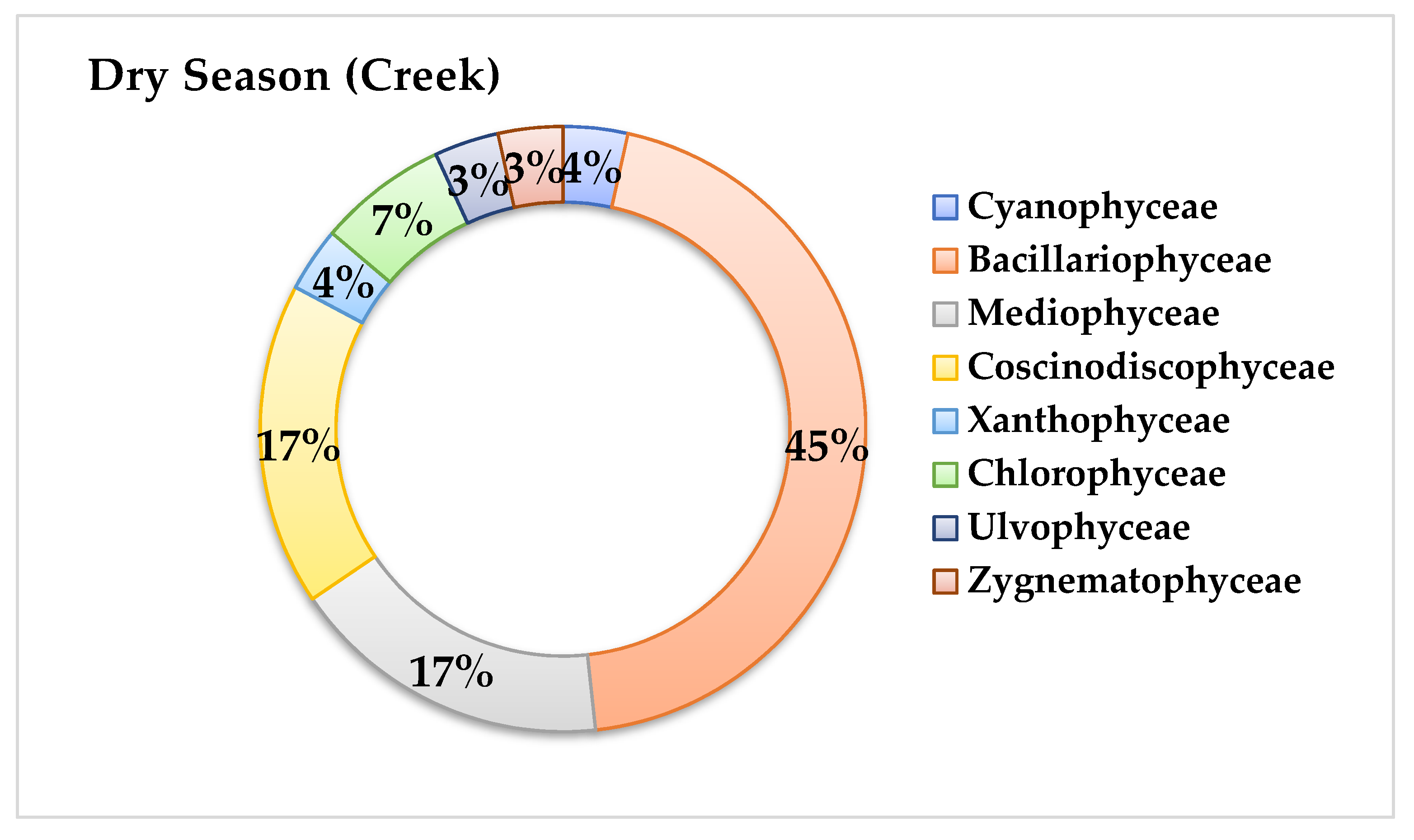
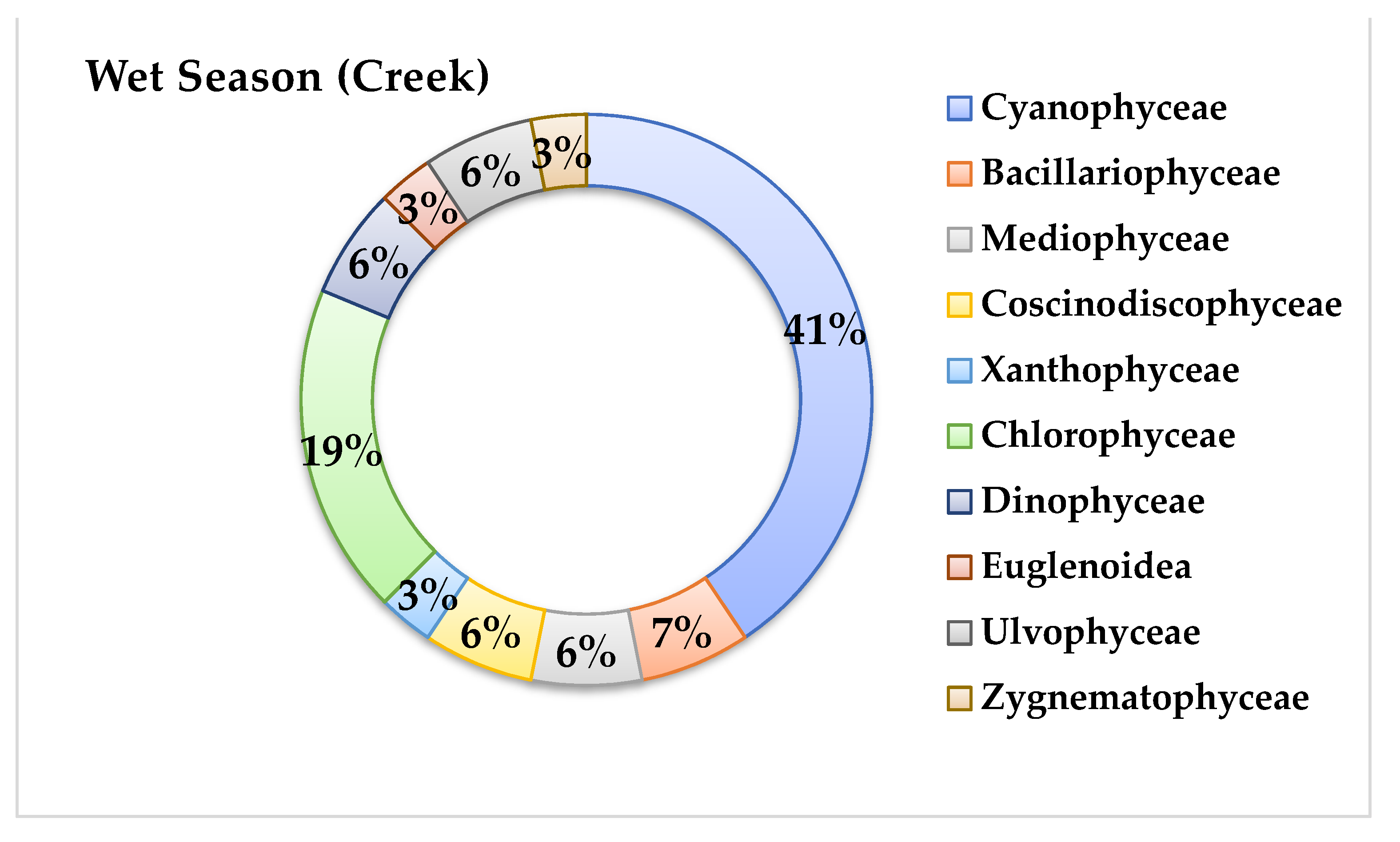
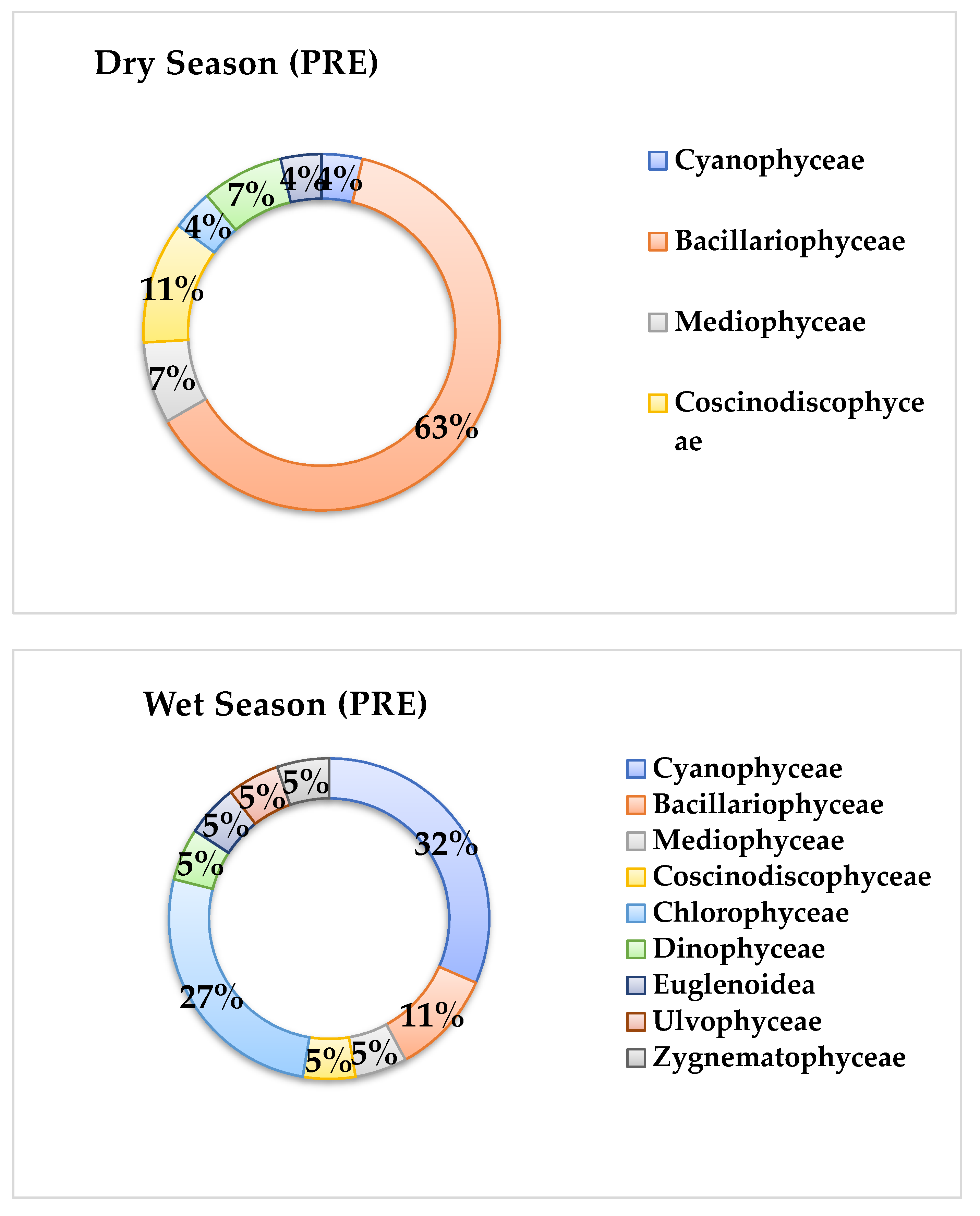
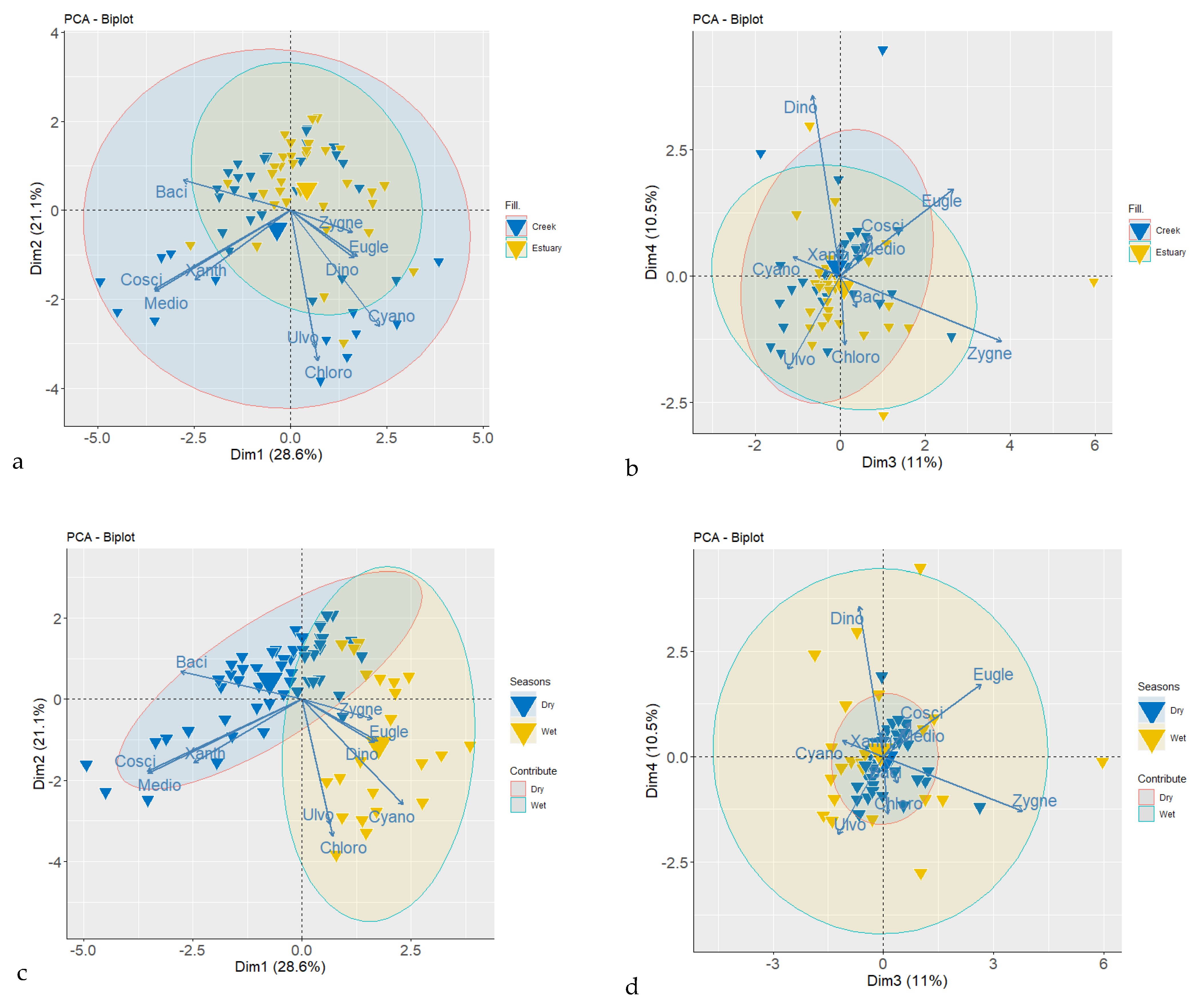
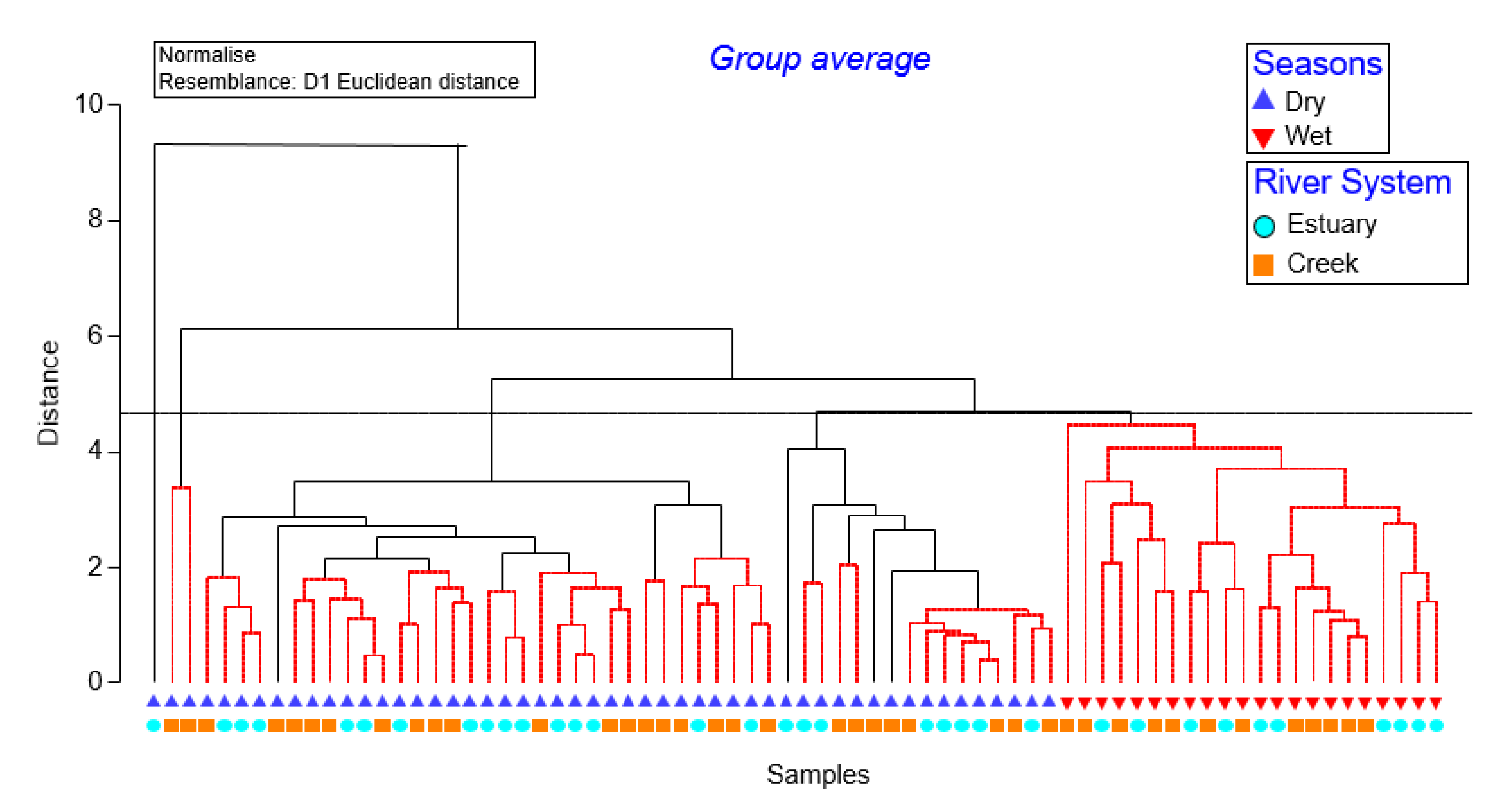
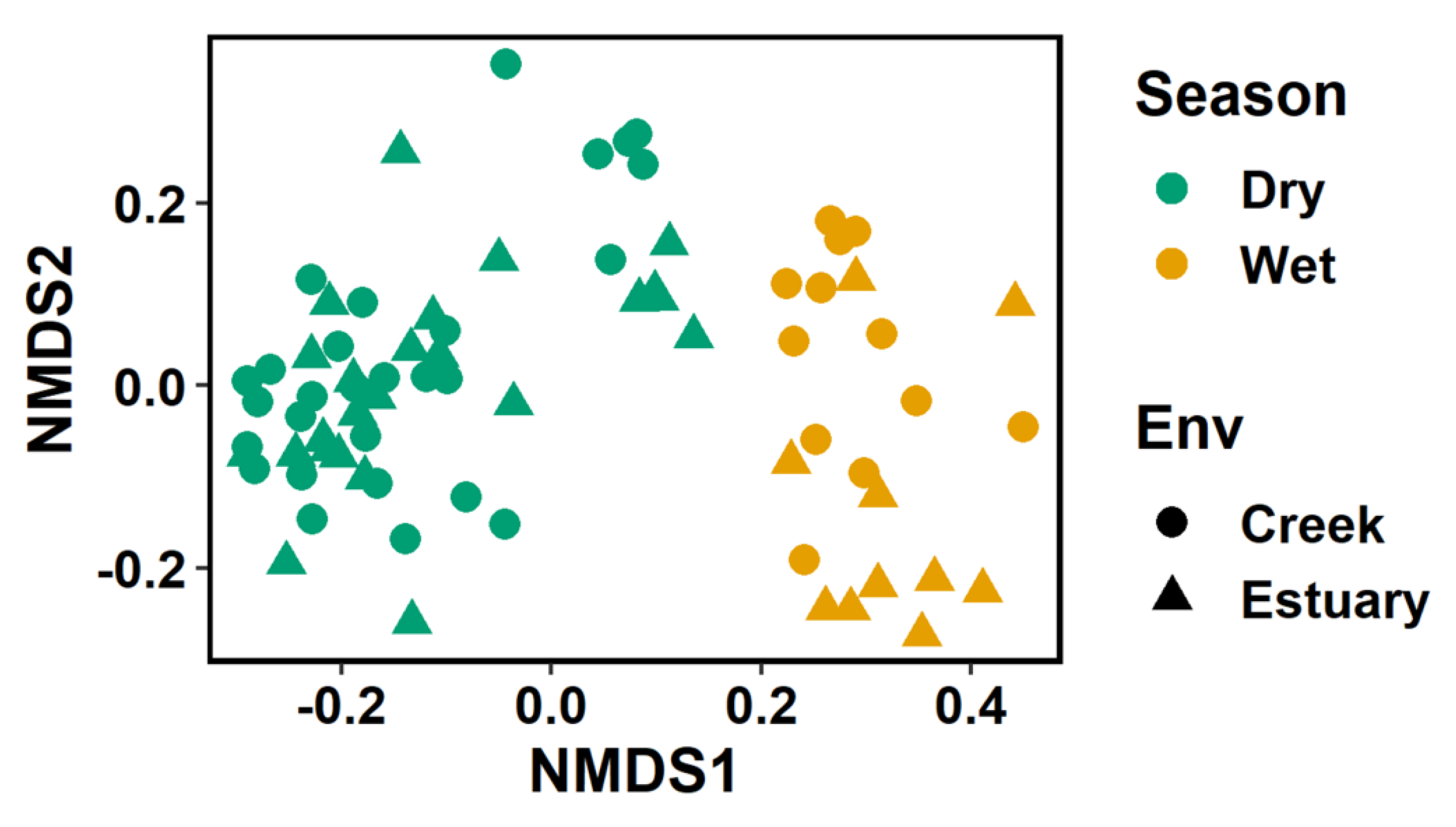
3.3. Phytoplankton Community Structure
3.4. Phytoplankton Species Diversity
4. Discussion
4.1. Physicochemical Parameters
4.2. Chlorophyll a
4.3. Phytoplankton Community Structure
4.4. Phytoplankton Species Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, W.M.; Pritchard, D.W. Estuaries in: MN Hill; The Sea John Wiley and Sons: New York, NY, USA, 1963; Volume 2, pp. 306–324. [Google Scholar]
- Cornils, A.; Niehoff, B.; Richter, C.; Al-Najjar, T.; Schnack-Schiel, S.B. Seasonal abundance and reproduction of clausocalanid copepods in the northern Gulf of Aqaba (Red Sea). J. Plankton Res. 2006, 29, 57–70. [Google Scholar] [CrossRef]
- Islam, S.; Ueda, H.; Tanaka, M. Spatial and seasonal variations in copepod communities related to turbidity maximum along the Chikugo estuarine gradient in the upper Ariake Bay, Japan. Estuar. Coast. Shelf Sci. 2006, 68, 113–126. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, M.; Liu, X.; Yu, H.; Otte, M.L.; Ma, C.; Her, Y.G. Environmental variables influencing phytoplankton com-munities in hydrologically connected aquatic habitats in the Lake Xingkai basin. Ecol. Indicat. 2018, 91, 1–12. [Google Scholar] [CrossRef]
- Hilaluddin, F.; Yusoff, F.; Toda, T. Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community. J. Mar. Sci. Eng. 2020, 8, 528. [Google Scholar] [CrossRef]
- Collins, N.R.; Williams, R. Zooplankton of the Bristol Channel and Severn Estuary. The distribution of four copepods in relation to salinity. Mar. Biol. 1981, 64, 273–283. [Google Scholar] [CrossRef]
- Bergström, A.-K.; Jonsson, A.; Isles, P.D.F.; Creed, I.F.; Lau, D.C.P. Changes in nutritional quality and nutrient limitation regimes of phytoplankton in response to declining N deposition in mountain lakes. Aquat. Sci. 2020, 82, 31. [Google Scholar] [CrossRef] [Green Version]
- Taipale, S.J.; Vuorio, K.; Aalto, S.L.; Peltomaa, E.; Tiirola, M. Eutrophication reduces the nutritional value of phytoplankton in boreal lakes. Environ. Res. 2019, 179, 108836. [Google Scholar] [CrossRef]
- Davies, O.A.; Abowei, J.F.; Tawari, C.C. Phytoplankton Community of Elechi Creek, Niger Delta, Nigeria-A Nutrient-Polluted Tropical Creek. Am. J. Appl. Sci. 2009, 6, 1143–1152. [Google Scholar] [CrossRef]
- Saravanakumar, A.; Rajkumar, M.; Serebiah, J.S.; A Thivakaran, G. Seasonal variations in physico-chemical characteristics of water, sediment and soil texture in arid zone mangroves of Kachchh-Gujarat. J. Environ. Biol. 2008, 29, 725–732. [Google Scholar]
- Hastuti, A.W.; Pancawati, Y.; Surana, I.N. The abundance and spatial distribution of plankton communities in Perancak Es-tuary, Bali. IOP Conf. Ser. Earth Environ. Sci. 2018, 176, 012042. [Google Scholar] [CrossRef]
- Dyer, K. Coastal and Estuarine Sediment Dynamics; John Wiley and Sons Chichester Sussex: London, UK, 1986; p. 358. [Google Scholar]
- Flynn, K.; Butler, I. Nitrogen sources for the growth of marine microalgae: Role of dissolved free amino acids. Mar. Ecol. Prog. Ser. 1986, 34, 281–304. [Google Scholar] [CrossRef]
- Valenzuela-Sanchez, C.G.; Pasten-Miranda, N.M.; Enriquez-Ocaña, L.F.; Barraza-Guardado, R.H.; Holguin, J.V.; Mar-tinez-Cordova, L.R. Phytoplankton composition and abundance as indicators of aquaculture effluents impact in coastal en-vironments of mid Gulf of California. Heliyon 2021, 7, e0620. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Das, R.; Vimexen, V. Seasonal phytoplankton succession in Netravathi–Gurupura estuary, Karnataka, India: Study on a three tier hydrographic platform. Estuar. Coast. Shelf Sci. 2020, 242, 106830. [Google Scholar]
- Lisitsyn, A.P. The marginal filter of the ocean. Oceanol. Rus. Acad. Sci. 1995, 34, 671–682. [Google Scholar]
- Gebhardt, A.; Schoster, F.; Gaye, B.; Beeskow, B.; Rachold, V.; Unger, D.; Ittekkot, V. The turbidity maximum zone of the Yenisei River (Siberia) and its impact on organic and inorganic proxies. Estuar. Coast. Shelf Sci. 2005, 65, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.N.; Gnauck, A. Water salinity investigation in the Sundarbans rivers in Bangladesh. Int. J. Water 2011, 6, 74. [Google Scholar] [CrossRef]
- Rahaman, S.M.B.; Golder, J.; Rahaman, M.S.; Hasanuzzaman, A.F.M.; Huq, K.A.; Begum, S.; Bir, J. Spatial and temporal variations in phytoplankton abundance and species diversity in the Sundarbans mangrove forest of Bangladesh. J. Mar. Sci. Res. Dev. 2013, 3, 1–9. [Google Scholar]
- Zinat, A.; Jewel, M.A.S.; Khatun, B.; Hasan, M.K.; Saleha, J.N. Seasonal variations of phytoplankton community structure in Pasur River estuary of Bangladesh. Int. J. Fish. Aquat. Stud. 2021, 9, 37–44. [Google Scholar]
- Uddin, M.N.; Tumpa, I.J.; Hossain, Z. Diversity of Biological Communities Along the Major Rivers of Sundarbans in Bangla-desh. Asian Fish. Sci. 2021, 34, 127–137. [Google Scholar]
- Shah, M.M.R.; Hossain, M.Y.; Begum, M.; Ahmed, Z.F.; Ohtomi, J.; Rahman, M.M.; Fulanda, B. Seasonal variations of phyto-planktonic community structure and production in relation to environmental factors of the southwest coastal waters of Bangladesh. J. Fish. Aquat. Sci. 2008, 392, 102–113. [Google Scholar]
- Chowdhury, M.A. Spatio-Temporal Variation of Plankton in the Pasur River Estuary. Master’s Thesis, Bangabandhu Sheikh Mujibur Rahman Agricultural University (BSMRAU), Gazipur, Bangladesh, 2019. [Google Scholar]
- Shefat, S.; Chowdhury, M.; Haque, F.; Hasan, J.; Salam, M.; Shaha, D. Assessment of Physico-Chemical Properties of the Pasur River Estuarine Water. Ann. Bangladesh Agric. 2021, 24, 1–16. [Google Scholar] [CrossRef]
- Aziz, A.; Rahman, M.; Ahmed, A. Diversity, Distribution and Density of Estuarine Phytoplankton in The Sundarban Mangrove Forests, Bangladesh. Bangladesh J. Bot. 2012, 41, 87–95. [Google Scholar] [CrossRef]
- Rahman, M.R.; Asaduzzaman, M. Ecology of Sundarban, Bangladesh. J. Sci. Found. 2010, 8, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Hach Company. Water Analysis Handbook Hach Company, 7th ed.; HACH Company: Loveland, CO, USA, 2012. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998; p. 1268. [Google Scholar]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A manual of chemical & biological methods for seawater analysis. Mar. Pollut. Bull. 1984, 1, 101–112. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. In World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2022. [Google Scholar]
- Stirling, H.P. Chemical and Biological Methods of Water Analysis for Aquaculturalists; Institute of Aquaculture, University of Stirling: Stirling, UK, 1985; pp. 118–119. [Google Scholar]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2017, 34, 1600–1602. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.C.; Bates, D.J.; DebRoy, S.; Sakar, D. The Nlme Package: Linear and Nonlinear Mixed Effects Models; R Version 3; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Statist. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Hu, M.; Wang, C.; Liu, Y.; Zhang, X.; Jian, S. Fish species composition, distribution and community structure in the lower reaches of Ganjiang River, Jiangxi, China. Sci. Rep. 2019, 9, 10100. [Google Scholar] [CrossRef]
- Jorgensen, S.E.; Xu, F.L.; Salas, F.; Marques, J.C. Application of indicators for the assessment of ecosystem health. In Handbook of Ecological Indicators for Assessment of Ecosystem Health; Taylor and Francis: Abingdon, UK, 2005. [Google Scholar]
- Abowei, J.F.N. Salinity, Dissolved Oxygen, pH and Surface Water Temperature Conditions in Nkoro River, Niger Delta, Ni-geria. Adv. J. Food Sci. Technol. 2010, 2, 36–40. [Google Scholar]
- Gadhia, M.; Surana, R.; Ansari, E. Seasonal Variations in Physico-Chemical Characterstics of Tapi Estuary in Hazira Industrial Area. Our Nat. 2013, 10, 249–257. [Google Scholar] [CrossRef] [Green Version]
- McLusky, D.S. The Estuarine Ecosystem, 2nd ed.; Chapman and Hall: New York, NY, USA, 1989. [Google Scholar]
- Tait, R.V. Elements of Marine Ecology: An Introductory Course, 3rd ed.; Butterworth-Heinemann: Oxford, UK, 1981. [Google Scholar]
- Plinski, M.; Jozwiak, T. Temperature and N: P ratio as factors causing blooms of blue-green algae in the Gulf of Gdansk. Oceanologia 1999, 41, 73–80. [Google Scholar]
- Fatema, K.; Maznah, W.O.W.; Isa, M.M. Spatial variation of water quality parameters in a mangrove estuary. Int. J. Environ. Sci. Technol. 2014, 12, 2091–2102. [Google Scholar] [CrossRef] [Green Version]
- Saifullah, A.; Kamal, A.H.M.; Idris, M.H.; Rajaee, A.H.; Alam Bhuiyan, K. Phytoplankton in tropical mangrove estuaries: Role and interdependency. For. Sci. Technol. 2015, 12, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.S.R.; Rao, V.R. Seasonal variation in temperature and salinity in the Gautami-Godavari estuary. Proc. Indian Acad. Sci.-Earth Planet. Sci. 1994, 103, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yang, C.; Wen, C.; Wen, G. S-type Dissolved Oxygen Distribution along Water Depth in a Canyon-shaped and Algae Blooming Water Source Reservoir: Reasons and Control. Int. J. Environ. Res. Public Health 2019, 16, 987. [Google Scholar] [CrossRef] [Green Version]
- Badran, M.I. Dissolved oxygen, chlorophyll-a and nutrients: Seasonal cycles in waters of the Gulf of Aquaba, Red Sea. Aquatic Ecosyst. Health Manag. 2001, 4, 139–150. [Google Scholar] [CrossRef]
- Pearce, M.W.; Schumann, E.H. Dissolved Oxygen Characteristics of the Gamtoos Estuary, South Africa. Afr. J. Mar. Sci. 2003, 25, 99–109. [Google Scholar] [CrossRef]
- Valiela, I.; Bowen, J.L. Nitrogen sources to watersheds and estuaries: Role of land cover mosaics and losses within water-sheds. Environ. Pollut. 2002, 118, 239–248. [Google Scholar] [CrossRef]
- Tanaka, K.; Choo, P.-S. Influences of Nutrient Outwelling from the Mangrove Swamp on the Distribution of Phytoplankton in the Matang Mangrove Estuary, Malaysia. J. Oceanogr. 2000, 56, 69–78. [Google Scholar] [CrossRef]
- Boney, A.D. Seasonal studies on the phytoplankton and primary production in the inner Firth of Clyde. Proc. R. Soc. Edinb. Sec. B Biol. Sci. 1986, 90, 203–222. [Google Scholar] [CrossRef]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled biogeochemical cycles: Eutrophi-cation and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Caraco, N.F.; Cole, J.J.; Likens, G.E. Evidence for sulphate-controlled phosphorus release from sediments of aquatic sys-tems. Nature 1989, 341, 316–318. [Google Scholar] [CrossRef]
- Pamplona, F.C.; Paes, E.T.; Nepomuceno, A. Nutrient fluctuations in the Quatipuru river: A macrotidal estuarine mangrove system in the Brazilian Amazonian basin. Estuar. Coast. Shelf. Sci. 2013, 133, 273–284. [Google Scholar] [CrossRef]
- Sospedra, J.; Niencheski, L.F.H.; Falco, S.; Andrade, C.F.; Attisano, K.K.; Rodilla, M. Identifying the main sources of silicate in coastal waters of the Southern Gulf of Valencia (Western Mediterranean Sea). Oceanologia 2018, 60, 52–64. [Google Scholar] [CrossRef]
- Steele, J.H. Environmental Control of Photosynthesis in the Sea. Limnol. Oceanogr. 1962, 7, 137–150. [Google Scholar] [CrossRef]
- Yao, P.; Yu, Z.; Deng, C.; Liu, S.; Zhen, Y. Spatial–temporal distribution of phytoplankton pigments in relation to nutrient status in Jiaozhou Bay, China. Estuar. Coast. Shelf. Sci. 2010, 89, 234–244. [Google Scholar] [CrossRef]
- Kumari, P.V.; Jayappa, K.S.; Thomas, S.; Joshi, M. Impact of south west monsoon on the variability of chlorophyll a in the coastal waters off Mangalore to Malpe, West Coast India. Int. J. Adv. Res. Sci. Eng. 2017, 6, 1264–1270. [Google Scholar]
- Robarts, R.D.; Zohary, T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. N. Z. J. Marine Freshwater Res. 1987, 21, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Taylor and Francis: London, UK, 2021; p. 858. [Google Scholar] [CrossRef]
- Suthers, I.; Rissik, D.; Richardson, A. Plankton: A guide to Their Ecology and Monitoring for Water Quality; CSIRO Publishing: Clayton, Australia, 2019. [Google Scholar]
- Winder, J.A.; Cheng, D.M. Quantification of Factors Controlling the Development of Anabaena Circinalis Blooms; Urban Water Research Association of Australia: Melbourne, Australia, 1995. [Google Scholar]
- Matsubara, T.; Nagasoe, S.; Yamasaki, Y.; Shikata, T.; Shimasaki, Y.; Oshima, Y.; Honjo, T. Effects of temperature, salinity, and irradiance on the growth of the dinoflagellate Akashiwo sanguinea. J. Exp. Mar. Biol. Ecol. 2007, 342, 226–230. [Google Scholar] [CrossRef]
- Srinivas, L.; Seeta, Y.; Reddy, P.M. Bacillariophyceae as Ecological Indicators of Water Quality in Manair Dam, Karimnagar, India. IJSRT 2018, 4, 468–474. [Google Scholar]
- Gamier, J.; Billen, G.; Coste, M. Seasonal succession of diatoms and Chlorophyceae in the drainage network of the Seine River: Observation and modeling. Limnol. Oceanogr. 1995, 40, 750–765. [Google Scholar] [CrossRef]
- Kumar, M.R.; Vishnu, S.R.; Sudhanandh, V.S.; Faisal, A.K.; Shibu, R.; Vimexen, V.; Krishnan, A.K. Proliferation of dinoflag-ellates in Kochi estuary, Kerala. J. Environ. Biol. 2014, 35, 877. [Google Scholar] [PubMed]
- Iqbal, M.M.; Masum, B.M.; Nurul, H.M.; Shafiqul, I.M.; Rajib, P.H.; Khurshid, A.B.M.; Dawood, M.A. Seasonal distribution of phytoplankton community in a subtropical estuary of the south-eastern coast of Bangladesh. Zool. Ecol. 2017, 27, 304–310. [Google Scholar] [CrossRef]
- Margalef, D.R. Information theory in ecology. Mem. Real Acad. Cienc. Y Artes Barc. 1957, 32, 374–559. [Google Scholar]
- Gogoi, P.; Sinha, A.; Das Sarkar, S.; Chanu, T.N.; Yadav, A.K.; Koushlesh, S.K.; Borah, S.; Das, S.K.; Das, B.K. Seasonal influence of physicochemical parameters on phytoplankton diversity and assemblage pattern in Kailash Khal, a tropical wetland, Sundarbans, India. Appl. Water Sci. 2019, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science Ltd., Blackwell Publishing Company: Oxford, UK, 2004. [Google Scholar]
- Yang, B.; Jiang, Y.-J.; He, W.; Liu, W.-X.; Kong, X.-Z.; Jørgensen, S.E.; Xu, F.-L. The tempo-spatial variations of phytoplankton diversities and their correlation with trophic state levels in a large eutrophic Chinese lake. Ecol. Indic. 2016, 66, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Li, Z.; Li, L.; Lu, F.; Liu, Y.; Lu, X.; Fan, Y. Phytoplankton alpha diversity indices response the trophic state variation in hydrologically connected aquatic habitats in the Harbin Section of the Songhua River. Sci. Rep. 2020, 10, 21337. [Google Scholar] [CrossRef]
- Rajkumar, M.; Perumal, P.; Ashok, P.V.; Vengadesh, P.N.; Thillai, R.K. Phytoplankton diversity in Pichavaram mangrove waters from south-east coast of India. J. Environ. Biol. 2009, 30, 489–498. [Google Scholar]
- Knowlton, M.F.; Jones, J.R. Connectivity Influences Temporal Variation of Limnological Conditions in Missouri River Scour Lakes. Lake Reserv. Manag. 2003, 19, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Stefanidou, N.; Katsiapi, M.; Tsianis, D.; Demertzioglou, M.; Michaloudi, E.; Moustaka-Gouni, M. Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons. Diversity 2020, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Chu, X. Quantification of the spatio-temporal variations in hydrologic connectivity of small-scale topographic surfaces under various rainfall conditions. J. Hydrol. 2013, 505, 65–77. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Creeks | Estuary vs. Creeks | |||||||
|---|---|---|---|---|---|---|---|---|
| Stations | Seasons (Dry and Wet) | Stations | Seasons (Dry and Wet) | |||||
| F | p | F | p | F | p | F | p | |
| Temperature | 0.079 | 0.995 | 1.953 | 0.169 | 1.303 | 0.257 | 6.126 | 0.015 |
| Salinity | 0.585 | 0.712 | 367.421 | 0.000 | 0.172 | 0.679 | 731.852 | 0.000 |
| DO | 0.585 | 0.712 | 0.130 | 0.720 | 3.720 | 0.057 | 2.704 | 0.103 |
| pH | 1.589 | 0.183 | 0.812 | 0.372 | 3.364 | 0.070 | 3.675 | 0.058 |
| Nitrate | 0.664 | 0.652 | 22.016 | 0.000 | 9.199 | 0.003 | 43.629 | 0.000 |
| Nitrite | 0.494 | 0.779 | 0.000 | 0.996 | 0.594 | 0.443 | 2.983 | 0.087 |
| Ammonia | 0.813 | 0.547 | 47.047 | 0.000 | 3.465 | 0.066 | 82.945 | 0.000 |
| Phosphate | 0.395 | 0.850 | 8.385 | 0.006 | 0.928 | 0.338 | 13.610 | 0.000 |
| TDS | 0.660 | 0.656 | 422.152 | 0.000 | 0.069 | 0.793 | 831.264 | 0.000 |
| DIN | 0.814 | 0.546 | 43.214 | 0.000 | 2.297 | 0.133 | 72.760 | 0.000 |
| Chlorophyll a | 0.978 | 0.442 | 1.058 | 0.309 | 7.799 | 0.006 | 0.714 | 0.400 |
| Silica | 7.336 | 0.014 | 22.86 | 0.000 | 104.89 | 0.000 | 341.64 | 0.000 |
| Dry Season (Non-Monsoon) (n = 25) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ST | Temp (°C) | DO (mg/L) | pH | Sal (psu) | TDS (ppt) | Nitrate (mg/L) | Nitrite (mg/L) | Ammonia (mg/L) | DIN (mg/L) | DIP (mg/L) | Chlorophyll a (µg/L) | Silica (mg/L) |
| E1 | 26.07 ± 2.59 31.30 − 20.4 | 6.98 ± 0.61 8.23 − 5.35 | 7.75 ± 0.08 7.91 − 7.49 | 11.57 ± 3.22 17.84 − 1.77 | 12.43 ± 3.33 18.66 − 2.17 | 0.009 ± 0.002 0.02 − 0.001 | 0.048 ± 0.02 0.13 − 0.01 | 0.61 ± 0.25 1.18 − 0.03 | 0.67 ± 0.25 1.22 − 0.17 | 0.20 ± 0.025 0.29 − 0.14 | 3.99 ± 0.77 6.48 − 2.8 | 2.65 ± 0.05 2.80 − 2.51 |
| E2 | 25.97 ± 2.47 31.09 − 20.8 | 6.88 ± 0.61 8.5 − 5.48 | 7.69 ± 0.7 7.87 − 7.47 | 11.64 ± 3.25 17.67 − 1.68 | 12.36 ± 3.30 18.35 − 2.08 | 0.005 ± 0.001 0.01 − 0.001 | 0.064 ± 0.02 0.14 − 0.02 | 0.81 ± 0.25 1.97 − 0.12 | 0.87 ± 0.35 2.02 − 0.22 | 0.22 ± 0.06 0.43 − 0.12 | 4.004 ± 0.69 6.19 − 2.35 | 1.48 ± 0.03 1.57 − 1.39 |
| E3 | 26.27 ± 2.60 31.61 − 20.8 | 6.73 ± 0.58 8.02 − 5.3 | 7.62 ± 0.12 7.94 − 7.35 | 11.42 ± 3.15 17.35 − 1.66 | 12.18 ± 3.25 18.05 − 2.03 | 0.005 ± 0.001 0.01 − 0.001 | 0.1 ± 0.03 0.28 − 0.02 | 0.44 ± 0.15 0.78 − 0.13 | 0.54 ± 0.12 0.82 − 0.24 | 0.17 ± 0.02 0.23 − 0.12 | 4.004 ± 0.72 6.29 − 2.28 | 1.78 ± 0.041 1.89 − 1.67 |
| E4 | 26.53 ± 2.5 31.4 − 21 | 6.78 ± 0.45 8.12 − 5.76 | 7.69 ± 0.11 7.93 − 7.42 | 10.94 ± 3.03 16.12 − 1.72 | 11.89 ± 3.09 17.15 − 2.09 | 0.01 ± 0.007 0.04 − 0.001 | 0.07 ± 0.02 0.14 − 0.02 | 0.74 ± 0.25 1.38 − 0.1 | 0.82 ± 0.25 1.47 − 0.24 | 0.45 ± 0.10 1.34 − 0.12 | 4.64 ± 0.71 6.69 − 3.07 | 1.59 ± 0.02 1.65 − 1.5 |
| E5 | 26.49 ± 2.57 31.94 − 20.5 | 6.73 ± 0.5 8.11 − 5.4 | 7.66 ± 0.08 7.85 − 7.43 | 10.83 ± 2.89 15.64 − 1.71 | 11.95 ± 3.02 16.66 − 2.1 | 0.006 ± 0.0015 0.01 − 0.001 | 0.05 ± 0.015 0.12 − 0.02 | 0.57 ± 0.18 0.98 − 0.03 | 0.63 ± 0.17 1.01 − 0.16 | 0.22 ± 0.08 0.5 − 0.1 | 4.13 ± 0.54 5.79 − 3.1 | 1.71 ± 0.025 1.79 − 1.63 |
| Wet season (monsoon) (n = 10) | ||||||||||||
| ST | Temp (°C) | DO (mg/L) | pH | Sal (psu) | TDS (ppt) | Nitrate (mg/L) | Nitrite (mg/L) | Ammonia (mg/L) | DIN (mg/L) | DIP (mg/L) | Chlorophyll a (µg/L) | Silica (mg/L) |
| E1 | 30.35 ± 0.38 30.9 − 29.8 | 6.04 ± 0.06 6.13 − 5.96 | 8 ± 0.045 8.07 − 7.93 | 0.21 ± 0.015 0.23 − 0.18 | 0.38 ± 0.09 0.51 − 0.25 | 0.002 ± 0.00035 0.003 − 0.001 | 0.16 ± 0.007 0.17 − 0.15 | 0.06 ± 0.02 0.11 − 0.01 | 0.22 ± 0.04 0.28 − 0.16 | 3.4 ± 0.77 4.5 − 2.3 | 3.65 ± 1.27 6.16 − 1.14 | 5.43 ± 0.03 5.47 − 5.38 |
| E2 | 30.35 ± 0.53 31.1 − 29.6 | 5.95 ± 0.13 6.15 − 5.76 | 7.74 ± 0.20 8.03 − 7.46 | 0.27 ± 0.06 0.36 − 0.19 | 0.36 ± 0.075 0.47 − 0.25 | 0.006 ± 0.001 0.01 − 0.001 | 0.14 ± 0.0005 0.14 − 0.12 | 0.06 ± 0.02 0.11 − 0.01 | 0.206 ± 0.03 0.25 − 0.16 | 3.25 ± 0.75 4.33 − 2.17 | 3.65 ± 1.13 5.96 − 1.34 | 3.59 ± 0.04 3.65 − 3.53 |
| E3 | 30.25 ± 0.6 31.1 − 29.4 | 6.05 ± 0.13 6.25 − 5.86 | 7.95 ± 0.08 8.08 − 7.83 | 0.28 ± 0.014 0.3 − 0.26 | 0.42 ± 0.05 0.5 − 0.34 | 0.004 ± 0.0005 0.01 − 0.001 | 0.085 ± 0.03 0.13 − 0.04 | 0.085 ± 0.03 0.13 − 0.04 | 0.174 ± 0.0005 0.18 − 0.16 | 4.65 ± 1.25 6.43 − 2.87 | 4.99 ± 1.23 9.56 − 0.42 | 6.38 ± 0.035 6.44 − 6.33 |
| E4 | 30.5 ± 0.77 31.6 − 29.4 | 5.97 ± 0.13 6.17 − 5.78 | 7.84 ± 0.14 8.05 − 7.63 | 0.28 ± 0.05 0.36 − 0.21 | 0.38 ± 0.07 0.49 − 0.28 | 0.005 ± 0.0005 0.01 − 0.001 | 0.09 ± 0.015 0.12 − 0.07 | 0.1 ± 0.00005 0.1 − 0.001 | 0.2 ± 0.007 0.23 − 0.17 | 4.6 ± 1.58 7.6 − 1.71 | 8.12 ± 2.15 15.46 − 0.79 | 1.86 ± 0.96 3.23 − 0.5 |
| E5 | 30.7 ± 1.13 32.3 − 29.1 | 6.13 ± 0.13 6.32 − 5.95 | 7.63 ± 0.26 8.01 − 7.26 | 0.83 ± 0.26 1.2 − 0.46 | 0.46 ± 0.10 0.61 − 0.31 | 0.003 ± 0.001 0.01 − 0.001 | 0.1 ± 0.0005 0.11 − 0.09 | 0.05 ± 0.015 0.1 − 0.01 | 0.15 ± 0.035 0.22 − 0.1 | 1.39 ± 0.52 2.13 − 0.66 | 5.19 ± 0.73 6.24 − 4.16 | 5.11 ± 0.81 6.27 − 3.96 |
| Dry Season (Non-Monsoon) (n = 30) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ST | Temp (°C) | DO (mg/L) | pH | Sal (psu) | TDS (ppt) | Nitrate (mg/L) | Nitrite (mg/L) | Ammonia (mg/L) | DIN (mg/L) | DIP (mg/L) | Chlorophyll a (µg/L) | Silica (mg/L) |
| C1, C2 | 26.13 ± 2.75 31.9 − 20.2 | 6.78 ± 0.6 8.03 − 5.32 | 7.74 ± 0.09 7.98 − 7.53 | 11.56 ± 3.23 17.84 − 1.75 | 12.23 ± 3.23 18.45 − 2.14 | 0.009 ± 0.0015 0.01 − 0.001 | 0.05 ± 0.009 0.08 − 0.04 | 0.96 ± 0.27 2.11 − 0.07 | 1.03 ± 0.45 2.15 − 0.14 | 0.99 ± 0.37 2.1 − 0.06 | 3.51 ± 0.59 4.84 − 2.13 | 5.01 ± 0.05 5.16 − 4.86 |
| C3, C4 | 26.04 ± 2.9 32.3 − 19.4 | 6.42 ± 0.69 8.23 − 4.62 | 7.63 ± 0.08 7.8 − 7.4 | 12.52 ± 3.06 18.66 − 3.59 | 11.45 ± 2.75 19.2 − 4.22 | 0.014 ± 0.004 0.02 − 0.001 | 0.05 ± 0.021 0.12 − 0.02 | 1.16 ± 0.45 2.17 − 0.02 | 1.23 ± 0.44 2.22 − 0.12 | 0.66 ± 0.23 1.31 − 0.11 | 5.08 ± 1.91 11.05 − 0.78 | 1.55 ± 0.05 1.71 − 1.41 |
| C5, C6 | 26.06 ± 2.71 31.2 − 20.1 | 6.35 ± 0.56 8.08 − 5.01 | 7.61 ± 0.07 7.81 − 7.5 | 11.73 ± 3.05 17.55 − 2.63 | 12.47 ± 3.07 18.23 − 3.15 | 0.006 ± 0.0015 0.01 − 0.001 | 0.044 ± 0.01 0.12 − 0.01 | 1.05 ± 0.28 1.61 − 0.08 | 1.11 ± 0.27 1.63 − 0.21 | 0.64 ± 0.23 1.26 − 0.16 | 4.98 ± 1.07 7.44 − 2.49 | 1.99 ± 0.05 2.14 − 1.85 |
| C7, C8 | 26.51 ± 2.62 31.7 − 20.65 | 6.70 ± 0.05 8.18 − 5.51 | 7.64 ± 0.07 7.77 − 7.48 | 11.12 ± 2.64 15.49 − 3.0 | 11.68 ± 2.88 16.16 − 2.43 | 0.004 ± 0.001 0.01 − 0.001 | 0.05 ± 0.01 0.1 − 0.03 | 0.51 ± 0.13 0.75 − 0.05 | 0.56 ± 0.12 0.80 − 0.15 | 0.55 ± 0.20 1.29 − 0.13 | 4.73 ± 1.37 9.41 − 2.17 | 3.41 ± 0.05 3.56 − 3.26 |
| C9, C10 | 26.66 ± 2.72 32.5 − 21 | 7.18 ± 0.69 8.77 − 5.76 | 7.62 ± 0.12 7.85 − 7.3 | 11.12 ± 3.05 16.48 − 1.71 | 11.85 ± 3.10 17.14 − 2.12 | 0.006 ± 0.001 0.01 − 0.001 | 0.06 ± 0.015 0.11 − 0.03 | 0.86 ± 0.25 2.32 − 0.09 | 0.93 ± 0.35 2.36 − 0.21 | 0.69 ± 0.22 1.97 − 0.17 | 9.84 ± 1.71 33.26 − 0.5 | 1.94 ± 0.05 2.1 − 1.82 |
| C11, C12 | 26.77 ± 2.81 32.95 − 20.7 | 6.95 ± 0.5 8.26 − 5.78 | 7.70 ± 0.09 7.88 − 7.47 | 11.35 ± 3.08 16.59 − 1.75 | 11.95 ± 3.1 17.32 − 2.14 | 0.010 ± 0.004 0.03 − 0.001 | 0.07 ± 0.025 0.16 − 0.03 | 0.73 ± 0.30 1.33 − 0.08 | 0.81 ± 0.25 1.41 − 0.12 | 0.47 ± 0.18 1.3 − 0.17 | 6.69 ± 2.52 15.61 − 3.55 | 3.07 ± 0.05 3.22 − 2.92 |
| Wet season (monsoon) (n = 12) | ||||||||||||
| ST | Temp (°C) | DO (mg/L) | pH | Sal (psu) | TDS (ppt) | Nitrate (mg/L) | Nitrite (mg/L) | Ammonia (mg/L) | DIN (mg/L) | DIP (mg/L) | Chlorophyll a (µg/L) | Silica (mg/L) |
| C1, C2 | 29.95 ± 0.7 30.9 − 28.95 | 6.12 ± 0.16 6.35 − 5.88 | 7.83 ± 0.54 7.96 − 7.7 | 0.22 ± 0.02 0.26 − 0.18 | 0.28 ± 0.02 0.32 − 0.25 | 0.023 ± 0.01 0.04 − 0.001 | 0.12 ± 0.01 0.14 − 0.1 | 0.06 ± 0.015 0.1 − 0.02 | 0.19 ± 0.025 0.24 − 0.15 | 2.74 ± 0.72 4.47 − 1.01 | 7.35 ± 0.005 7.36 − 7.35 | 2.73 ± 1.18 4.4 − 1.06 |
| C3, C4 | 30.25 ± 0.45 30.9 − 29.6 | 5.97 ± 0.09 6.11 − 5.85 | 7.67 ± 0.15 7.89 − 7.46 | 0.55 ± 0.24 0.9 − 0.22 | 0.61 ± 0.24 0.96 − 0.27 | 0.004 ± 0.0003 0.006 − 0.002 | 0.057 ± 0.015 0.08 − 0.04 | 0.06 ± 0.015 0.09 − 0.04 | 0.12 ± 0.03 0.17 − 0.07 | 4.76 ± 1.08 8.44 − 1.1 | 9.73 ± 0.59 10.57 − 8.9 | 4.12 ± 0.70 5.11 − 3.12 |
| C5, C6 | 29.77 ± 0.8 30.9 − 28.65 | 5.73 ± 0.08 5.85 − 5.61 | 7.52 ± 0.25 7.87 − 7.17 | 0.97 ± 0.08 1.1 − 0.85 | 1.03 ± 0.26 1.41 − 0.66 | 0.007 ± 0.0001 0.01 − 0.01 | 0.08 ± 0.02 0.11 − 0.05 | 0.11 ± 0.01 0.13 − 0.09 | 0.19 ± 0.03 0.25 − 0.15 | 1.27 ± 0.22 1.6 − 0.95 | 7.57 ± 0.09 7.71 − 7.43 | 3.59 ± 0.04 3.66 − 3.53 |
| C7, C8 | 29.92 ± 1.6 32.2 − 27.65 | 5.69 ± 0.19 5.98 − 5.42 | 7.73 ± 0.04 7.8 − 7.67 | 1.36 ± 0.33 1.84 − 0.88 | 2.87 ± 0.40 3.45 − 2.3 | 0.009 ± 0.003 0.02 − 0.001 | 0.05 ± 0.02 0.1 − 0.02 | 0.08 ± 0.0005 0.09 − 0.06 | 0.15 ± 0.02 0.18 − 0.12 | 3.09 ± 1.39 6.04 − 0.16 | 9.04 ± 0.85 10.25 − 7.84 | 3.33 ± 0.64 4.25 − 2.42 |
| C9, C10 | 30.15 ± 0.95 31.5 − 28.8 | 6.13 ± 0.14 6.34 − 5.93 | 7.64 ± 0.12 7.82 − 7.47 | 0.54 ± 0.06 0.64 − 0.45 | 0.66 ± 0.06 0.76 − 0.56 | 0.006 ± 0.0015 0.01 − 0.001 | 0.09 ± 0.01 0.11 − 0.08 | 0.15 ± 0.0005 0.2 − 0.1 | 0.19 ± 0.005 0.21 − 0.18 | 2.57 ± 1.15 4.91 − 0.24 | 9.07 ± 0.8 10.2 − 7.95 | 4.78 ± 1.3 6.63 − 2.94 |
| C11, C12 | 29.17 ± 1.04 30.65 − 27.7 | 5.82 ± 0.22 6.13 − 5.52 | 7.75 ± 0.01 7.77 − 7.73 | 1.28 ± 0.15 1.5 − 1.06 | 0.41 ± 0.05 0.49 − 0.34 | 0.006 ± 0.002 0.01 − 0.001 | 0.08 ± 0.03 0.14 − 0.04 | 0.09 ± 0.02 0.14 − 0.06 | 0.19 ± 0.01 0.21 − 0.17 | 3.50 ± 1.55 6.27 − 0.75 | 11.41 ± 1.23 13.17 − 9.66 | 3.04 ± 0.02 3.07 − 3.01 |
| Taxon | Sampling Stations | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C1, C2 | E1 | C3, C4 | E2 | C5, C6 | E3 | C11, C12 | E4 | C9, C10 | C7, C8 | E5 | ||
| Cyanophyceae | Oscillatoria sp. | + | + | + | + | + | ||||||
| Aphanizomenon sp. | + | + | + | + | + | |||||||
| Gloeocapsa sp. | + | + | + | |||||||||
| Dolichospermum sp. | + | |||||||||||
| Merismopedium sp. | + | + | + | + | + | + | + | + | + | |||
| Microcystis sp. | + | + | ||||||||||
| Arthrospira sp. | + | |||||||||||
| Gomphosphaeria sp. | + | + | + | |||||||||
| Bacillariophyceae | Lioloma sp. | + | + | + | + | + | + | + | + | + | ||
| Pleorosigma sp. | + | + | ||||||||||
| Asterionella sp. | + | + | ||||||||||
| Fragilaria sp. | + | |||||||||||
| Diatoma sp. | + | |||||||||||
| Chaetoceros sp. | + | |||||||||||
| Surirella sp. | + | + | ||||||||||
| Thalassionema sp. | + | + | ||||||||||
| Nitzschia sp. | + | + | ||||||||||
| Navicula sp. | + | |||||||||||
| Synedra sp. | + | + | + | |||||||||
| Coscinonodiscophyceae | Coscinodiscus sp. | + | + | + | + | + | + | + | + | + | + | |
| Triceratium sp. | + | + | ||||||||||
| Melosira sp. | + | |||||||||||
| Mediophyceae | Ditylum sp. | + | + | + | + | + | ||||||
| Odontella sp. | + | |||||||||||
| Tropidoneis sp. | + | |||||||||||
| Xanthophyceae | Tribonema sp. | + | + | + | + | + | + | |||||
| Chlorophyceae | Stigeoclonium sp. | + | ||||||||||
| Hydrodictyon sp. | + | + | + | |||||||||
| Pediastrum sp. | + | + | ||||||||||
| Volvox sp. | + | |||||||||||
| Oedogonium sp. | + | + | + | + | + | |||||||
| Zygnematophyceae | Spirogyra sp. | + | + | + | + | + | ||||||
| Mougeotia sp. | + | + | ||||||||||
| Dinophyceae | Tripos sp. | + | ||||||||||
| Polykrikos sp. | + | |||||||||||
| Ulvophyceae | Cladophora sp. | + | ||||||||||
| Ulothrix sp. | + | |||||||||||
| Euglenophyceae | Euglena sp. | + | + | + | ||||||||
| Shannon–Weaver Index | Peilou Index | Margalef Index | Simpson Diversity Index | Simpson’s Reciprocal Index | ||
|---|---|---|---|---|---|---|
| PRE | Dry season | 2.82 | 0.8 | 6.32 | 0.1 | 9.81 |
| Wet season | 2.68 | 0.66 | 6.08 | 0.19 | 7.02 | |
| Creeks | Dry season | 2.91 | 0.86 | 6.66 | 0.07 | 11.08 |
| Wet season | 3.09 | 0.97 | 6.86 | 0.03 | 14.57 | |
| t-test | ||||||
| Dry × Wet season | p < 0.05 | p < 0.05 | p < 0.05 | p < 0.05 | p < 0.05 | |
| PRE × Creeks | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | |
| Avg. Dissimilarity | Discriminating Species (Average Dissimilarity and Contribution Percentage) |
|---|---|
| Dry vs. Wet (85.13) | Synedra sp. (3.90, 4.58%), Fragilaria sp. (3.73, 4.39%), Asterionella sp. (3.66, 4.30%), Pleorosigma sp. (3.57, 4.19%), Coscinodiscus sp. (3.55, 4.17%), Diatoma sp. (3.54, 4.16%), Lioloma sp. (3.25, 3.81%), Arthrospira sp. (3.18, 3.74%), Merismopedium sp. (3.17, 3.73%), Chaetoceros sp. (3.04, 3.58%), Ulothrix sp. (2.93, 3.44%), Microcystis sp. (2.85, 3.34%), Pediastrum sp. (2.82, 3.31%), Oedogonium sp. (2.80, 3.29%), Melosira sp. (2.67, 3.14%), Gomphosphaeria sp. (2.67, 3.14%), Triceratium sp. (2.63, 3.1%), Nitzschia sp. (2.50, 2.94%), Aphanizomenon sp. (2.28, 2.68%), Gloeocapsa sp. (2.22, 2.61%), Stigeoclonium sp. (2.22, 2.60%), Ditylum sp. (2.18, 2.56%), Oscillatoria sp. (2.17, 2.55%), Muogeotia sp. (2.05, 2.41%), Odontella sp. (1.81, 2.12%), Dolichospermum sp. (1.80, 2.12%), Spirogyra sp. (1.67, 1.97%), Hydrodictyon sp. (1.66, 1.95%), Volvox sp. (1.43, 1.68%), Tropidoneis sp. (1.28, 1.51%), Surirella sp. (1.27, 1.49%), Navicula sp. (1.26, 1.48%), Thalassionema sp. (1.23, 1.44%), Tribonema sp. (1.04, 1.22%), Cladophora sp. (0.37, 0.43%), Euglena sp. (0.29, 0.35%), Tripos sp. (0.14, 0.16%) and Polykrikos sp. (0.13, 0.16%). |
| Creek vs. Estuary (74.93) | Synedra sp. (3.89, 5.19%), Diatoma sp. (3.63, 4.85%), Asterionella sp. (3.48, 4.65%), Coscinodiscus sp. (3.46, 4.61%), Fragilaria sp. (3.39, 4.53%), Lioloma sp. (3.38, 4.52%), Pleorosigma sp. (3.20, 4.27%), Chaetoceros sp. (3.06, 4.09%), Nitzschia sp. (2.91, 3.89%), Triceratium sp. (2.78, 3.71%), Melosira sp. (2.75, 3.67%), Pediastrum sp. (2.48, 3.31%), Gomphosphaeria sp. (2.31, 3.09%), Ulothrix sp. (2.27, 3.03%), Arthrospira sp. (2.25, 3.01%), Ditylum sp. (2.18, 2.92%), Odontella sp. (2.14, 2.86%), Oedogonium sp. (2.02, 2.70%), Muogeotia sp. (1.99, 2.65%), Microcystis sp. (1.70, 2.27%), Merismopedium sp. (1.63, 2.18%), Stigeoclonium sp. (1.50, 2.0%), Hydrodictyon sp. (1.50, 2.0%), Surirella sp. (1.49, 1.98%), Navicula sp. (1.48, 1.97%), Tropidoneis sp. (1.43, 1.92%), Thalassionema sp. (1.39, 1.86%), Spirogyra sp. (1.25, 1.68%), Aphanizomenon sp. (1.24, 1.66%), Gloeocapsa sp. (1.23, 1.64%), Volvox sp. (1.22, 1.63%), Oscillatoria sp. (1.21, 1.61%), Tribonema sp. (1.11, 1.49%), Dolichospermum sp. (1.01, 1.35%), Cladophora sp. (0.36, 0.48%), Tripos sp. (0.18, 0.24%), Euglena sp. (0.17, 0.23%) and Polykrikos sp. (0.08, 0.12%). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, J.; Shaha, D.C.; Kundu, S.R.; Yusoff, F.M.; Cho, Y.-K.; Haque, F.; Salam, M.A.; Ahmed, S.; Wahab, M.A.; Ahmed, M.; et al. Phytoplankton Community in Relation to Environmental Variables in the Tidal Mangrove Creeks of the Pasur River Estuary, Bangladesh. Conservation 2022, 2, 587-612. https://doi.org/10.3390/conservation2040039
Hasan J, Shaha DC, Kundu SR, Yusoff FM, Cho Y-K, Haque F, Salam MA, Ahmed S, Wahab MA, Ahmed M, et al. Phytoplankton Community in Relation to Environmental Variables in the Tidal Mangrove Creeks of the Pasur River Estuary, Bangladesh. Conservation. 2022; 2(4):587-612. https://doi.org/10.3390/conservation2040039
Chicago/Turabian StyleHasan, Jahid, Dinesh Chandra Shaha, Sampa Rani Kundu, Fatimah Md Yusoff, Yang-Ki Cho, Farhana Haque, Mohammad Abdus Salam, Salman Ahmed, Md. Abdul Wahab, Minhaz Ahmed, and et al. 2022. "Phytoplankton Community in Relation to Environmental Variables in the Tidal Mangrove Creeks of the Pasur River Estuary, Bangladesh" Conservation 2, no. 4: 587-612. https://doi.org/10.3390/conservation2040039
APA StyleHasan, J., Shaha, D. C., Kundu, S. R., Yusoff, F. M., Cho, Y. -K., Haque, F., Salam, M. A., Ahmed, S., Wahab, M. A., Ahmed, M., Hossain, M. I., & Afrad, M. S. I. (2022). Phytoplankton Community in Relation to Environmental Variables in the Tidal Mangrove Creeks of the Pasur River Estuary, Bangladesh. Conservation, 2(4), 587-612. https://doi.org/10.3390/conservation2040039