


Impact of European Beaver (Castor fiber L.) on Vegetation Diversity in Protected Area River Valleys

Abstract
:1. Introduction
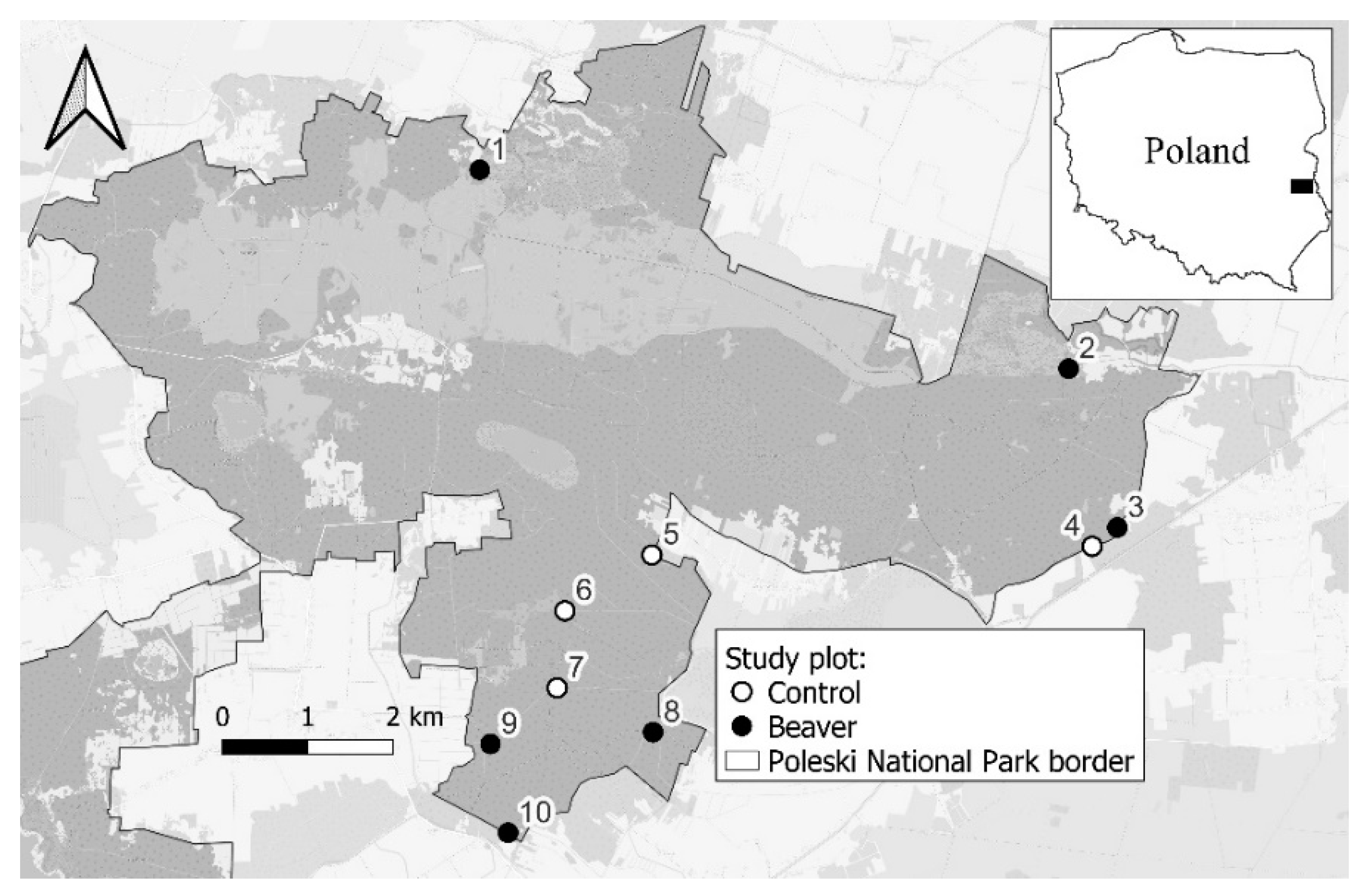
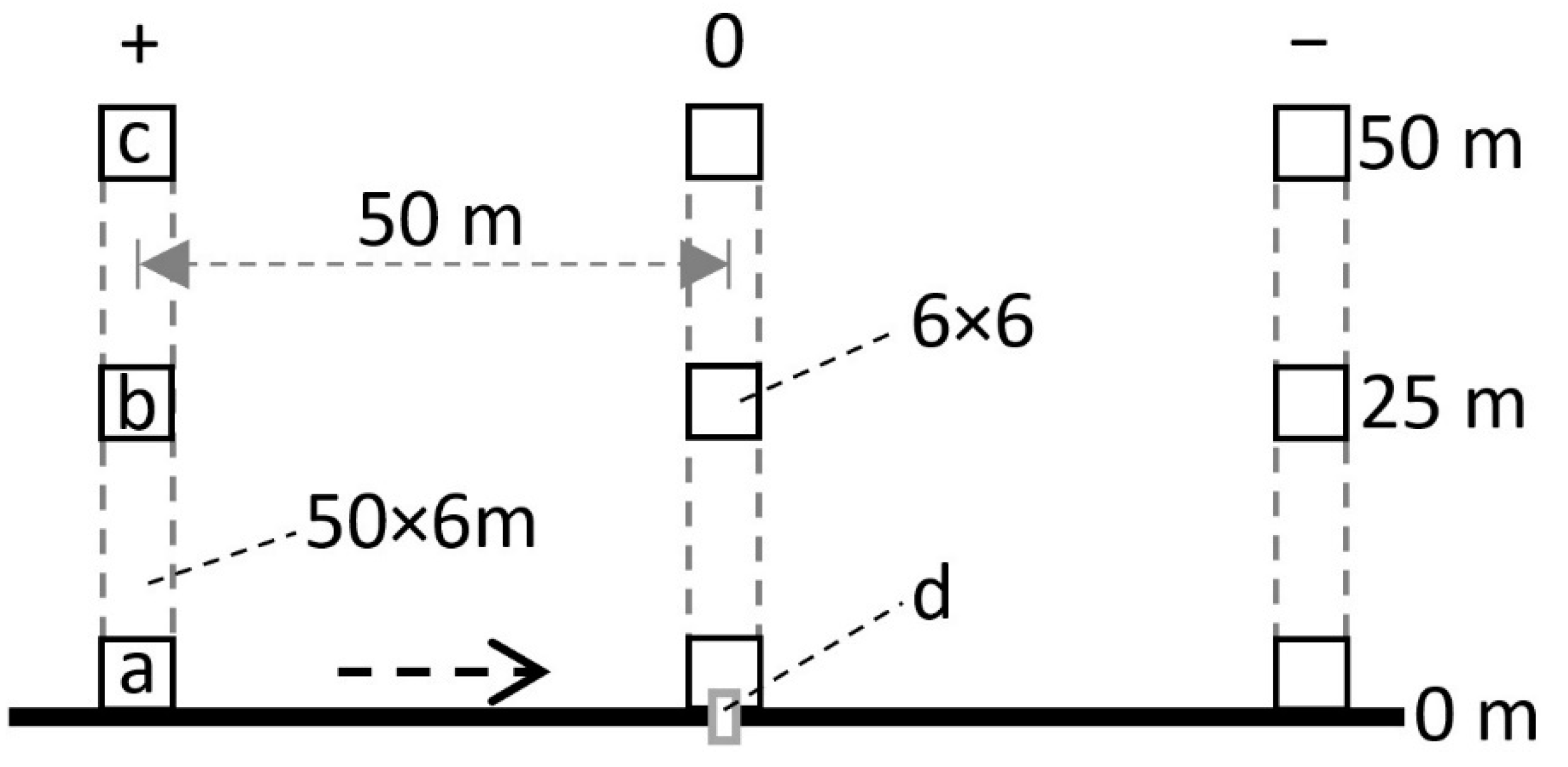
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
- The impact of beaver was limited to the immediate vicinity of watercourses. The farthest bite marks were recorded only 25 m from the axis of the watercourse.
- Beaver significantly increased species diversity in the tall-tree layer and in the forest floor layer, with no significant effects on the low tree and shrub layer.
- Five species of alien plants were found during the survey: Giant goldenrod, devil’s beggartick, small balsam, hedge bindweed, and black cherry. Except for small balsam, all the listed alien species were present in plots colonized by European beaver. Giant goldenrod, small balsam and hedge bindweed were found in comparison plots where beaver were not present.
- The alien species most strongly associated with beaver activity was devil’s beggartick, which occurred mainly in the vicinity of dams. Giant goldenrod benefited from the presence of beaver but did not require its presence for expansion. Black cherry occurred only in areas with dams and was not gnawed by beavers. It may benefit from the strong preference of beaver for native species.
- Our study confirmed hypotheses found in literature according to which beaver activities that reduce the proportion of native species can promote the expansion of alien species.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Species | Beaver Presence/Absence, Plot Number and Distance Zone | |||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| With Beaver | Without Beaver | With Beaver | ||||||||||||||||||||||||||||
| 1 | 2 | 3 1 | 4 | 5 | 6 | 7 | 8 | 9 | 10 2 | |||||||||||||||||||||
| a | b | c | a | b | c | a | b | c | a | b | c | a | b | c | a | b | c | a | b | c | a | b | c | a | b | c | a | b | c | |
| Layer I (trees taller than 10 m) | ||||||||||||||||||||||||||||||
| Black alder | = | = | – | = | ± | ± | ± | = | + | = | – | + | + | + | – | – | = | = | + | – | – | |||||||||
| Silver birch | = | = | – | – | – | – | – | + | – | – | – | – | – | = | = | – | – | – | ||||||||||||
| Scots pine | – | – | = | = | = | = | – | – | – | |||||||||||||||||||||
| Pedunculate oak | – | – | = | = | – | |||||||||||||||||||||||||
| Common aspen | – | – | – | – | – | – | – | |||||||||||||||||||||||
| Norway spruce | = | = | ||||||||||||||||||||||||||||
| Norway maple | – | |||||||||||||||||||||||||||||
| Layer II (trees shorter than 10 m and shrubs) | ||||||||||||||||||||||||||||||
| Black alder | = | – | – | = | – | – | – | |||||||||||||||||||||||
| Silver birch | – | – | – | – | – | – | – | |||||||||||||||||||||||
| Scots pine | – | |||||||||||||||||||||||||||||
| Pedunculate oak | – | – | – | – | = | |||||||||||||||||||||||||
| Common aspen | = | – | – | – | – | |||||||||||||||||||||||||
| Norway spruce | ||||||||||||||||||||||||||||||
| Norway maple | – | – | ||||||||||||||||||||||||||||
| Common hornbeam | – | – | – | + | ||||||||||||||||||||||||||
| Common ash | – | – | ||||||||||||||||||||||||||||
| Mountain ash | – | – | – | – | – | – | – | – | ||||||||||||||||||||||
| Bird cherry | – | ± | – | – | – | – | = | – | – | – | – | = | – | – | – | – | – | |||||||||||||
| Black cherry | – | – | + | – | ||||||||||||||||||||||||||
| Common hazel | = | – | – | = | = | – | + | = | – | |||||||||||||||||||||
| Alder buckthorn | – | = | – | – | – | = | – | – | – | + | = | – | – | – | – | – | – | – | ||||||||||||
| Willows | – | – | – | – | – | – | – | – | – | |||||||||||||||||||||
| Common dogwood | – | – | – | |||||||||||||||||||||||||||
| European spindle | – | |||||||||||||||||||||||||||||
| Common hawthorn | – | |||||||||||||||||||||||||||||
| Guelder rose | – | – | ||||||||||||||||||||||||||||

References
- Andersson, E.; Nilsson, C.; Johansson, M.E. Plant dispersal in boreal rivers and its relation to the diversity of riparian flora. J. Biogeogr. 2001, 27, 1095–1106. [Google Scholar] [CrossRef]
- Merritt, D.M.; Wohl, E.E. Plant dispersal along rivers fragmented by dams. River Res. Appl. 2006, 22, 1–26. [Google Scholar] [CrossRef]
- Dajdok, Z.; Tokarska-Guzik, B. Doliny rzeczne i wody stojące jako siedliska gatunków inwazyjnych. In Inwazyjne Gatunki Roślin Ekosystemów Mokradłowych Polski; Dajdok, Z., Pawlaczyk, P., Eds.; Wydawnictwo Klubu Przyrodników: Świebodzin, Poland, 2009; pp. 24–31. [Google Scholar]
- Kołaczkowska, E. Obce inwazyjne gatunki roślin w krajobrazie dolin Świdra i Rządzy [Invasive alien plant species in the landscape of Świder and Rządza valleys]. Pr. Kom. Kraj. Kult. 2010, 13, 152–163. [Google Scholar]
- Burkart, M. River corridor plants (Stromtalpflanzen) in Central European lowland: A review of a poorly understood plant distribution pattern. Glob. Ecol. Biogeogr. 2001, 10, 449–468. [Google Scholar] [CrossRef]
- Uziębło, A.; Ciapała, S. Zróżnicowanie florystyczne i ekologiczne roślinności przypotokowej w masywie Babiej Góry [Floristical and ecological differentiation of stream-bank vegetation in the Babia Góra massif]. Ochr. Beskidów Zach. 2006, 1, 93–103. [Google Scholar]
- Kehr, J.M.; Merritt, D.M.; Stromberg, J.C. Linkages between primary seed dispersal, hydrochory and flood timing in a semi- arid region river. J. Veg. Sci. 2014, 25, 287–300. [Google Scholar] [CrossRef]
- Wąsowicz, P. Migracje roślin naczyniowych w obrębie systemu rzecznego Wisły. [Vascular plant migrations within the Vistula river basin]. In Wisła Jako Warsztat Badawczy Biologów; Matyjasiak, P., Romanowski, J., Eds.; UKSW: Warsaw, Poland, 2011. [Google Scholar]
- Andersson, E.; Nilsson, C.; Johansson, M.E. Effects of river fragmentation on plant dispersal and riparian flora. Regul. Rivers Res. Manag. Int. J. Devoted River Res. Manag. 2000, 16, 83–89. [Google Scholar] [CrossRef]
- Rood, J.; Braatne, J.H.; Goater, L.A. Favorable fragmentation: River reservoirs can impede downstream expansion of riparian weeds. Ecol. Appl. 2010, 20, 1664–1677. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.L.; Thomas, C.C.; Cubley, E.C.; Clausen, A.J.; Shafroth, P.B. Does large dam removal restore downstream riparian vegetation diversity? Testing predictions on the Elwha River, Washington, USA. Ecol. Appl. 2022, 32, e2591. [Google Scholar] [CrossRef]
- Kowarik, I.; Säumel, I. Water dispersal as an additional pathway to invasions by the primarily wind-dispersed tree Ailanthus altissima. Plant Ecol. 2008, 198, 241–252. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warsaw, Poland, 2012; pp. 1–197. [Google Scholar]
- Foxcroft, L.C.; Pyšek, P.; Richardson, D.M.; Pergl, J.; Hulme, P.E. The bottom line: Impacts of alien plant invasions in protected areas. In Plant Invasions in Protected Areas: Patterns, Problems, and Challenges; Foxcroft, L.C., Pyšek, P., Richardson, D.M., Genovesi, P., Eds.; Springer: New York, NY, USA, 2013; pp. 19–41. [Google Scholar] [CrossRef]
- Zając, I.; Jaśkiewicz, K.; Jędrzejewska, E. Rozmieszczenie stanowisk inwazyjnych gatunków roślin naczyniowych w Suwalskim Parku Krajobrazowym. Fragm. Florist. Geobot. Polon 2015, 22, 65–78. [Google Scholar]
- Foxcroft, L.C.; Pyšek, P.; Richardson, D.M.; Genovesi, P.; MacFadyen, S. Plant invasion science in protected areas: Progress and priorities. Biol. Invasions 2017, 19, 1353–1378. [Google Scholar] [CrossRef]
- Bomanowska, A.; Adamowski, W.; Kirpluk, I.; Otręba, A.; Rewicz, A. Invasive alien plants in Polish national parks-threats to species diversity. PeerJ 2019, 7, e8034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foxcroft, L.C.; Jarošík, V.; Pyšek, P.; Richardson, D.M.; Rouget, M. Protected area boundaries as filters of plant invasions. Conserv. Biol. 2011, 25, 400–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wróbel, D. Zróżnicowanie fitocenotyczne wybranych gatunków inwazyjnych w dolinach rzecznych Karpat i Kotliny Sandomierskiej [Phytocoenotic differentiation of some invasive species in the river valleys of the Carpathians and the Sandomierz Basin]. Fragm. Florist. Geobot. Polon 2015, 22, 47–63. [Google Scholar]
- Wróbel, D. Występowanie roślin inwazyjnych w obrębie budowli i powierzchni utwardzonych w dolinach rzecznych karpat i kotliny sandomierskiej. Czas. Inżynierii Lądowej Sr. Archit. [JCEEA] 2017, XXXIV, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Koczur, A. Rozprzestrzenianie się rodzimych gatunków roślin niżowych i kenofitów w wyniku zabudowy hydrotechnicznej rzeki Czarny Dunajec. Fragm. Flor. Geobot. Polonica 2006, 13, 77–87. [Google Scholar]
- Wallem, P.K.; Anderson, C.B.; Martinez-Pastur, G.; Lencinas, M.V. Using assembly rules to measure the resilience of riparian plant communities to beaver invasion in subantarctic forests. Biol. Invasions 2010, 12, 325–335. [Google Scholar] [CrossRef]
- Janiszewski, P.; Hanzal, V.; Misiukiewicz, W. The Eurasian Beaver (Castor fiber) as a Keystone Species. Balt. For. 2014, 20, 277–286. [Google Scholar]
- Johnston, C.A. Vegetation of Beaver Impoundments. In Beavers: Boreal Ecosystem Engineers; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; Chapter 6; pp. 137–176. [Google Scholar] [CrossRef]
- Janczak, B.; Zieliński, J. Wybrane aspekty biologii nasion inwazyjnego terofita Impatiens glandulifera Royle (Balsaminaceae) [Selected aspects of seed biology of invasive therophyte Impatiens glandulifera Royle (Balsaminaceae)]. Stud. Mater. Cent. Edukac. Przyr.-Leśnej 2012, 14, 226–233. [Google Scholar]
- Stringer, A.P.; Gaywood, M.J. The impacts of beavers Castor spp. on biodiversity and the ecological basis for their reintroduction to Scotland, UK. Mammal Rev. 2016, 46, 270–283. [Google Scholar] [CrossRef]
- Juhász, A.; Katona, K.; Molnár, Z.; Hahn, I.; Biró, M. Reintroduced ecosystem engineer species may exacerbate ongoing biological invasion: Selective foraging of the Eurasian beaver in floodplains. Glob. Ecol. Conserv. 2020, 24, e01383. [Google Scholar] [CrossRef]
- Lesica, P.; Miles, S. Beavers indirectly enhance the growth of russian olive and tamarisk along eastern Montana rivers. West. N. Am. Nat. 2004, 64, 93–100. [Google Scholar]
- Juhász, A.; Bede-Fazekas, Á.; Katona, K.; Molnár, Z.; Biró, M. Foraging decisions with conservation consequences: Interaction between beavers and invasive tree species. Ecol. Evol. 2022, 12, e8899. [Google Scholar] [CrossRef]
- Kimball, B.A.; Perry, K.R. Manipulating Beaver (Castor canadensis) Feeding Responses to Invasive Tamarisk (Tamarix spp.). J. Chem. Ecol. 2008, 34, 1050–1056. [Google Scholar] [CrossRef]
- Mięsak-Wójcik, K.; Turczyński, M.; Sposób, J. Natural and anthropogenic changes of standing water bodies in west Polesie (East Poland). In Proceedings of the 2nd International Conference—Water Resources and Wetlands, Tulcea, Romania, 11–13 September 2014; Available online: http://www.limnology.ro/water2014/proceedings.html (accessed on 15 March 2022).
- Omelczuk, Ł.; Kozieł, M. Wpływ działalności bobra europejskiego Castor fiber L. na zmiany stosunków wodnych na Polesiu Zachodnim. Bad. Fizjogr. 2017, 68, 99–113. [Google Scholar] [CrossRef]
- Polesie National Park Website. Available online: http://www.poleskipn.pl/index.php/historia-i-przyroda/ochrona-przyrody/169-gatunki-inwazyjne (accessed on 23 August 2022).
- Shimano, K. Analysis of the Relationship between DBH and Crown Projection Area Using a New Model. J. For. Res. 1997, 2, 237–242. [Google Scholar] [CrossRef]
- Lockgart, R.B.; Weih, R.C., Jr.; Smith, K.M. Crown radius and Diameter at Breast height relationships for six bottomland hardwood species. J. Ark. Acad. Sci. 2005, 59, 110–115. [Google Scholar]
- Sousa, A.M.O.; Gonçalves, A.C.; da Silva, J.R.M. Above-Ground Biomass Estimation with High Spatial Resolution Satellite Images. In Biomass Volume Estimation and Valorization for Energy; IntechOpen: London, UK, 2017; Volume 3, pp. 47–70. [Google Scholar] [CrossRef] [Green Version]
- Iizuka, K.; Kosugi, Y.; Noguchi, S.; Iwagami, S. Toward a comprehensive model for estimating diameter at breast height of Japanese cypress (Chamaecyparis obtusa) using crown size derived from unmanned aerial systems. Comput. Electron. Agric. 2022, 192, 106579. [Google Scholar] [CrossRef]
- Nagendra, H. Opposite trends in response for the Shannon and Simpson indices of landscape diversity. Appl. Geogr. 2002, 22, 175–186. [Google Scholar] [CrossRef]
- Sienkiewicz, J. Koncepcje bioróżnorodności—Ich wymiary i miary w świetle literatury. Ochr. Śr. Zasobów Nat. 2010, 45, 7–29. [Google Scholar]
- Lasota, J.; Wiechec, M.; Blonska, E.; Brożek, S. Wybrane wskaźniki różnorodności biologicznej na tle cech utworów glebowych w wyżynnym borze jodłowym Abietetum albae. Leśne Pr. Badaw. 2017, 78, 120–128. [Google Scholar]
- Drescher, A.; Prots, B. Distribution patterns of Himalayan balsam (Impatiens glandulifera Royle) in Austria. Phytogeogr. Probl. Synanthropic Plants 2003, 11, 137–146. [Google Scholar]
- Kasperek, G. Fluctuations in numbers of neophytes, especially Impatiens glandulifera, in permanent plots in a west German floodplain during 13 years. In Biological Invasions: Challenges for Science; Kühn, I., Klotz, S., Eds.; Neobiota: Sofia, Bulgaria, 2022; Volume 3; pp. 27–37. [Google Scholar]
- Tokarska-Guzik, B. The Establishment and Spread of Alien Plant Species (Kenophytes) in the Flora of Poland; Prace Naukowe Uniwersytetu Śląskiego w Katowicach; Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2005; Volume 2372, pp. 79–82. [Google Scholar]
- Śliwiński, M. Occurrence of anthropophytes along streams of the Sowie Mountains and Dzierżoniów Basin (South-Western Poland) in dependence on land use. Biodiv. Res. Conserv. 2008, 11–12, 33–40. [Google Scholar]


| Index | Equation | Legend |
|---|---|---|
| Margalef (R) | S—number of species ni—area occupied by plants of particular species N—area occupied by plants of all species | |
| Shannon-Wiener (H) | ||
| Simpson (C) |
| Species | Share of Species in the Number of Bite Marks (Average Distance of Bite Marks from the River Edge) | With Beaver | Without Beaver | ||
|---|---|---|---|---|---|
| Layer 1 | |||||
| I | II | I | II | ||
| % (m) | % | % | % | % | |
| Black alder | 11.6 (10.0) | 48.3 | 3.1 | 49.8 | 19.9 |
| Silver birch | 26.1 (4.1) | 16.1 | 3.9 | 23.1 | 5.4 |
| Scots pine | - | 10.7 | 1.7 | 18.1 | 0.0 |
| Pedunculate oak | - | 7.9 | 5.8 | 7.6 | 0.6 |
| Common aspen | 14.5 (4.3) | 6.8 | 8.0 | 1.4 | 0.2 |
| Norway spruce | - | 8.7 | 0.0 | 0.0 | 0.0 |
| Norway maple | - | 1.5 | 0.7 | 0.0 | 0.0 |
| Common ash | - | 0.8 | 0.0 | ||
| Hornbeam | 10.1 (10.0) | 4.0 | 10.4 | ||
| Willows | 2.9 (1.5) | 5.0 | 5.6 | ||
| Mountain ash | - | 2.2 | 2.0 | ||
| Bird cherry | - | 23.7 | 12.6 | ||
| Black cherry | - | 5.8 | 0.0 | ||
| Common hazel | 34.8 (8.8) | 13.7 | 12.8 | ||
| Alder buckthorn | - | 14.5 | 29.1 | ||
| Common dogwood | - | 1.2 | 1.1 | ||
| Euonymus | - | 1.1 | 0.0 | ||
| Common hawthorn | - | 5.0 | 0.0 | ||
| Guelder rose | - | 0.0 | 0.4 | ||
| Total | 100 (6.2) | 100.0 | 100.0 | 100.0 | 100.0 |
| Index | Variant | Zone | All | ||||||
|---|---|---|---|---|---|---|---|---|---|
| a (0–6 m) | b (22–28 m) | c (44–50 m) | |||||||
| (i) Average ± Standard Deviation 1,2 (n) Average Number of Species | |||||||||
| Layer I (trees taller than 10 m) | |||||||||
| i | n | i | n | i | n | i | n | ||
| H | B | 0.64 ± 0.31 Aa | 2.2 | 0.63 ± 0.38 Aa | 2.2 | 1.08 ± 0.38 Aa | 3.0 | 0,76 ± 0.41 A | 2.5 |
| N | 0.37 ± 0.10 Ba | 2.0 | 0.43 ± 0.24 Aa | 2.2 | 0.60 ± 0.71 Aa | 2.5 | 0.47 ± 0.41 B | 2.3 | |
| R | B | 0.47 ± 0.19 Aa | 2.2 | 0.44 ± 0.17 Aa | 2.2 | 0.73 ± 0.31 Aa | 3.0 | 0.53 ± 0.25 A | 2.5 |
| N | 0.41 ± 0.04 Aa | 2.0 | 0.43 ± 0.09 Aa | 2.2 | 0.47 ± 0.32 Aa | 2.5 | 0.44 ± 0.18 A | 2.3 | |
| C | B | 0.59 ± 0.15 Aa | 2.2 | 0.58 ± 0.24 Aa | 2.2 | 0.39 ± 0.13 Aa | 3.0 | 0.53 ± 0.22 B | 2.5 |
| N | 0.79 ± 0.07 Ba | 2.0 | 0.75 ± 0.16 Aa | 2.2 | 0.67 ± 0.24 Aa | 2.5 | 0.74 ± 0.23 A | 2.3 | |
| Layer II (trees shorter than 10 m and shrubs) | |||||||||
| H | B | 1.10 ± 0.28 Aa | 4.4 | 0.74 ± 0.50 Aa | 3.6 | 0.72 ± 0.41 Aa | 3.8 | 0.85 ± 0.42 A | 3.9 |
| N | 1.00 ± 0.20 Aa | 3.5 | 0.54 ± 0.45 Aa | 3.5 | 1.11 ± 0.17 Aa | 4.2 | 0.87 ± 0.39 A | 3.8 | |
| R | B | 1.09 ± 0.49 Aa | 4.4 | 0.78 ± 0.26 Aa | 3.6 | 0.81 ± 0.49 Aa | 3.8 | 0.89 ± 0.42 A | 3.9 |
| N | 0.97 ± 0.38 Aa | 3.5 | 0.74 ± 0.51 Aa | 3.5 | 0.87 ± 0.10 Aa | 4.2 | 0.85 ± 0.34 A | 3.8 | |
| C | B | 0.42 ± 0.14 Aa | 4.4 | 0.58 ± 0.29 Aa | 3.6 | 0.61 ± 0.19 Aa | 3.8 | 0.54 ± 0.22 A | 3.9 |
| N | 0.41 ± 0.06 Ab | 3.5 | 0.71 ± 0.25 Aa | 3.5 | 0.38 ± 0.07 Ab | 4.2 | 0.51 ± 0.22 A | 3.8 | |
| Layer III (forest floor vegetation) | |||||||||
| H | B | 2.16 ± 0.42 Aa | 24.4 | 1.67 ± 0.34 Aa | 14.0 | 2.02 ± 0.31 Aa | 12.4 | 1.95 ± 0.40 A | 16.9 |
| N | 1.95 ± 0.97 Aa | 18.5 | 1.84 ± 0.69 Aa | 16.0 | 2.02 ± 0.64 Aa | 13.0 | 1.85 ± 0.71 A | 16.2 | |
| R | B | 4.02 ± 0.50 Aa | 24.4 | 2.61 ± 0.49 Ab | 14.0 | 2.78 ± 0.71 Ab | 12.4 | 3.14 ± 0.69 A | 16.9 |
| N | 3.28 ± 0.67 Ba | 18.5 | 2.84 ± 1.02 Aa | 16.0 | 2.43 ± 0.64 Aa | 13.0 | 2.85 ± 0.80 A | 16.4 | |
| C | B | 0.21 ± 0.15 Aa | 24.4 | 0.30 ± 0.13 Aa | 14.0 | 0.19 ± 0.06 Aa | 12.4 | 0.23 ± 0.12 A | 16.9 |
| N | 0.29 ± 0.31 Aa | 18.5 | 0.29 ± 0.23 Aa | 16.0 | 0.30 ± 0.23 Aa | 13.0 | 0.29 ± 0.23 A | 16.2 | |
| Index | Transect Location | |||||
|---|---|---|---|---|---|---|
| Upstream | Dam | Downstream | ||||
| (i) Average ± Standard Deviation 1 (n) Average Number of Species | ||||||
| Layer I (trees taller than 10 m) | ||||||
| i | n | i | n | i | n | |
| H | 0.67 ± 0.34 a | 2.5 | 0.57 ± 0.55 ab | 2.1 | 0.39 ± 0.26 b | 1.7 |
| R | 0.57 ± 0.35 a | 2.5 | 0.48 ± 0.26 ab | 2.1 | 0.36 ± 0.11 b | 1.7 |
| C | 0.65 ± 0.33 a | 2.5 | 0.66 ± 0.31 a | 2.1 | 0.75 ± 0.17 a | 1.7 |
| Layer II (trees shorter than 10 m and shrubs) | ||||||
| H | 0.71 ± 0.59 a | 3.5 | 0.85 ± 0.58 a | 4.0 | 0.90 ± 0.34 a | 3.8 |
| R | 0.73 ± 0.49 a | 3.5 | 0.87 ± 0.44 a | 4.0 | 0.84 ± 0.37 a | 3.8 |
| C | 0.61 ± 0.31 a | 3.5 | 0.54 ± 0.29 a | 4.0 | 0.50 ± 0.19 a | 3.8 |
| Layer III (forest floor vegetation) | ||||||
| H | 2.27 ± 0.27 a | 22.0 | 1.69 ± 0.41 b | 16.8 | 1.98 ± 0.28 ab | 18.4 |
| R | 3.82 ± 0.51 a | 22.0 | 2.95 ± 0.40 b | 16.8 | 3.34 ± 0.60 ab | 18.4 |
| C | 0.16 ± 0.03 b | 22.0 | 0.35 ± 0.14 a | 16.8 | 0.23 ± 0.16 ab | 18.4 |
| Variant/Location | Sum of Occupied Area (m2) | Area Share (%) |
|---|---|---|
| Presence of beaver | ||
| With beaver | 98.5 | 5.1 |
| Without beaver | 44.6 | 3.2 |
| Transect location | ||
| Upstream | 24.5 | 3.8 |
| Dam | 68.4 | 10.6 |
| Downstream | 5.6 | 0.9 |
| Distance from river | ||
| a (0–6 m) | 62.6 | 1.7 |
| b (22–28 m) | 22.9 | 0.6 |
| c (44–50 m) | 57.6 | 1.6 |
| Total | 143.1 | 4.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piętka, S.; Misiukiewicz, W. Impact of European Beaver (Castor fiber L.) on Vegetation Diversity in Protected Area River Valleys. Conservation 2022, 2, 613-626. https://doi.org/10.3390/conservation2040040
Piętka S, Misiukiewicz W. Impact of European Beaver (Castor fiber L.) on Vegetation Diversity in Protected Area River Valleys. Conservation. 2022; 2(4):613-626. https://doi.org/10.3390/conservation2040040
Chicago/Turabian StylePiętka, Sławomir, and Wojciech Misiukiewicz. 2022. "Impact of European Beaver (Castor fiber L.) on Vegetation Diversity in Protected Area River Valleys" Conservation 2, no. 4: 613-626. https://doi.org/10.3390/conservation2040040
APA StylePiętka, S., & Misiukiewicz, W. (2022). Impact of European Beaver (Castor fiber L.) on Vegetation Diversity in Protected Area River Valleys. Conservation, 2(4), 613-626. https://doi.org/10.3390/conservation2040040