

Assaying and Classifying T Cell Function by Cell Morphology


Abstract
:1. Introduction
2. Materials and Methods
3. Results
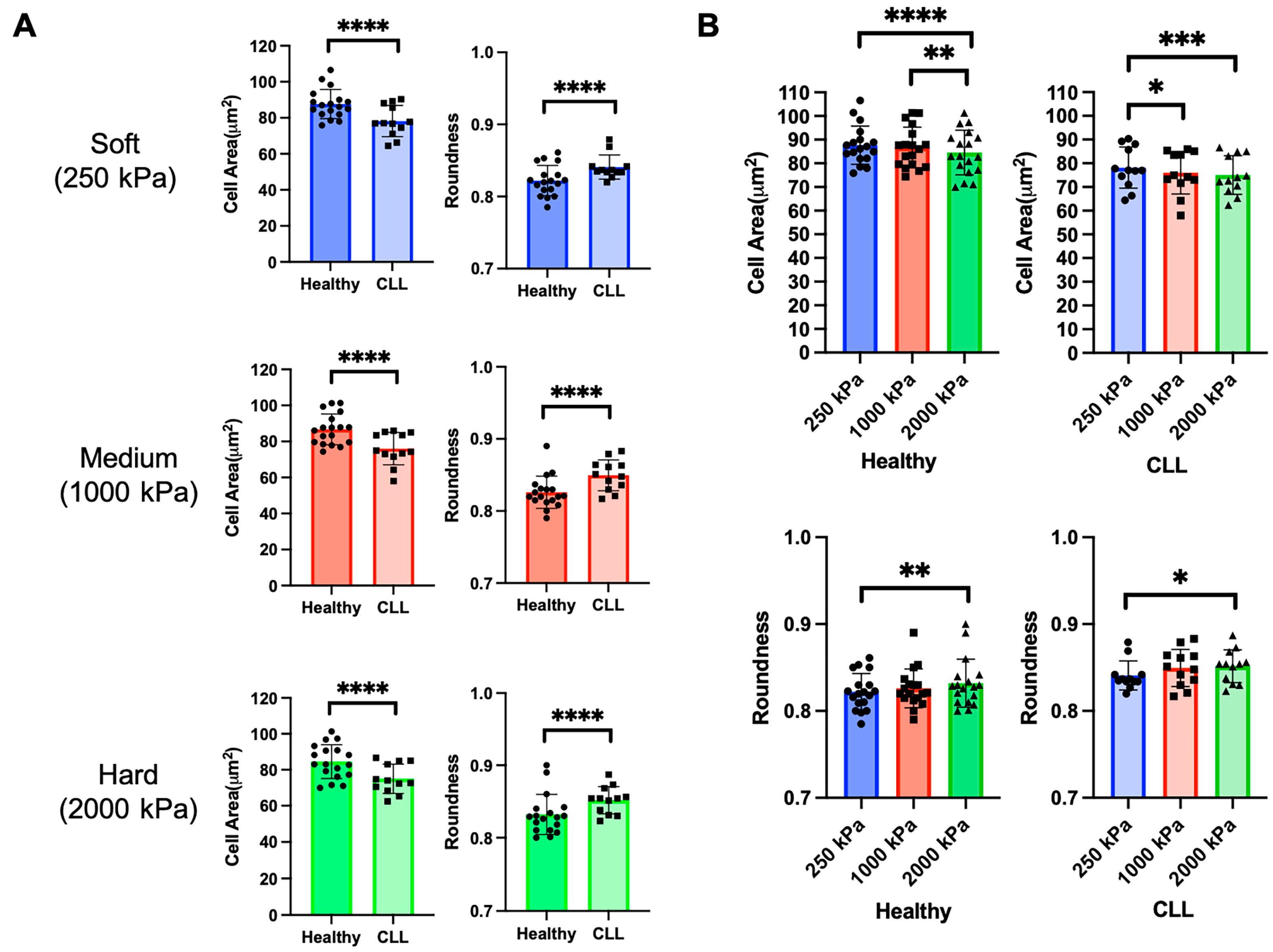
3.1. Disease State Affects T Cell Morphology
3.2. T Cells from Both Healthy Donors and CLL Patients Respond to Substrate Stiffness
3.3. PCA Reveals the Variance between CLL and Healthy T Cells and Identifies Important Morphological Features Contributing to the Variance
3.4. Machine Learning Classifies CLL Patients Based on Morphological Features
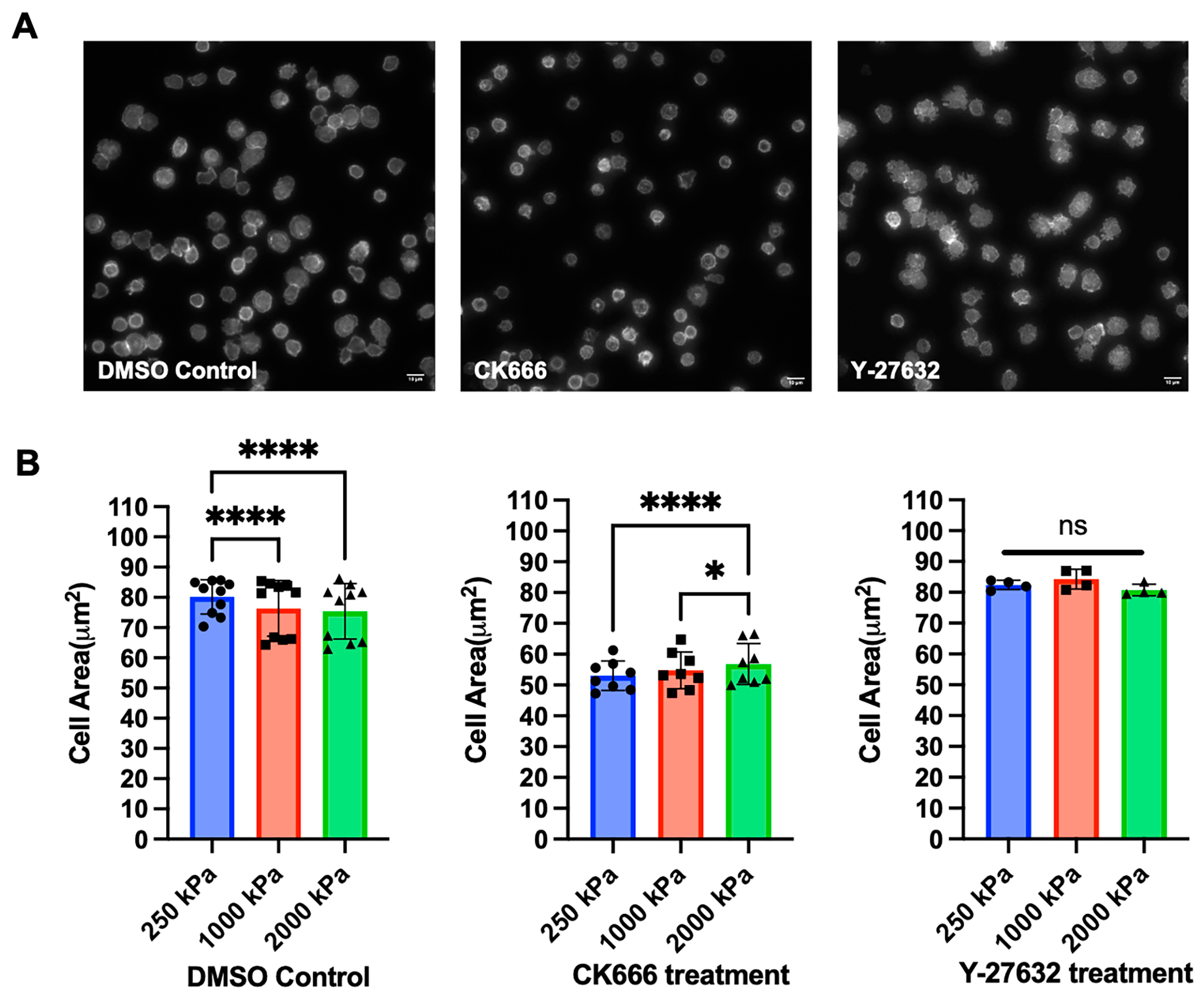
3.5. Effect of Cytoskeletal Protein Inhibitors on T Cell Response to Substrate Stiffness
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fesnak, A.D.; Levine, B.L.; June, C.H. Engineered T Cells: The Promise and Challenges of Cancer Immunotherapy. Nat. Rev. Cancer 2016, 16, 566–581. [Google Scholar] [CrossRef] [PubMed]
- Restifo, N.P.; Dudley, M.E.; Rosenberg, S.A. Adoptive immunotherapy for cancer: Harnessing the T cell response. Nat. Rev. Immunol. 2012, 12, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Kalos, M.; Levine, B.L.; Porter, D.L.; Katz, S.; Grupp, S.A.; Bagg, A.; June, C.H. T cells with chimeric antigen receptors have potent antitumor effects and can establish memory in patients with advanced leukemia. Sci. Transl. Med. 2011, 3, 95ra73. [Google Scholar] [CrossRef] [PubMed]
- Porter, D.L.; Levine, B.L.; Kalos, M.; Bagg, A.; June, C.H. Chimeric antigen receptor-modified T cells in chronic lymphoid leukemia. N. Engl. J. Med. 2011, 365, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, A.G.; Johnson, A.J.; Lee, A.M.; Gorgün, G.; Le Dieu, R.; Blum, W.; Byrd, J.C.; Gribben, J.G. Chronic lymphocytic leukemia T cells show impaired immunological synapse formation that can be reversed with an immunomodulating drug. J. Clin. Investig. 2008, 118, 2427–2437. [Google Scholar] [CrossRef] [PubMed]
- Riches, J.C.; Davies, J.K.; McClanahan, F.; Fatah, R.; Iqbal, S.; Agrawal, S.; Ramsay, A.G.; Gribben, J.G. T cells from CLL patients exhibit features of T-cell exhaustion but retain capacity for cytokine production. Blood 2013, 121, 1612–1621. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.; Gentilcore, G.; Heimersson, K.; Mozaffari, F.; Näsman-Glaser, B.; Young, E.; Rosenquist, R.; Hansson, L.; Österborg, A.; Mellstedt, H. T cells in chronic lymphocytic leukemia display dysregulated expression of immune checkpoints and activation markers. Haematologica 2017, 102, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Scrivener, S.; Kaminski, E.R.; Demaine, A.; Prentice, A.G. Analysis of the expression of critical activation/interaction markers on peripheral blood T cells in B-cell chronic lymphocytic leukemia: Evidence of immune dysregulation. Br. J. Haematol. 2001, 112, 959–964. [Google Scholar] [CrossRef]
- Burger, J.A.; Gribben, J.G. The microenvironment in chronic lymphocytic leukemia (CLL) and other B cell malignancies: Insight into disease biology and new targeted therapies. Semin. Cancer Biol. 2014, 24, 71–81. [Google Scholar] [CrossRef]
- Gassner, F.J.; Zaborsky, N.; Catakovic, K.; Rebhandl, S.; Huemer, M.; Egle, A.; Hartmann, T.N.; Greil, R.; Geisberger, R. Chronic lymphocytic leukaemia induces an exhausted T cell phenotype in the TCL1 transgenic mouse model. Br. J. Haematol. 2015, 170, 515–522. [Google Scholar] [CrossRef]
- Wu, J.; Xu, X.; Lee, E.-J.; Shull, A.Y.; Pei, L.; Awan, F.; Wang, X.; Choi, J.-H.; Deng, L.; Xin, H.-B.; et al. Phenotypic alteration of CD8+ T cells in chronic lymphocytic leukemia is associated with epigenetic reprogramming. Oncotarget 2016, 7, 40558–40570. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Shao, S.; Kim, M.; Fernandes, S.M.; Brown, J.R.; Kam, L.C. Multi-Factor Clustering Incorporating Cell Motility Predicts T Cell Expansion Potential. Front. Cell Dev. Biol. 2021, 9, 648925. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Suo, Y.; Deng, Y.; Fan, Z.; Zheng, Y.; Wei, X.; Chu, Y. Morphological change of CD4(+) T cell during contact with DC modulates T-cell activation by accumulation of F-actin in the immunology synapse. BMC Immunol. 2015, 16, 49. [Google Scholar] [CrossRef] [PubMed]
- Grakoui, A.; Bromley, S.K.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The immunological synapse: A molecular machine controlling T cell activation. Science 1999, 285, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Wahl, A.; Dinet, C.; Dillard, P.; Nassereddine, A.; Puech, P.-H.; Limozin, L.; Sengupta, K. Biphasic mechanosensitivity of T cell receptor-mediated spreading of lymphocytes. Proc. Natl. Acad. Sci. USA 2019, 116, 5908–5913. [Google Scholar] [CrossRef] [PubMed]
- Cretel, E.; Touchard, D.; Bongrand, P.; Pierres, A. A new method for rapid detection of T lymphocyte decision to proliferate after encountering activating surfaces. J. Immunol. Methods 2011, 364, 33–39. [Google Scholar] [CrossRef]
- O’Connor, R.S.; Hao, X.; Shen, K.; Bashour, K.; Akimova, T.; Hancock, W.W.; Kam, L.C.; Milone, M.C. Substrate rigidity regulates human T cell activation and proliferation. J. Immunol. 2012, 189, 1330–1339. [Google Scholar] [CrossRef]
- Judokusumo, E.; Tabdanov, E.; Kumari, S.; Dustin, M.L.; Kam, L.C. Mechanosensing in T Lymphocyte Activation. Biophys. J. 2012, 102, L5–L7. [Google Scholar] [CrossRef]
- Saitakis, M.; Dogniaux, S.; Goudot, C.; Bufi, N.; Asnacios, S.; Maurin, M.; Randriamampita, C.; Asnacios, A.; Hivroz, C. Different TCR-induced T lymphocyte responses are potentiated by stiffness with variable sensitivity. eLife 2017, 6, e23190. [Google Scholar] [CrossRef]
- Sachar, C.; Kam, L.C. Probing T Cell 3D Mechanosensing With Magnetically-Actuated Structures. Front. Immunol. 2021, 12, 704693. [Google Scholar] [CrossRef]
- Palchesko, R.N.; Zhang, L.; Sun, Y.; Feinberg, A.W. Development of polydimethylsiloxane substrates with tunable elastic modulus to study cell mechanobiology in muscle and nerve. PLoS ONE 2012, 7, e51499. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.J.; Shi, L.; Kam, L.C. Biphasic response of T cell activation to substrate stiffness. Biomaterials 2021, 273, 120797. [Google Scholar] [CrossRef] [PubMed]
- Citing. ImageJ Wiki. Available online: https://imagej.github.io/contribute/citing (accessed on 28 January 2024).
- Chaudhuri, P.K.; Wang, M.S.; Black, C.T.; Huse, M.; Kam, L.C. Modulating T Cell Activation Using Depth Sensing Topographic Cues. Adv. Biosyst. 2020, 4, e2000143. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Hotelling, H. Analysis of a complex of statistical variables into principal components. J. Educ. Psychol. 1933, 24, 417–441. [Google Scholar] [CrossRef]
- Gonnord, P.; Costa, M.; Abreu, A.; Peres, M.; Ysebaert, L.; Gadat, S.; Valitutti, S. Multiparametric analysis of CD8+ T cell compartment phenotype in chronic lymphocytic leukemia reveals a signature associated with progression toward therapy. Oncoimmunology 2019, 8, e1570774. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, J.R. Induction of decision trees. Mach. Learn. 1986, 1, 81–106. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Breiman, L. Classification and Regression Trees; Routledge: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Dang, A.; De Leo, S.; Bogdanowicz, D.R.; Yuan, D.J.; Fernandes, S.M.; Brown, J.R.; Lu, H.H.; Kam, L.C. Enhanced activation and expansion of T cells using mechanically soft elastomer fibers. Adv. Biosyst. 2018, 2, 1700167. [Google Scholar] [CrossRef]
- Anandakumaran, P.N.; Ayers, A.G.; Muranski, P.; Creusot, R.J.; Sia, S.K. Rapid video-based deep learning of cognate versus non-cognate T cell-dendritic cell interactions. Sci. Rep. 2022, 12, 559. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Single-Feature (Area) Decision Tree | Multi-Feature Decision Tree | Random Forest | |
|---|---|---|---|
| Accuracy | 0.677 | 0.651 | 0.753 |
| AUC | 0.683 | 0.650 | 0.812 |
| Sensitivity | 0.673 | 0.680 | 0.750 |
| Specificity | 0.693 | 0.616 | 0.753 |
| MCC | 0.372 | 0.649 | 0.596 |
| Single-Feature (Area) Decision Tree | Multi-Feature Decision Tree | Random Forest | |
|---|---|---|---|
| Accuracy | 0.729 | 0.693 | 0.765 |
| AUC | 0.735 | 0.694 | 0.846 |
| Sensitivity | 0.746 | 0.708 | 0.725 |
| Specificity | 0.725 | 0.680 | 0.819 |
| MCC | 0.409 | 0.549 | 0.580 |
| Single-Feature (Area) Decision Tree | Multi-Feature Decision Tree | Random Forest | |
|---|---|---|---|
| Accuracy | 0.749 | 0.690 | 0.756 |
| AUC | 0.760 | 0.698 | 0.836 |
| Sensitivity | 0.749 | 0.670 | 0.713 |
| Specificity | 0.770 | 0.725 | 0.812 |
| MCC | 0.490 | 0.510 | 0.627 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Fernandes, S.M.; Brown, J.R.; Kam, L.C. Assaying and Classifying T Cell Function by Cell Morphology. BioMedInformatics 2024, 4, 1144-1154. https://doi.org/10.3390/biomedinformatics4020063
Wang X, Fernandes SM, Brown JR, Kam LC. Assaying and Classifying T Cell Function by Cell Morphology. BioMedInformatics. 2024; 4(2):1144-1154. https://doi.org/10.3390/biomedinformatics4020063
Chicago/Turabian StyleWang, Xin, Stacey M. Fernandes, Jennifer R. Brown, and Lance C. Kam. 2024. "Assaying and Classifying T Cell Function by Cell Morphology" BioMedInformatics 4, no. 2: 1144-1154. https://doi.org/10.3390/biomedinformatics4020063
APA StyleWang, X., Fernandes, S. M., Brown, J. R., & Kam, L. C. (2024). Assaying and Classifying T Cell Function by Cell Morphology. BioMedInformatics, 4(2), 1144-1154. https://doi.org/10.3390/biomedinformatics4020063