
The Plagiochilins from Plagiochila Liverworts: Binding to α-Tubulin and Drug Design Perspectives


Abstract
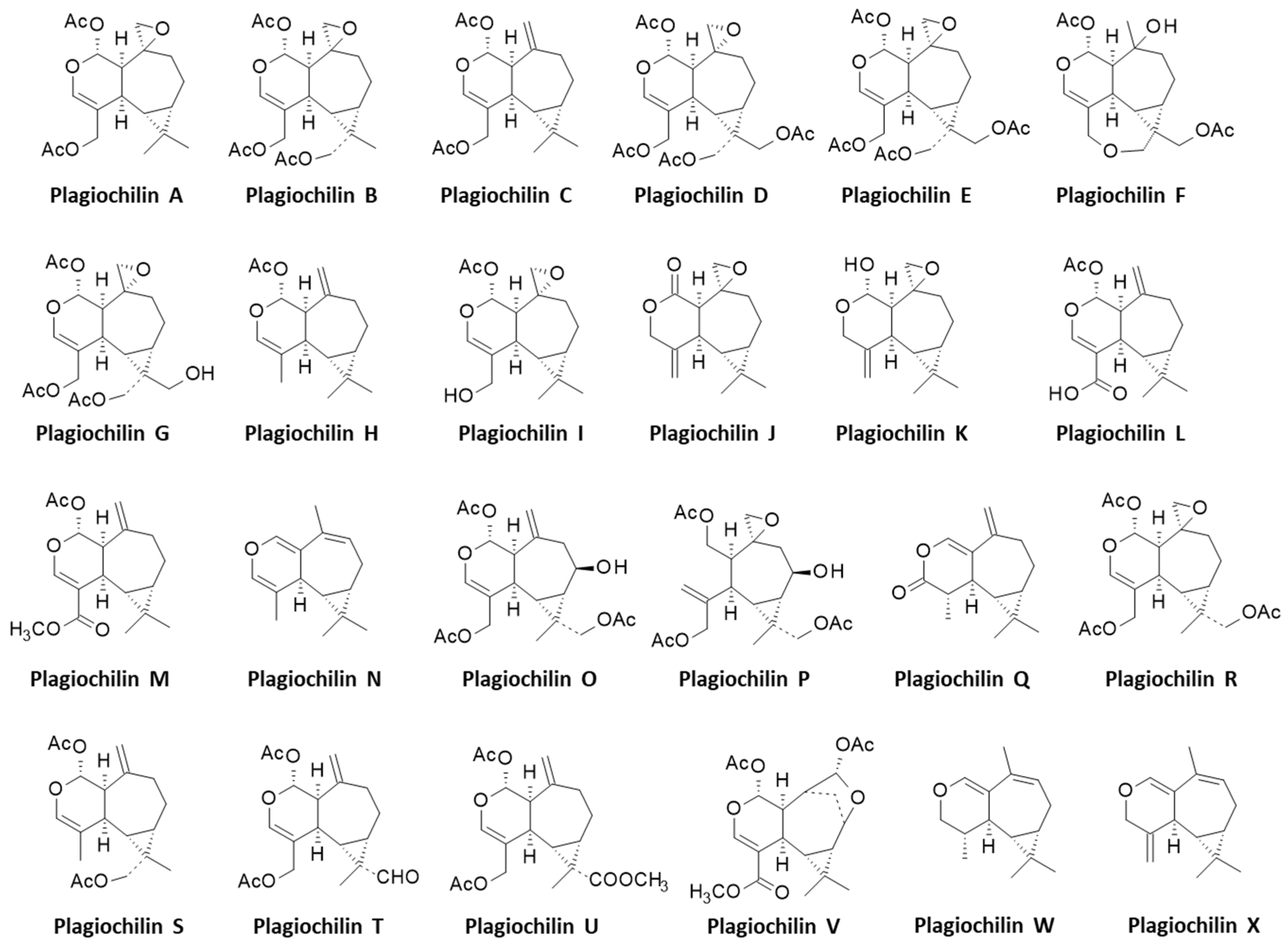
:1. Introduction
2. Materials and Methods
2.1. Molecular Structure and Software
2.2. Molecular Modeling Procedure
3. Results
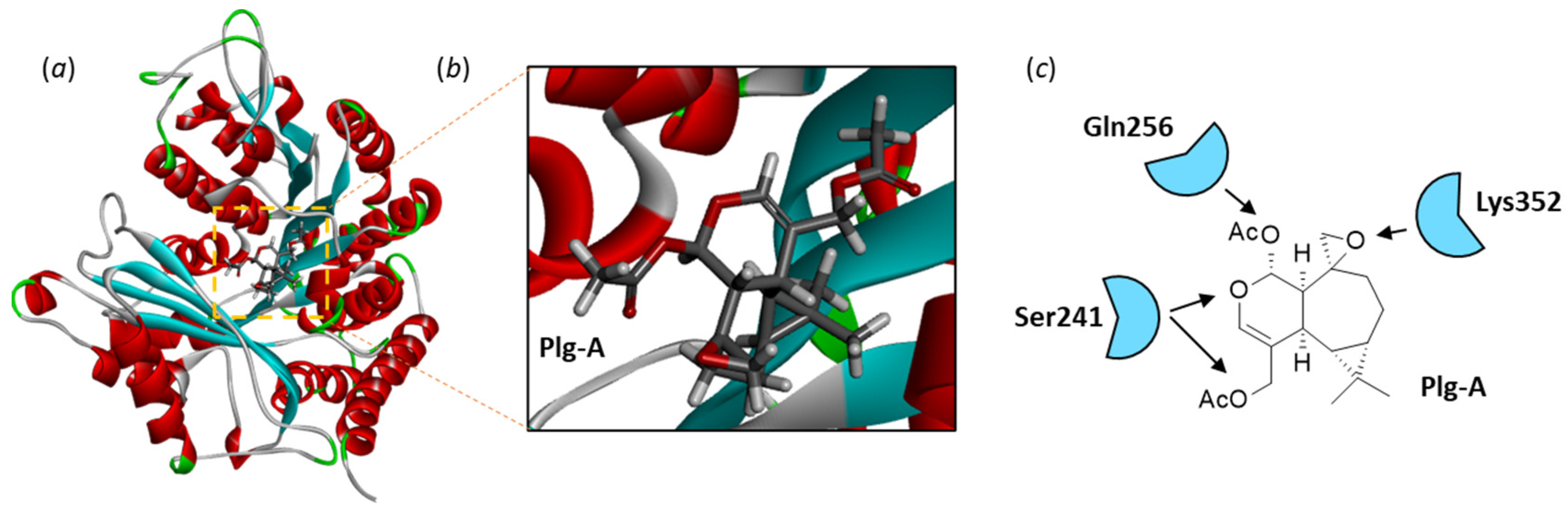
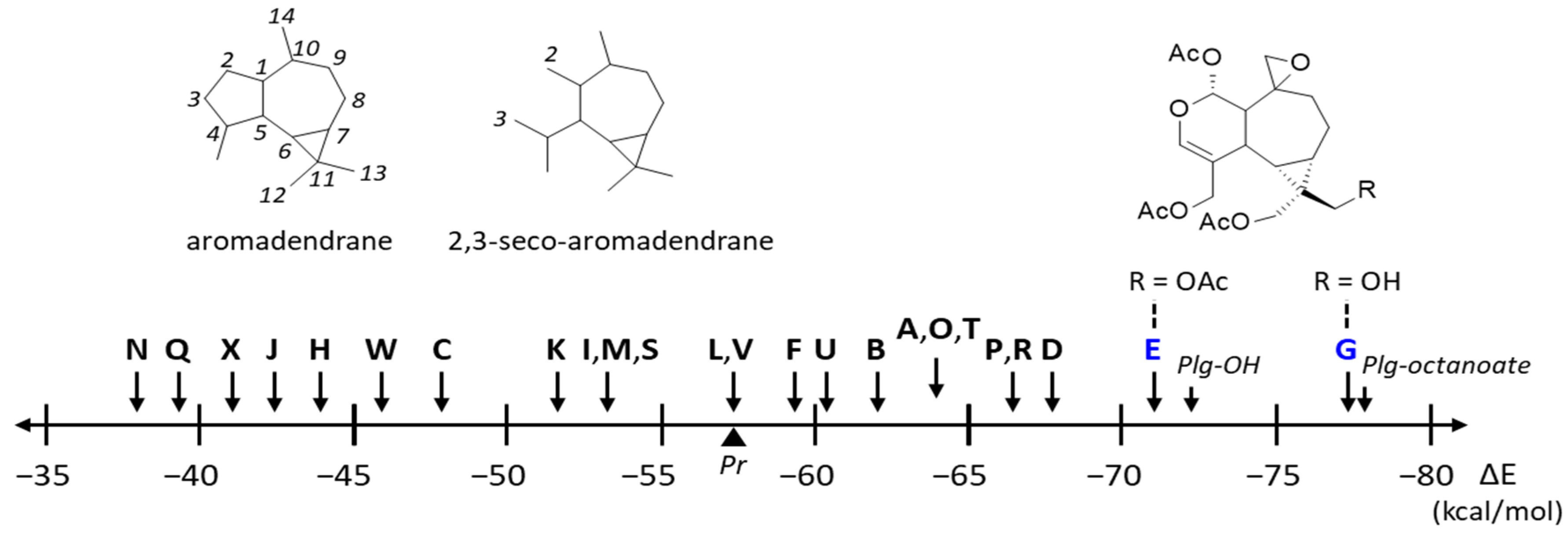
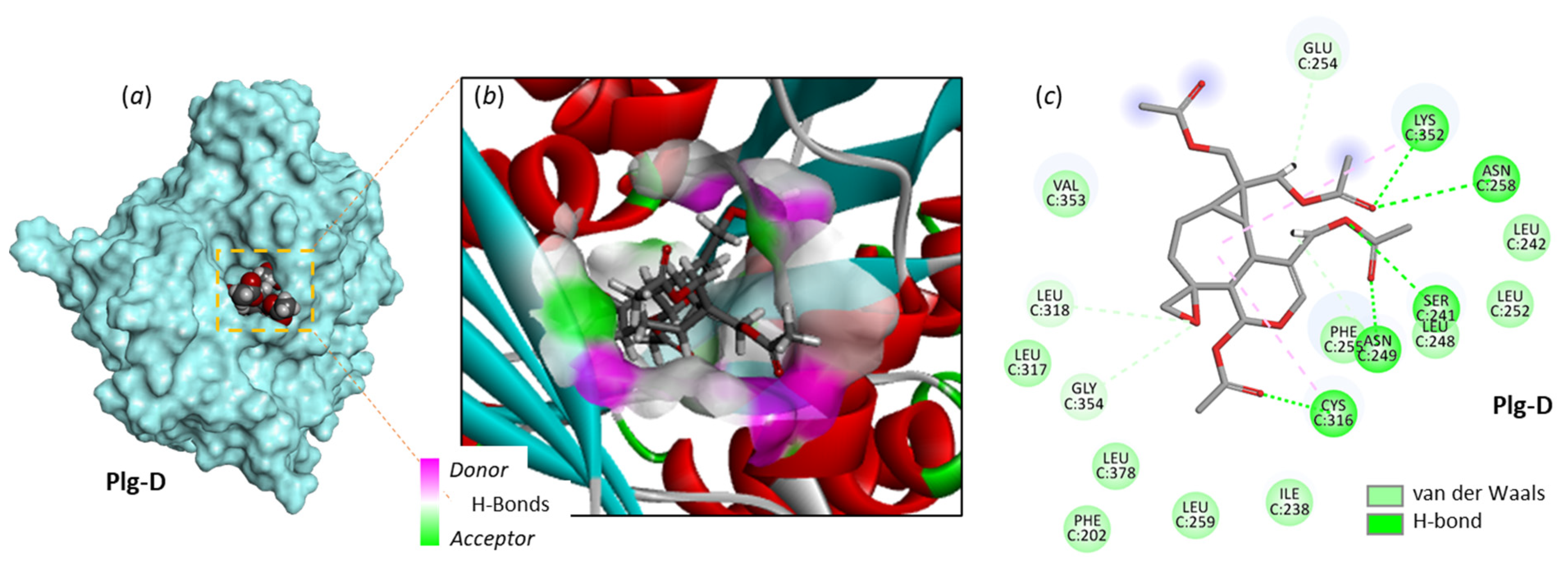
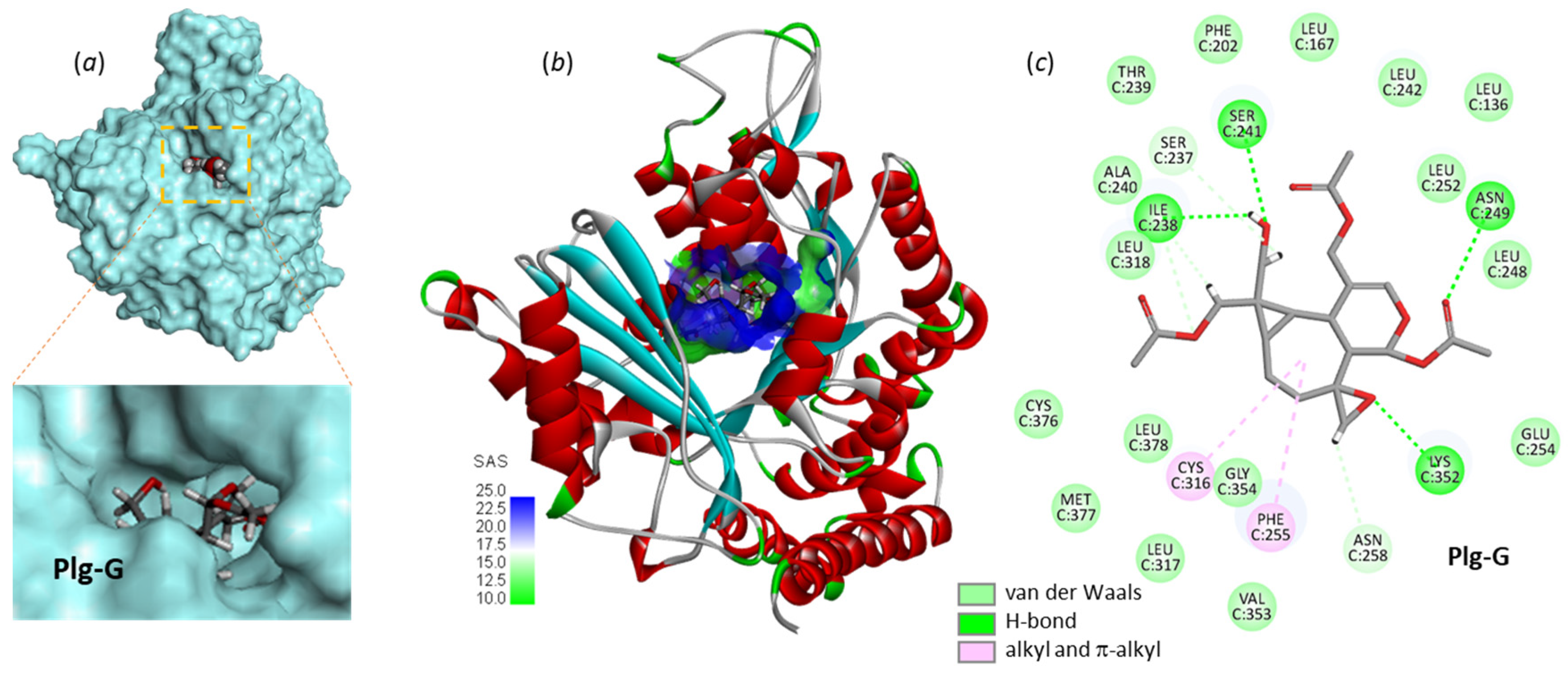
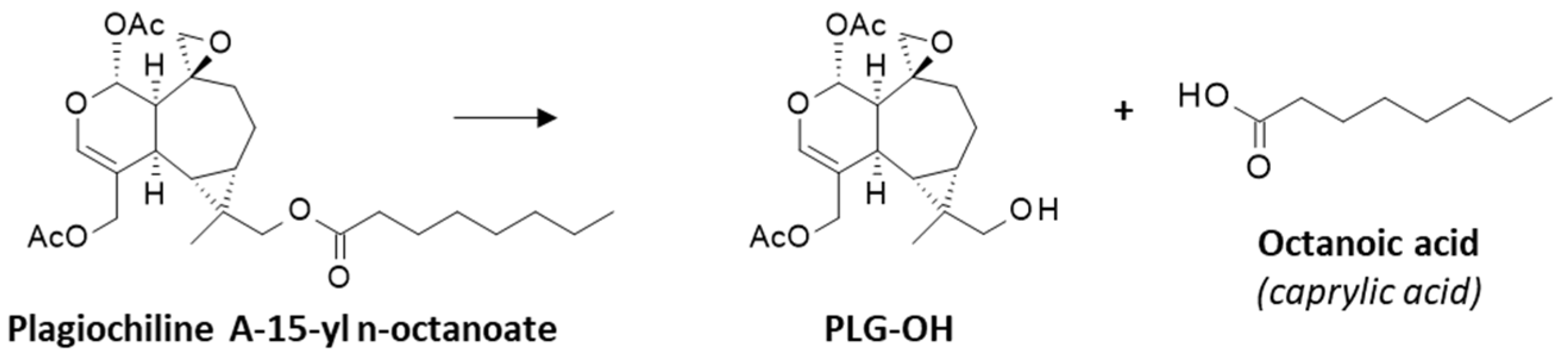
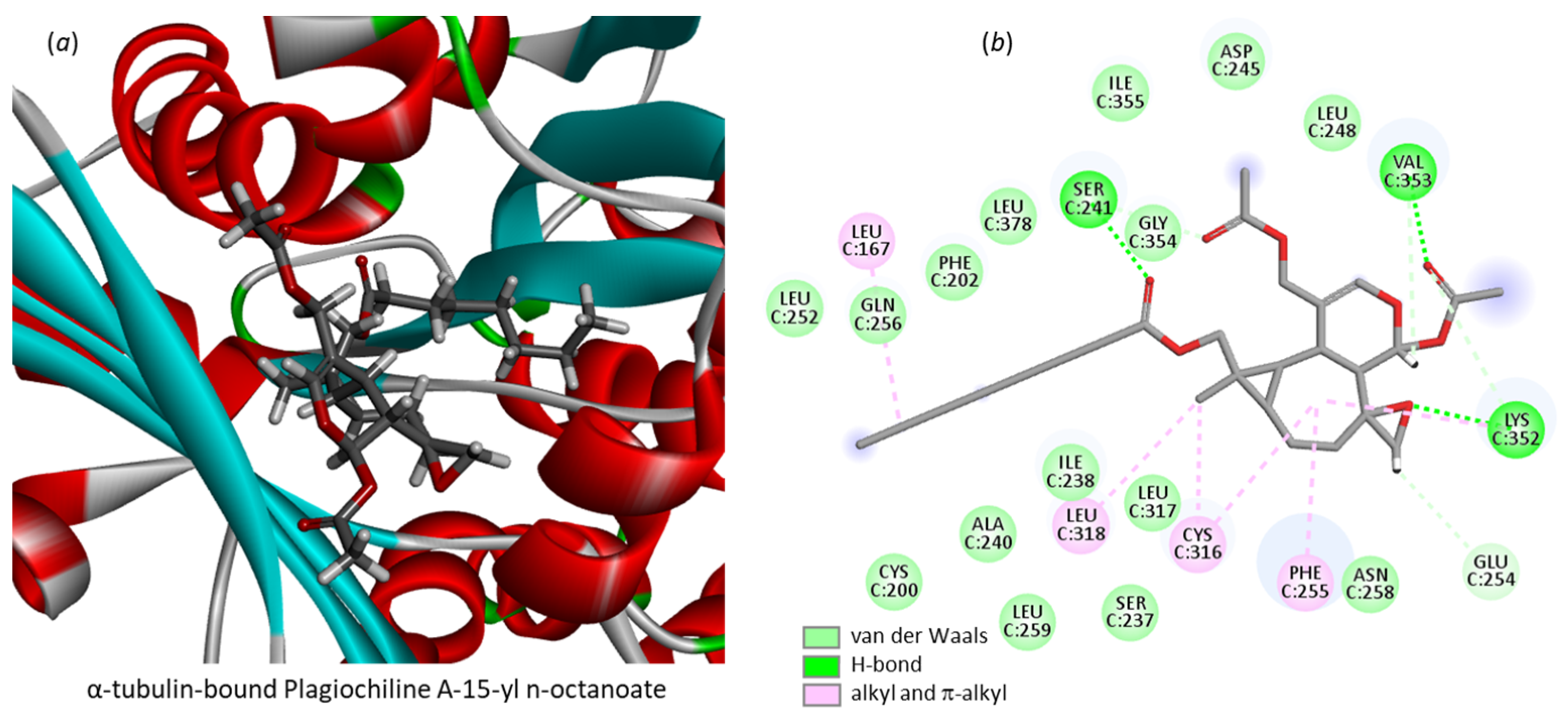
3.1. Comparative Docking of Plagiochilins to α-Tubulin
3.2. Drug Design Implications
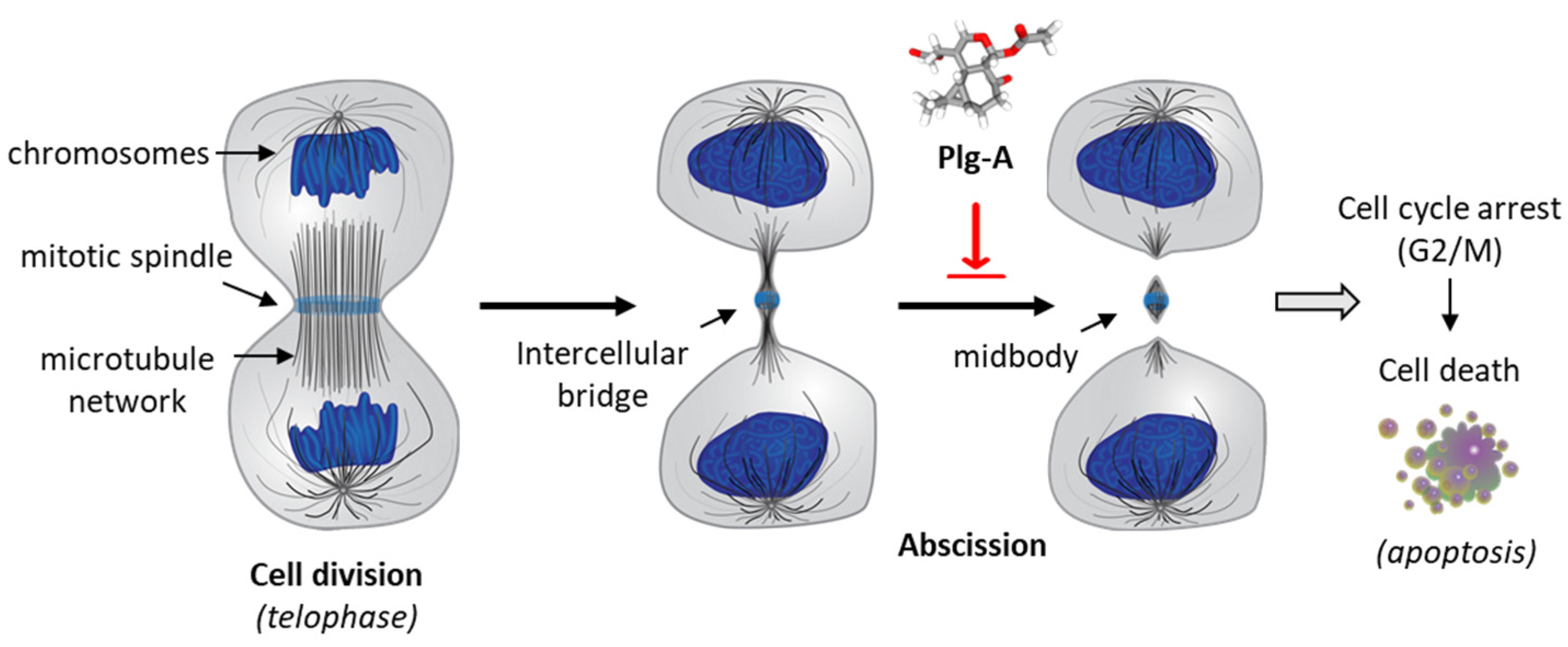
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frangedakis, E.; Shimamura, M.; Villarreal, J.C.; Li, F.W.; Tomaselli, M.; Waller, M.; Sakakibara, K.; Renzaglia, K.S.; Szövényi, P. The hornworts: Morphology, evolution and development. New Phytol. 2021, 229, 735–754. [Google Scholar] [CrossRef] [PubMed]
- Drobnik, J.; Stebel, A. Four Centuries of Medicinal Mosses and Liverworts in European Ethnopharmacy and Scientific Pharmacy: A Review. Plants 2021, 10, 1296. [Google Scholar] [CrossRef] [PubMed]
- Commisso, M.; Guarino, F.; Marchi, L.; Muto, A.; Piro, A.; Degola, F. Bryo-Activities: A Review on How Bryophytes Are Contributing to the Arsenal of Natural Bioactive Compounds against Fungi. Plants 2021, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, Y.; Nagashima, F.; Ludwiczuk, A. Distribution of Bibenzyls, Prenyl Bibenzyls, Bis-bibenzyls, and Terpenoids in the Liverwort Genus Radula. J. Nat. Prod. 2020, 83, 756–769. [Google Scholar] [CrossRef]
- Ludwiczuk, A.; Asakawa, Y. Bryophytes as a source of bioactive volatile terpenoids—A review. Food Chem. Toxicol. 2019, 132, 110649. [Google Scholar] [CrossRef]
- Asakawa, Y.; Toyota, M.; Takemoto, T. Plagiochilide et plagiochiline a, secoaromadendrane-type sesquiterpenes de la mousse, plagiochila yokogurensis (plagiochilaceae). Tetrahedron Lett. 1978, 19, 1553–1556. [Google Scholar] [CrossRef]
- Asakawa, Y.; Toyota, M.; Takemoto, T. La plagiochiline a et la plagiochiline b, les sesquiterpenes du type secoaromadendrane de la mousse, Plagiochila hattoriana. Phytochemistry 1978, 17, 1794. [Google Scholar] [CrossRef]
- Asakawa, Y.; Toyota, M.; Takemoto, T.; Suire, C. Plagiochilines C, D, E and F, four novel secoaromadendrane-type sesquiterpene hemiacetals from Plagiochila asplenioides and Plagiochila semidecurrens. Phytochemistry 1979, 18, 1355–1357. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Toyota, M.; Asakawa, Y. Ent-kaurene diterpene from the liverwort Plagiochila pulcherrima. Phytochemistry 1988, 27, 1425–1427. [Google Scholar] [CrossRef]
- Aponte, J.C.; Yang, H.; Vaisberg, A.J.; Castillo, D.; Málaga, E.; Verástegui, M.; Casson, L.K.; Stivers, N.; Bates, P.J.; Rojas, R.; et al. Cytotoxic and anti-infective sesquiterpenes present in Plagiochila disticha (Plagiochilaceae) and Ambrosia peruviana (Asteraceae). Planta Med. 2010, 76, 705–707. [Google Scholar] [CrossRef]
- Asakawa, Y.; Inoue, H.; Toyota, M.; Takemoto, T. Sesquiterpenoids of fourteen Plagiochila species. Phytochemistry 1980, 19, 623–2626. [Google Scholar] [CrossRef]
- Bailly, C. Discovery and anticancer activity of the plagiochilins from the liverwort genus Plagiochila. Life 2023, 13, 758. [Google Scholar] [CrossRef]
- Asakawa, Y. Chemosystematics of the hepaticae. Phytochemistry 2004, 65, 623–669. [Google Scholar] [CrossRef]
- Stivers, N.S.; Islam, A.; Reyes-Reyes, E.M.; Casson, L.K.; Aponte, J.C.; Vaisberg, A.J.; Hammond, G.B.; Bates, P.J. Plagiochiline A Inhibits Cytokinetic Abscission and Induces Cell Death. Molecules 2018, 23, 1418. [Google Scholar] [CrossRef]
- McNeely, K.C.; Dwyer, N.D. Cytokinetic Abscission Regulation in Neural Stem Cells and Tissue Development. Curr. Stem Cell Rep. 2021, 7, 161–173. [Google Scholar] [CrossRef]
- Peterman, E.; Prekeris, R. The postmitotic midbody: Regulating polarity, stemness, and proliferation. J. Cell. Biol. 2019, 218, 3903–3911. [Google Scholar] [CrossRef]
- Huang, D.S.; Wong, H.L.; Georg, G.I. Synthesis and Cytotoxicity Evaluation of C4- and C5-Modified Analogues of the α,β-Unsaturated Lactone of Pironetin. ChemMedChem 2017, 12, 520–528. [Google Scholar] [CrossRef]
- Coulup, S.K.; Georg, G.I. Revisiting microtubule targeting agents: α-Tubulin and the pironetin binding site as unexplored targets for cancer therapeutics. Bioorg. Med. Chem. Lett. 2019, 29, 1865–1873. [Google Scholar] [CrossRef]
- Toyota, M.; Tanimura, K.; Asakawa, Y. Cytotoxic 2,3-secoaromadendrane-type sesquiterpenoids from the liverwort Plagiochila ovalifolia. Planta Med. 1998, 64, 462–464. [Google Scholar] [CrossRef]
- Toyota, M.; Nakamura, I.; Huneck, S.; Asakawa, Y. Sesquiterpene esters from the liverwort Plagiochila porelloides. Phytochemistry 1994, 37, 1091–1093. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Wang, T.; Jiang, J.; Botting, C.H.; Liu, H.; Chen, Q.; Yang, J.; Naismith, J.H.; Zhu, X.; et al. Pironetin reacts covalently with cysteine-316 of α-tubulin to destabilize microtubule. Nat. Commun. 2016, 7, 12103. [Google Scholar] [CrossRef] [PubMed]
- Dundas, J.; Ouyang, Z.; Tseng, J.; Binkowski, A.; Turpaz, Y.; Liang, J. CASTp: Computed atlas of surface topography of proteins with structural and topographical mapping of functionally annotated residues. Nucleic Acids Res. 2006, 34, W116–W118. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed atlas of surface topography of proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Tirado-Rives, J. Monte Carlo versus Molecular Dynamics for conformational sampling. J. Phys. Chem. 1996, 100, 14508–14513. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Tirado-Rives, J. Molecular modeling of organic and biomolecular systems using BOSS and MCPRO. J. Comput. Chem. 2005, 26, 1689–1700. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Ulmschneider, J.P.; Tirado-Rives, J. Free energies of hydration from a generalized Born model and an ALL-atom force field. J. Phys. Chem. B 2004, 108, 16264–16270. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef]
- Meziane-Tani, M.; Lagant, P.; Semmoud, A.; Vergoten, G. The SPASIBA force field for chondroitin sulfate: Vibrational analysis of D-glucuronic and N-acetyl-D-galactosamine 4-sulfate sodium salts. J. Phys. Chem. A 2006, 110, 11359–11370. [Google Scholar] [CrossRef]
- Vergoten, G.; Mazur, I.; Lagant, P.; Michalski, J.C.; Zanetta, J.P. The SPASIBA force field as an essential tool for studying the structure and dynamics of saccharides. Biochimie 2003, 85, 65–73. [Google Scholar] [CrossRef]
- Lagant, P.; Nolde, D.; Stote, R.; Vergoten, G.; Karplus, M. Increasing Normal Modes Analysis Accuracy: The SPASIBA Spectroscopic Force Field Introduced into the CHARMM Program. J. Phys. Chem. A 2004, 108, 4019–4029. [Google Scholar] [CrossRef]
- Homans, S.W. A molecular mechanical force field for the conformational analysis of oligosaccharides: Comparison of theoretical and crystal structures of Man alpha 1-3Man beta 1-4GlcNAc. Biochemistry 1990, 29, 9110–9118. [Google Scholar] [CrossRef]
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; da Costa, D.P.; Crandall-Stotler, B.J.; et al. World checklist of hornworts and liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef]
- Heinrichs, J.; Hentschel, J.; Feldberg, K.; Bombosch, A.; Schneider, H. Phylogenetic biogeography and taxonomy of disjunctly distributed bryophytes. J. Syst. Evol. 2009, 47, 497–508. [Google Scholar] [CrossRef]
- Asakawa, Y.; Toyota, M.; Takemoto, T. Three ent-secoaromadendrane-type sesquiterpene hemiacetals and a bicyclogermacrene from Plagiochila ovalifolia and Plagiochila yokogurensis. Phytochemistry 1980, 19, 2141–2145. [Google Scholar] [CrossRef]
- Asakawa, Y.; Toyota, M.; Takemoto, T.; Kubo, I.; Nakanishi, K. Insect antifeedant secoaromadendrane-type sesquiterpenes from Plagiochila species. Phytochemistry 1980, 19, 2147–2154. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Phytochemical and biological studies of bryophytes. Phytochemistry 2013, 91, 52–80. [Google Scholar] [CrossRef]
- Wang, S.; Liu, S.S.; Lin, Z.M.; Li, R.J.; Wang, X.N.; Zhou, J.C.; Lou, H.X. Terpenoids from the Chinese liverwort Plagiochila pulcherrima and their cytotoxic effects. J. Asian Nat. Prod. Res. 2013, 15, 473–481. [Google Scholar] [CrossRef]
- Vergoten, G.; Bailly, C. Molecular Docking of Cryptoconcatones to α-Tubulin and Related Pironetin Analogues. Plants 2023, 12, 296. [Google Scholar] [CrossRef]
- Blay, G.; Cardona, L.; García, B.; Lahoz, L.; Pedro, J.R. Synthesis of plagiochilin N from santonin. J. Org. Chem. 2001, 66, 7700–7705. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Ćosić, M.V.; Jadranin, B.Z.; Pantović, J.P.; Giba, Z.S.; Vujičić, M.M.; Sabovljević, A.D. The Conservation Physiology of Bryophytes. Plants 2022, 11, 1282. [Google Scholar] [CrossRef]
- Pandey, S.; Alam, A. Bryo-Pharmaceuticals: An Emerging Era of Pharmaceutical Products. In Advanced Pharmacological Uses of Medicinal Plants and Natural Products; IGI Global: Hershey, PA, USA, 2020; Chapter 14; pp. 269–284. [Google Scholar] [CrossRef]
- Pannequin, A.; Quetin-Leclercq, J.; Costa, J.; Tintaru, A.; Muselli, A. First Phytochemical Profiling and In-Vitro Antiprotozoal Activity of Essential Oil and Extract of Plagiochila porelloides. Molecules 2023, 28, 616. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.N.; Jin, X.Y.; Zhou, J.C.; Zhang, J.Z.; Chang, W.Q.; Li, Y.; Chen, W.; Ren, Z.J.; Zhang, C.Y.; Yuan, S.Z.; et al. Terpenoids from the Liverwort Plagiochila fruticosa and Their Antivirulence Activity against Candida albicans. J. Nat. Prod. 2020, 83, 1766–1777. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, F.; Sekiguchi, T.; Takaoka, S.; Asakawa, Y. Terpenoids and aromatic compounds from the New Zealand liverworts Plagiochila, Schistochila, and Heteroscyphus species. Chem. Pharm. Bull. 2004, 52, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, G.; Mühlbacher, J.; Reichert, M.; Dreyer, M.; Kolz, J.; Speicher, A. Stereochemistry of isoplagiochin C, a macrocyclic bisbibenzyl from liverworts. J. Am. Chem. Soc. 2004, 126, 9283–9290. [Google Scholar] [CrossRef]
- So, M.L.; Grolle, R. Studies on Plagiochila sect. Plagiochila (Hepaticae) in East and South Asia. J. Bryol. 2000, 22, 17–28. [Google Scholar] [CrossRef]
- Matsuo, A.; Ono, K.; Hamasaki, K.; Nozaki, H. Phaeophytins from a cell suspension culture of the liverwort Plagiochila ovalifolia. Phytochemistry 1996, 42, 427–430. [Google Scholar] [CrossRef]
- Ogungbe, I.V.; Setzer, W.N. In-silico Leishmania target selectivity of antiparasitic terpenoids. Molecules 2013, 18, 7761–7847. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | ΔE (kcal/mol) | ΔG (kcal/mol) |
|---|---|---|
| PlgA-octanoate | −77.50 | −28.40 |
| Plagiochilin G | −77.00 | −26.00 |
| Plg-OH | −72.40 | −22.20 |
| Plagiochilin E | −71.00 | −17.20 |
| Plagiochilin D | −67.30 | −19.10 |
| Plagiochilin P | −66.85 | −24.00 |
| Plagiochilin R | −66.50 | −25.50 |
| Plagiochilin O | −64.40 | −23.70 |
| Plagiochilin A | −63.80 | −19.90 |
| Plagiochilin T | −63.10 | −23.30 |
| Plagiochilin B | −62.30 | −26.00 |
| Plagiochilin U | −60.40 | −20.90 |
| Plagiochilin F | −59.30 | −23.15 |
| Plagiochilin L | −57.60 | −22.70 |
| Plagiochilin V | −57.45 | −16.10 |
| Plagiochilin M | −53.70 | −21.20 |
| Plagiochilin S | −53.40 | −20.70 |
| Plagiochilin I | −53.25 | −16.80 |
| Plagiochilin K | −52.10 | −15.50 |
| Plagiochilin C | −47.80 | −18.55 |
| Plagiochilin W | −46.60 | −19.90 |
| Plagiochilin H | −44.00 | −21.30 |
| Plagiochilin J | −42.40 | −18.40 |
| Plagiochilin X | −41.30 | −20.00 |
| Plagiochilin Q | −39.30 | −17.30 |
| Plagiochilin N | −37.90 | −15.10 |
| Pironetin | −57.32 | −24.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergoten, G.; Bailly, C. The Plagiochilins from Plagiochila Liverworts: Binding to α-Tubulin and Drug Design Perspectives. AppliedChem 2023, 3, 217-228. https://doi.org/10.3390/appliedchem3020014
Vergoten G, Bailly C. The Plagiochilins from Plagiochila Liverworts: Binding to α-Tubulin and Drug Design Perspectives. AppliedChem. 2023; 3(2):217-228. https://doi.org/10.3390/appliedchem3020014
Chicago/Turabian StyleVergoten, Gérard, and Christian Bailly. 2023. "The Plagiochilins from Plagiochila Liverworts: Binding to α-Tubulin and Drug Design Perspectives" AppliedChem 3, no. 2: 217-228. https://doi.org/10.3390/appliedchem3020014
APA StyleVergoten, G., & Bailly, C. (2023). The Plagiochilins from Plagiochila Liverworts: Binding to α-Tubulin and Drug Design Perspectives. AppliedChem, 3(2), 217-228. https://doi.org/10.3390/appliedchem3020014