Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Food Consumption Experiment
2.2. Data Analyses
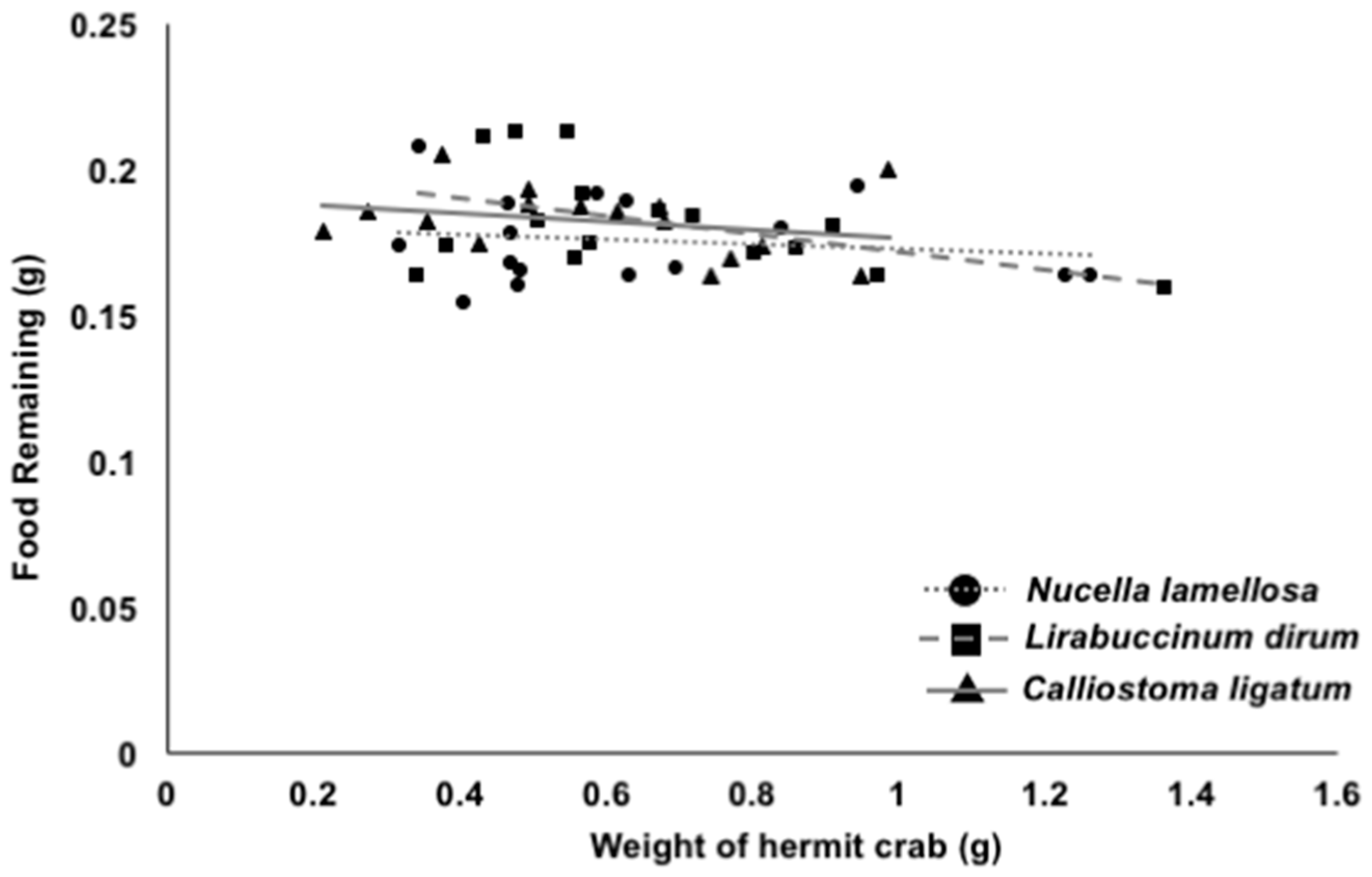
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hazlett, B.A. Social behavior of the Paguridae and Diogenidae of Curacao. Stud. Fauna Curacao Other Caribb. Isl. 1966, 88, 1–143. [Google Scholar]
- Reese, E.R. Behavioral adaptations of intertidal hermit crabs. Am. Zool. 1969, 9, 343–355. [Google Scholar] [CrossRef]
- Vance, R.R. The role of shell adequacy in behavioral interactions involving hermit crabs. Ecology 1972, 53, 1075–1083. [Google Scholar] [CrossRef]
- Fotheringham, N. Population consequences of shell utilization by hermit crabs. Ecology 1976, 57, 570–578. [Google Scholar] [CrossRef]
- Hazlett, B.A. The behavioral ecology of hermit crabs. Annu. Rev. Ecol. Syst. 1981, 12, 1–22. [Google Scholar] [CrossRef]
- Markham, J.C. Notes on growth-patterns and shell-utilization of the hermit crab Pagurus bernhardus. Ophelia 1968, 5, 189–205. [Google Scholar] [CrossRef]
- Nyblade, C.F. Coexistence in Sympatric Hermit Crabs. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 1974. [Google Scholar]
- Abrams, P.; Nyblade, C.; Sheldon, S. Resource partitioning and competition for shells in a subtidal hermit crab species assemblage. Oecologia 1986, 69, 429–445. [Google Scholar] [CrossRef] [PubMed]
- Torjman, B.Z.; Iyengar, E.V. Increased potential for shell competition among intertidal hermit crabs (Pagurus beringanus and Pagurus granosimanus) in the rocky intertidal. J. Mar. Biol. Assoc. UK 2021, 101, 957–967. [Google Scholar] [CrossRef]
- Vermeij, G.J. Marine faunal dominance and molluscan shell form. Evolution 1974, 28, 656–664. [Google Scholar] [CrossRef]
- Bertness, M.D.; Cunningham, C. Crab shell-crushing predation and gastropod architectural defense. J. Exp. Mar. Biol. Ecol. 1981, 50, 213–230. [Google Scholar] [CrossRef]
- Spight, T.M. Availability and use of shells by intertidal hermit crabs. Biol. Bull. 1977, 152, 120–133. [Google Scholar] [CrossRef]
- Arce, E.; Alcaraz, G. Shell preference in a hermit crab: Comparison between a matrix of paired comparisons and a multiple-alternative experiment. Mar. Biol. 2012, 159, 853–862. [Google Scholar] [CrossRef]
- Alcaraz, G.; Toledo, B.; Burciaga, L.M. The energetic costs of living in the surf and impacts on zonation of shells occupied by hermit crabs. J. Exp. Biol. 2020, 223, jeb222703. [Google Scholar] [CrossRef]
- Briffa, M.; Elwood, R.W. Metabolic consequences of shell choice in Pagurus bernhardus: Do hermit crabs prefer cryptic or portable shells? Behav. Ecol. Sociobiol. 2005, 59, 143–148. [Google Scholar] [CrossRef]
- Alcaraz, G.; Garcia-Cabello, K.N. Feeding and metabolic compensations in response to different foraging costs. Hydrobiologia 2017, 787, 217–227. [Google Scholar] [CrossRef]
- Abrams, P.A. Resource partitioning and competition for shells between intertidal hermit crabs on the outer coast of Washington. Oecologia 1987, 72, 248–258. [Google Scholar] [CrossRef]
- Cote, I.M.; Reverdy, B.; Cooke, P.K. Less choosy or different preference? Impact of hypoxia on hermit crab shell assessment and selection. Anim. Behav. 1998, 56, 867–873. [Google Scholar] [CrossRef]
- Elwood, R.W.; Marks, N.; Dick, J.T.A. Consequences of shell-species preferences for female reproductive success in the hermit crab Pagurus bernhardus. Mar. Biol. 1995, 123, 431–434. [Google Scholar] [CrossRef]
- Caruso, T.; Chemello, R. The size and shape of shells used by hermit crabs: A multivariate analysis of Clibanarius erythropus. Acta Oecologica 2009, 35, 349–354. [Google Scholar] [CrossRef]
- Reese, E.S. The behavioral mechanisms underlying shell selection by hermit crabs. Behaviour 1963, 21, 78–126. [Google Scholar] [CrossRef]
- Lopes da Silva, D.J.; Oliveira, T.A.; Oliveira, A. Body composition and food intake in athletes and non athletes portuguese male adolescents. In Stochastic Modeling Data Analysis & Statistical Applications, 2nd ed.; Filus, L., Oliveira, T., Skiadas, C.H., Eds.; ISAST: Groningen, The Netherlands, 2015; pp. 215–224. [Google Scholar]
- Turra, A.; Leite, F.P.P. Shell-size selection by intertidal sympatric hermit crabs. Mar. Biol. 2004, 145, 251–257. [Google Scholar] [CrossRef]
- LaBarbera, M.; Merz, R.A. Postmortem changes in strength of gastropod shells: Evolutionary implications for hermit crabs, snails, and their mutual predators. Paleobiology 1992, 18, 367–377. [Google Scholar] [CrossRef]
- Pechenik, J.A.; Hsieh, J.; Owara, S.; Wong, P.; Marshall, D.; Untersee, S.; Li, W. Factors selecting for avoidance of drilled shells by the hermit crab Pagurus longicarpus. J. Exp. Mar. Biol. Ecol. 2001, 262, 75–89. [Google Scholar] [CrossRef]
- Billock, W.L.; Dunbar, S.G. Shell and food acquisition behaviors: Evidence for contextual decision hierarchies in hermit crabs. J. Exp. Mar. Biol. Ecol. 2011, 398, 26–32. [Google Scholar] [CrossRef]
- Angel, J.E. Effects of shell fit on the biology of the hermit crab Pagurus longicarpus (Say). J. Exp. Mar. Biol. Ecol. 2000, 243, 169–184. [Google Scholar] [CrossRef]
- Renganathan, V. Overview of Bayesian Approach to Statistical Methods. 2022; pp. 10–76. Available online: https://www.amazon.com.au/Overview-Bayesian-Approach-Statistical-Methods/dp/9356201188 (accessed on 16 April 2024).
- Hahn, D.R. Hermit crab shell use patterns: Response to previous shell experience and to water flow. J. Exp. Mar. Biol. Ecol. 1998, 228, 35–51. [Google Scholar] [CrossRef]
- Bell, J.J. Influence of occupant microhabitat on the composition of encrusting communities on gastropod shells. Mar. Biol. 2005, 147, 653–661. [Google Scholar] [CrossRef]
- Hazlett, B.A. Daily movements of the hermit crab Clibanarius vittatus. Bull. Mar. Sci. 1981, 31, 177–183. [Google Scholar]
- Doake, S.; Scantlebury, M.; Elwood, R.W. The costs of bearing arms and armour in the hermit crab Pagurus bernhardus. Anim. Behav. 2010, 80, 637–642. [Google Scholar] [CrossRef]
- Daly-Schveitzer, S.; Beugnon, G.; Lachaud, J.P. Prey weight and overwhelming difficulty impact the choice of retrieval strategy in the Neotropical ant Gnamptogenys sulcata (F. Smith). Insectes Sociaux 2007, 54, 319–328. [Google Scholar] [CrossRef]
- Nguyen, V.; Lilly, B.; Castro, C.E. The exoskeletal structure and tensile loading behavior of an ant neck joint. J. Biomech. 2014, 47, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Segre, P.S.; Taylor, E.D. Large ants do not carry their fair share: Maximal load-carrying performance of leaf-cutter ants (Atta cephalotes). J. Exp. Biol. 2019, 222, jeb199240. [Google Scholar] [CrossRef]
- Young, A.M. Differential utilization of gastropod shells by three hermit crab species in North Inlet, South Carolina, U.S.A. In Crustaceana. Supplement, No. 5, Studies on Decapoda (Biology, Ecology, Morphology, and Systematics); Brill: Leiden, The Netherlands, 1979; pp. 101–104. [Google Scholar]
- Raup, D.M.; Michelson, A. Theoretical morphology of the coiled shell. Science 1965, 147, 1294–1295. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, G.; Chávez-Solís, C.E.; Kruesi, K. Mismatch between body growth and shell preference in hermit crabs is explained by protection from predators. Hydrobiologia 2015, 743, 151–156. [Google Scholar] [CrossRef]
- Micheli, F. Behavioural plasticity in prey-size selectivity of the blue crab Callinectes sapidus feeding on bivalve prey. J. Anim. Ecol. 1995, 64, 63–74. [Google Scholar] [CrossRef]
- Billock, W.L.; Dunbar, S.G. Influence of motivation on behaviour in the hermit crab, Pagurus samuelis. J. Mar. Biol. Assoc. UK 2009, 89, 775–779. [Google Scholar] [CrossRef]
- Long, Q.; Liu, J.; Sun, Y.; Yang, Z.; Tang, B.; Cheng, Y. The effect of food deprivation on foraging behavior and digestive and metabolic capacities of the Chinese mitten crab, Eriocheir sinensis. Fishes 2023, 8, 47. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Dong, S. A comparative study of the effect of starvation regimes on the foraging behavior of Portunus trituberculatus and Charybdis japonica. Physiol. Behav. 2015, 151, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, J.C. Allometric studies on energy relationships in the spider crab Libinia emarginata (Leach). Biol. Bull. 1974, 147, 257–273. [Google Scholar] [CrossRef]
- Orians, G.H.; King, C.E. Shell selection and invasion rates of some Pacific hermit crabs. Pac. Sci. 1964, 18, 297–306. [Google Scholar]
- Rittschof, D.; Sarrica, J.; Rubenstein, D. Shell dynamics and microhabitat selection by striped legged hermit crabs, Clibanarius vittatus (Bosc). J. Exp. Mar. Biol. Ecol. 1995, 192, 157–172. [Google Scholar] [CrossRef]
- Vance, R.R. Competition and mechanism of coexistence in three sympatric species of intertidal hermit crabs. Ecology 1972, 53, 1062–1074. [Google Scholar] [CrossRef]
- Garcia, R.B.; Mantelatto, F.L. Shell selection by the tropical hermit crab Calcinus tibicen (Herbst, 1791) (Anomura, Diogenidae) from Southern Brazil. J. Exp. Mar. Biol. Ecol. 2001, 265, 1–14. [Google Scholar] [CrossRef]
- Alcaraz, G.; Arce, E. Predator discrimination in the hermit crab Calcinus californiensis: Tight for shell breakers, loose for shell peelers. Oikos 2017, 126, 1299–1307. [Google Scholar] [CrossRef]
- Rotjan, R.D.; Blum, J.; Lewis, S.M. Shell choice in Pagurus longicarpus hermit crabs: Does predation threat influence shell selection behavior? Behav. Ecol. Sociobiol. 2004, 56, 171–176. [Google Scholar]
- Krieger, J.; Hörnig, M.K.; Laidre, M.E. Shells as ‘extended architecture’: To escape isolation, social hermit crabs choose shells with the right external architecture. Anim. Cogn. 2020, 23, 1177–1187. [Google Scholar]
- Herreid, C.F.; Full, R.J. Energetics of hermit crabs during locomotion: The costs of carrying a shell. J. Exp. Biol. 1996, 120, 297–308. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species of Shell | Shell Length | Aperture Length | Shell Weight | |||
|---|---|---|---|---|---|---|
| Average | 1SE | Average | 1SE | Average | 1SE | |
| Calliostoma ligatum | 18.57 | 1.28 | 9.30 | 0.34 | 2.13 | 0.21 |
| Nucella lamellosa | 33.62 | 0.78 | 9.93 | 0.30 | 5.42 | 0.35 |
| Lirabuccinum dirum | 29.33 | 1.15 | 10.01 | 0.41 | 1.97 | 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torjman, B.Z.; Iyengar, E.V. Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus. Hydrobiology 2024, 3, 74-84. https://doi.org/10.3390/hydrobiology3020006
Torjman BZ, Iyengar EV. Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus. Hydrobiology. 2024; 3(2):74-84. https://doi.org/10.3390/hydrobiology3020006
Chicago/Turabian StyleTorjman, Brooke Z., and Erika V. Iyengar. 2024. "Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus" Hydrobiology 3, no. 2: 74-84. https://doi.org/10.3390/hydrobiology3020006
APA StyleTorjman, B. Z., & Iyengar, E. V. (2024). Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus. Hydrobiology, 3(2), 74-84. https://doi.org/10.3390/hydrobiology3020006