Socio-Thermodynamics—Evolutionary Potentials in a Population of Hawks and Doves

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Evolutionary Equilibria
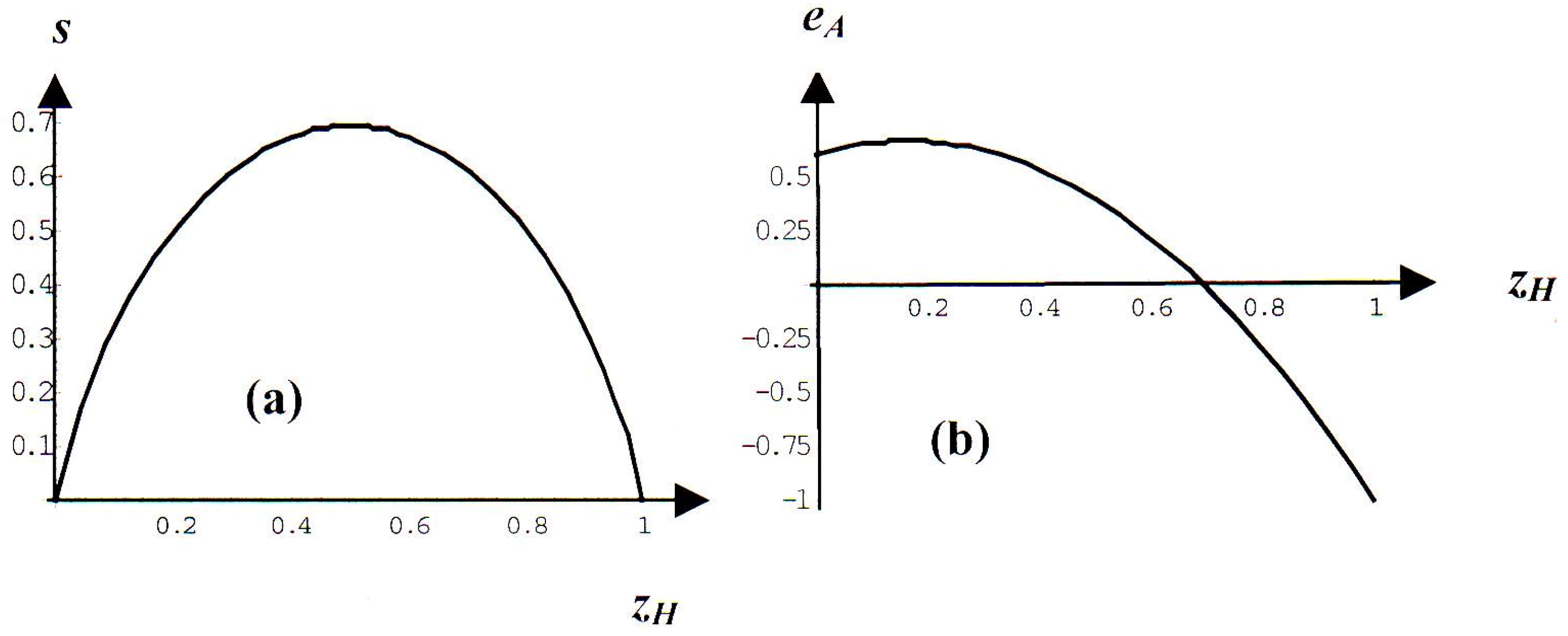
2.1. Evolutionary Entropic Drift toward an Equi-Distribution

2.2. Gain-Driven Drift toward an Evolutionarily Stable Distribution
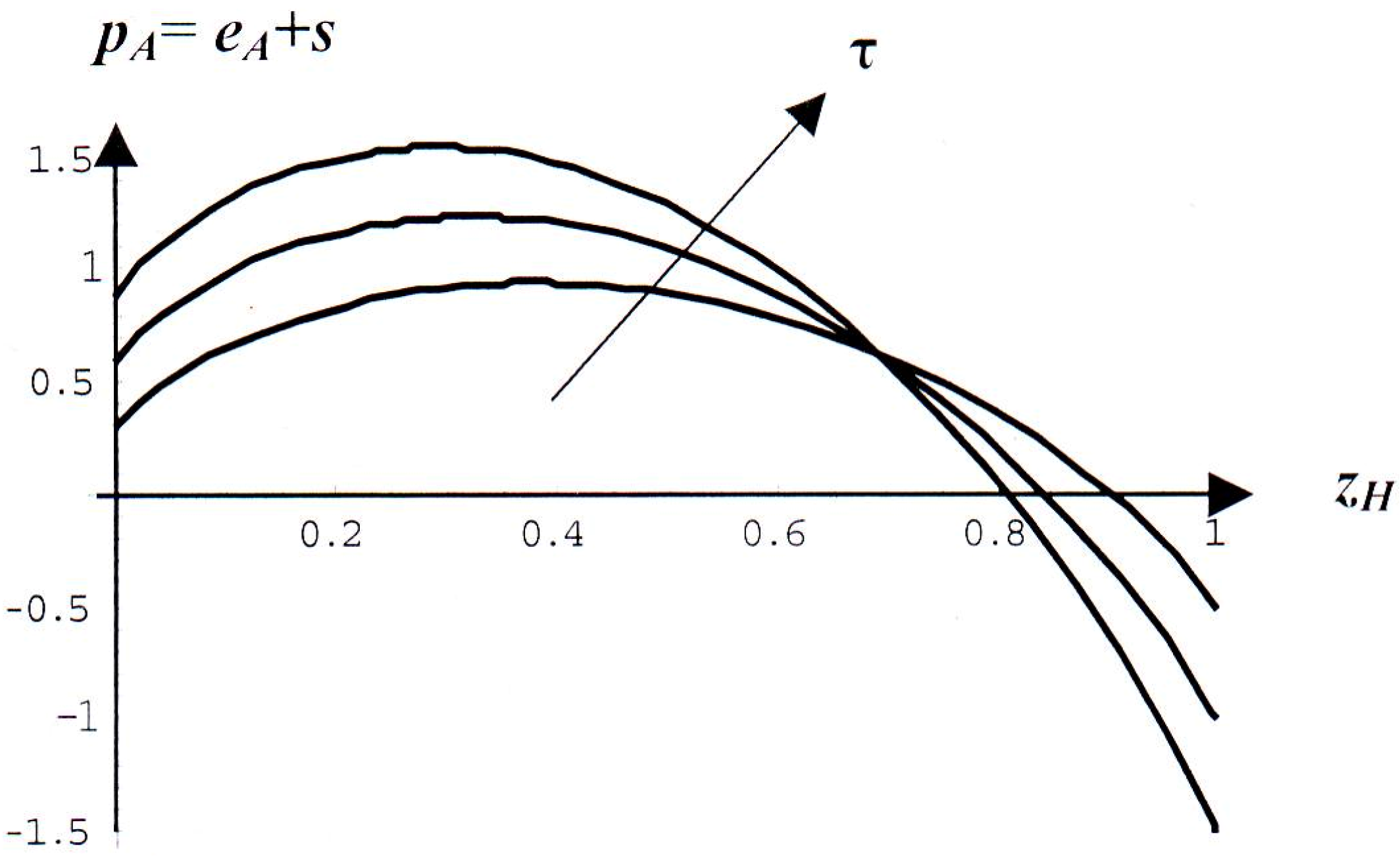
2.3. Combined Entropic and Gain Driven Trends

2.4. Analogy with Thermodynamics of Mixtures
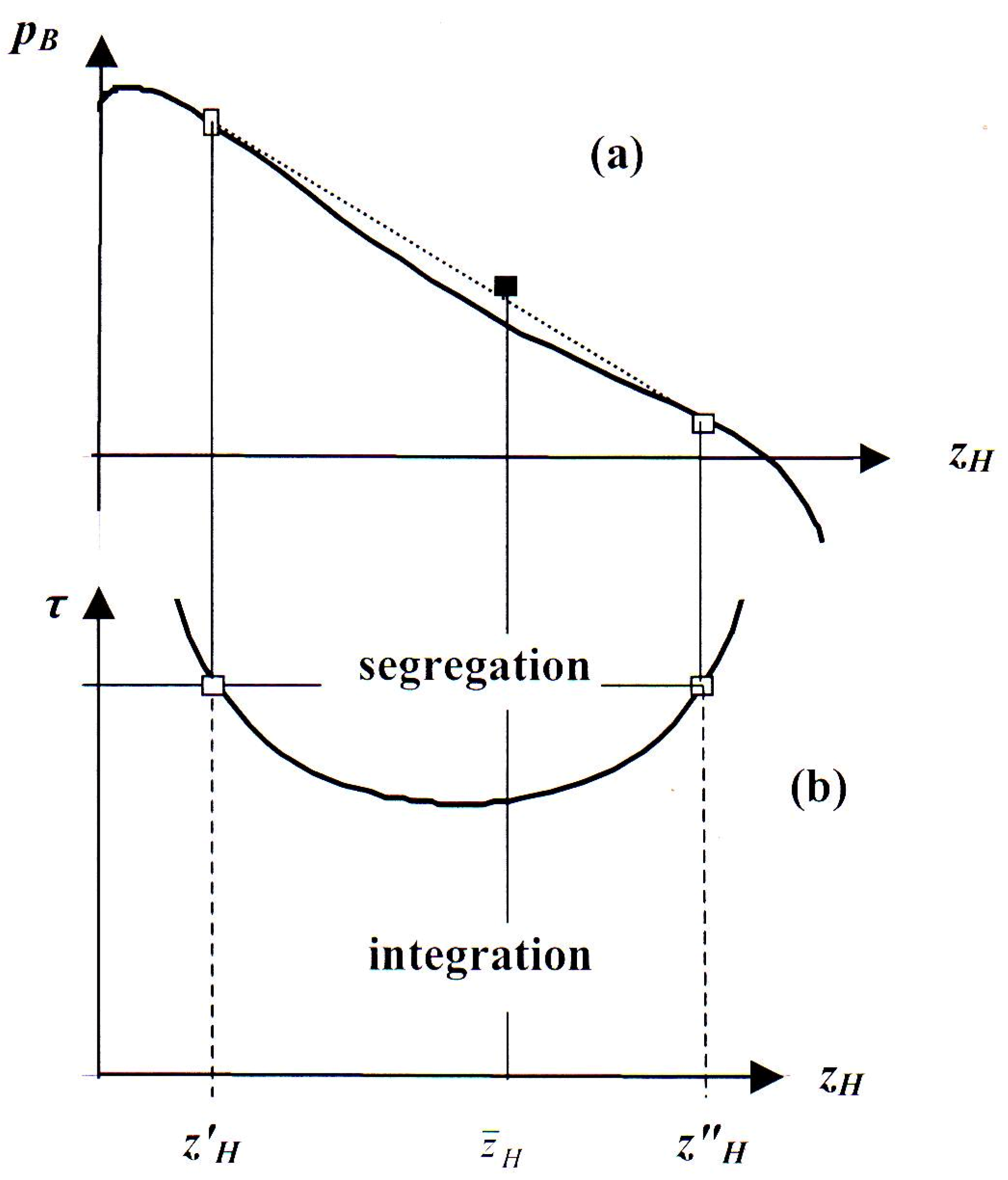
3. Segregation of Hawks and Doves
3.1. Non-Convex Potentials. Concavification

3.2. Thermodynamic Analogy
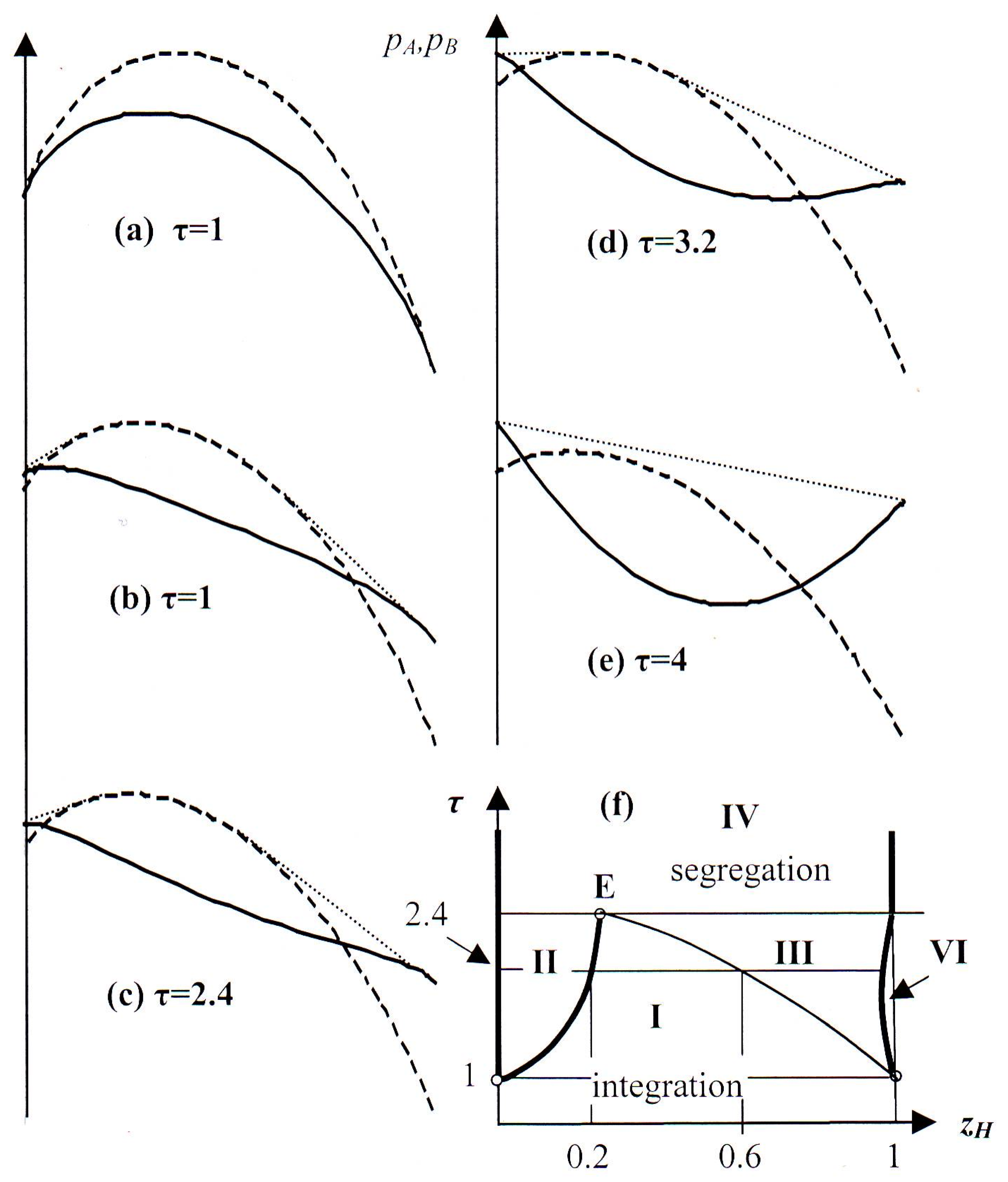
4. Segregation in a Population with a Choice of Strategies
4.1. Intersecting Graphs pA and pB
- Homogeneous population employing Strategy A for all hawk fractions.
- Segregated population with two types of colonies: Dove-rich colonies employing Strategy B and colonies with moderate hawk fractions and Strategy A.
- Segregated population with two types of colonies: Hawk-rich ones with Strategy B and colonies with moderate hawk fractions and Strategy A.
- Segregated population with hawk-rich and dove-rich colonies both employing Strategy B.
- Homogeneous dove-rich populations with Strategy B. (This region is not visible in Figure 4f, because on the scale of the figure it lies virtually on the τ-axis.)
- Homogeneous hawk-rich populations with Strategy B.

4.2. Analogy with Thermodynamics of Solutions and Alloys
References and Notes
- Maynard-Smith, J.; Price, G.R. The logic of animal conflict. Nature 1973, 246, 15–18. [Google Scholar] [CrossRef]
- Straffin, P.D. Game Theory and Strategy; New Mathematical Library, The Mathematical Association of America: Washington, DC, USA, 1993. [Google Scholar]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Müller, I. Socio-thermodynamics—Integration and segregation in a population. Continuum. Mech. Therm. 2002, 14, 389–404. [Google Scholar]
- Müller, I. A History of Thermodynamics—The Doctrine of Energy and Entropy; Springer: Heidelberg, Germany, 2007. [Google Scholar]
- Müller, I. Integration and segregation in a population—A thermodynamicst’s view. In Thermodynamics and the Destruction of Resources; Bakshi, B.R., Gutowski, T.G., Sekulic, D.P., Eds.; Cambridge University Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Mimkes, J. Binary alloys as a model for the multicultural society. J. Therm. Anal. 1995, 43, 512–537. [Google Scholar] [CrossRef]
- We let the total number of N birds be unchanged from generation to generation.
- Here and in the sequel we adopt the Stirling formula in order to replace factorials of large numbers: ln a! ≈ a ln a − a.
- This strategy was invented by the biologists J. Maynard-Smith and G.R. Price [1] and Dawkins [3] in order to show that a mixed population of two competing species may be evolutionarily stable. Here I modify the strategy of those biologists in a trivial manner by introducing the price τ, which I consider as dimensionless. In anticipation of misunderstandings or criticism I say this: Dawkins does not consider eA in (6) as relevant, because he refuses the practicality of pacts or conspiracies which favour the gain for the population as a whole rather than the gain of the selfish individual. The relevance of eA requires enforcement of social measures to make the gain of the population a criterion for behaviour. We assume that such measures are agreed upon in the population.
- τ = 1 is a reference price in which both strategies coincide, except for the grab-and-run feature of strategy B. Penalties for either fighting or posturing should never turn into rewards for whatever permissible value of τ. This condition imposes an upper bound on τ: 0 < τ< 4.33. That constraint could be avoided, if we allowed non-linear penalty reductions which we do not do for the sake of simplicity.
- If there were evolution, the phase fraction would shift to the abscissa of the maximum of pB over subsequent generations.
- It is fortunate that (14) lends itself to such a simple graphical interpretation, since an analytic solution is impossible because of the ln-terms in the entropic part of pB(zH). Actually, however, in Figure 3 the influence of entropy is arbitrarily enhanced by a factor 2, because otherwise the concave parts of pB(zH) would not be sufficiently well pronounced to make the construction clear.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Müller, I. Socio-Thermodynamics—Evolutionary Potentials in a Population of Hawks and Doves. Entropy 2012, 14, 1285-1295. https://doi.org/10.3390/e14071285
Müller I. Socio-Thermodynamics—Evolutionary Potentials in a Population of Hawks and Doves. Entropy. 2012; 14(7):1285-1295. https://doi.org/10.3390/e14071285
Chicago/Turabian StyleMüller, Ingo. 2012. "Socio-Thermodynamics—Evolutionary Potentials in a Population of Hawks and Doves" Entropy 14, no. 7: 1285-1295. https://doi.org/10.3390/e14071285
APA StyleMüller, I. (2012). Socio-Thermodynamics—Evolutionary Potentials in a Population of Hawks and Doves. Entropy, 14(7), 1285-1295. https://doi.org/10.3390/e14071285