Dynamical Structure of a Traditional Amazonian Social Network

Abstract
:Se hacía chicha, después se juntaba la gente y después todita la gente estaba borracha y cantaban las canciones de los animales: del chancho de tropa, del marimono, del maneche, y seguían cantando... Chicha was made, and the people got together, and then everyone was drunk and singing the songs of the animals—the white-lipped peccary, the spider monkey, the howler monkey—and on they sang...–Felipe Huallata [1]
1. Introduction
1.1. Evolutionary Background and Motivation
1.2. Current Aims and Approach
2. Methods
2.1. Population and Field Setting
2.2. Data Collection and Preparation
2.3. Multi-Level Regression

2.4. Time-Series Analysis
2.4.1. Estimating Conditional-Action Reciprocity
2.4.2. A Three-Level Hierarchy of Null Models for Conditional-Action Reciprocity
3. Results


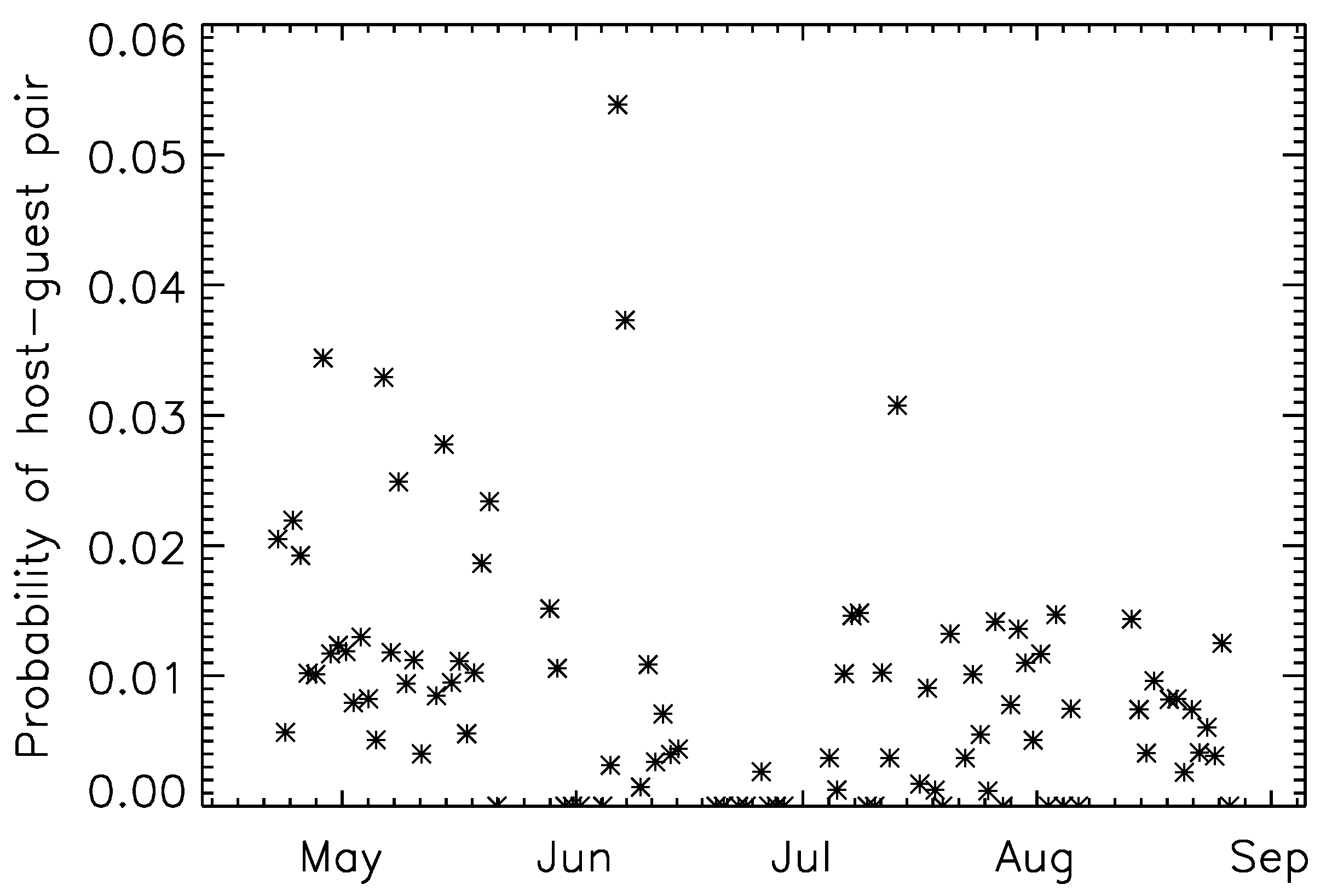
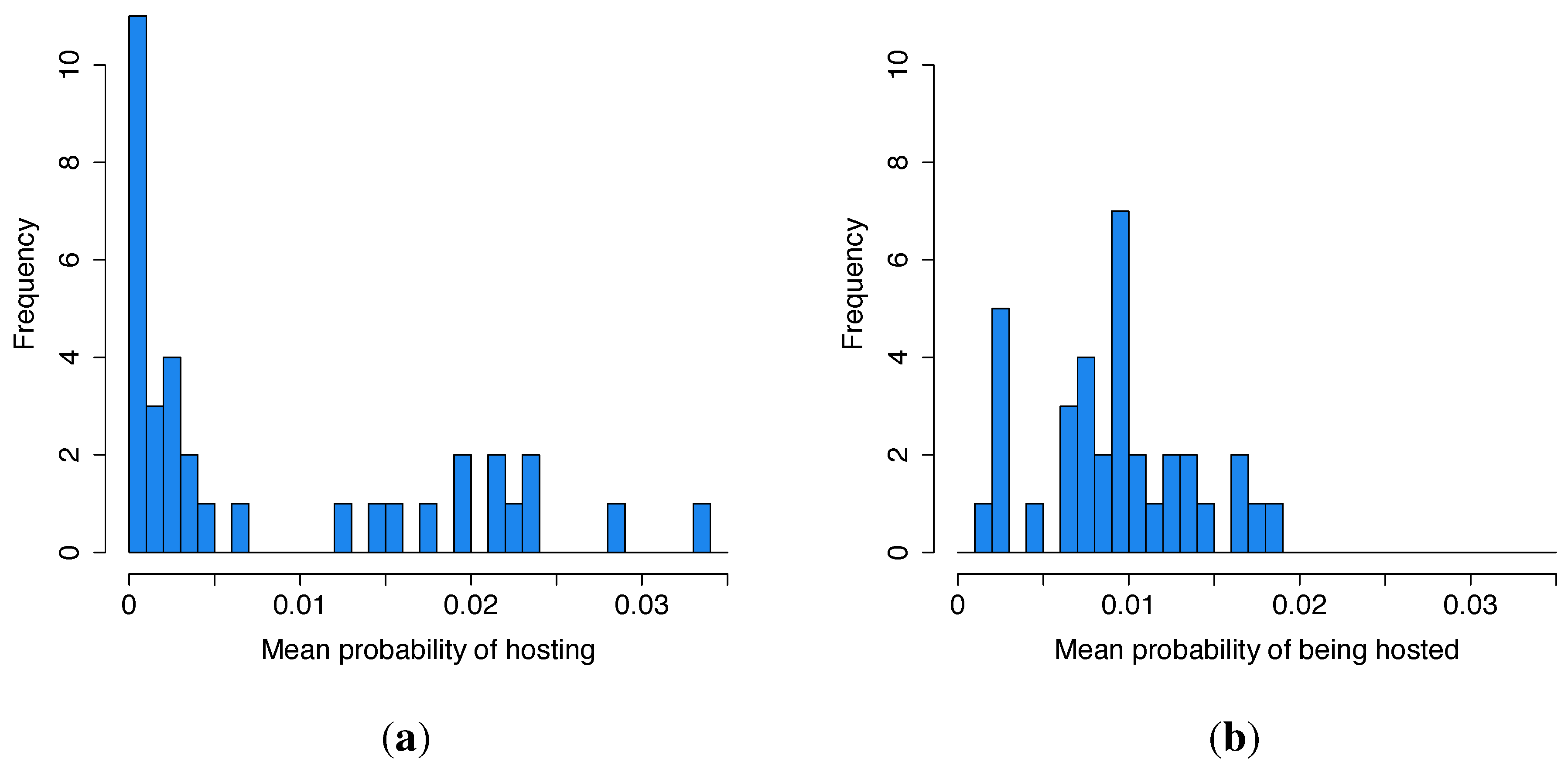
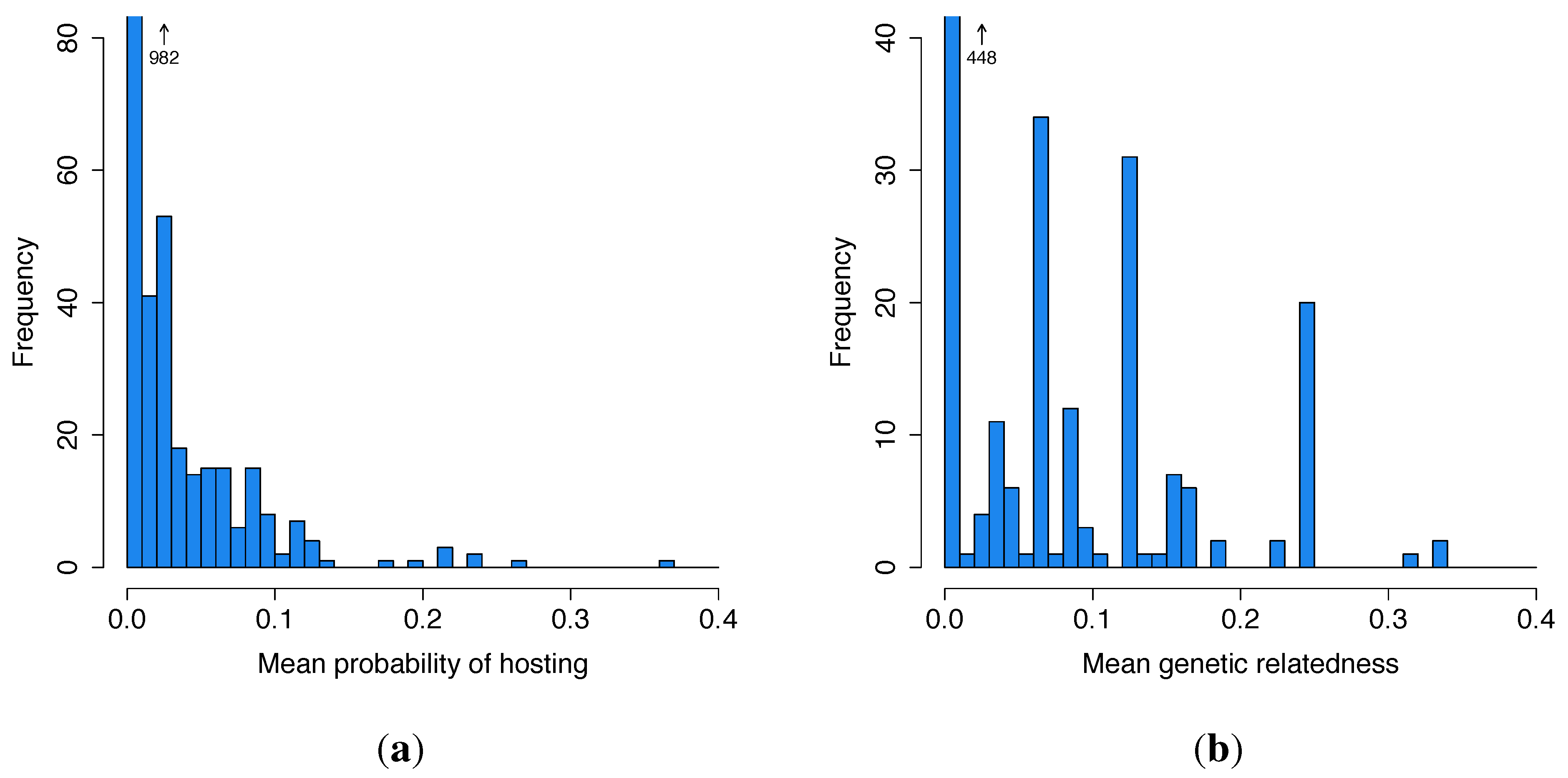
3.1. Summary Statistics
3.2. Multi-Level Regression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | B | C | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Random effects | Var(α) | p | Var(α) | p | Var(α) | p | |||
| Host identity () | 2.581 | <0.001 | 1.875 | <0.001 | 3.262 | <0.001 | |||
| Guest identity () | 0.046 | 0.008 | 0.018 | 0.082 | 0.135 | <0.001 | |||
| Host-guest dyad () | 2.733 | <0.001 | 2.817 | <0.001 | 1.166 | <0.001 | |||
| Predictors | β | p | β | p | β | p | |||
| Guest report only | −0.972 | 0.378 | <0.001 | −0.964 | 0.381 | <0.001 | −0.956 | 0.384 | <0.001 |
| Host age | 0.223 | 1.250 | <0.001 | 0.293 | 1.340 | <0.001 | |||
| Host age | −0.002 | 0.998 | <0.001 | −0.003 | 0.997 | <0.001 | |||
| Guest age | −0.009 | 0.991 | <0.001 | −0.007 | 0.993 | 0.006 | |||
| Log distance | −1.050 | 0.350 | <0.001 | ||||||
| AIC | 4,565 | 4,560 | 4,365 | ||||||
| D | E | F | |||||||
| Random effects | Var(α) | p | Var(α) | p | Var(α) | p | |||
| Host identity () | 2.778 | <0.001 | 3.164 | <0.001 | 3.276 | <0.001 | |||
| Guest identity () | 0.129 | <0.001 | 0.111 | <0.001 | 0.032 | 0.030 | |||
| Host-guest dyad () | 1.120 | <0.001 | 0.824 | <0.001 | 0.619 | <0.001 | |||
| Predictors | β | p | β | p | β | p | |||
| Guest report only | −0.965 | 0.381 | <0.001 | −0.954 | 0.385 | <0.001 | −0.959 | 0.383 | <0.001 |
| Host age | 0.229 | 1.257 | <0.001 | 0.268 | 1.307 | <0.001 | 0.276 | 1.318 | <0.001 |
| Host age | −0.002 | 0.998 | <0.001 | −0.003 | 0.997 | <0.001 | −0.003 | 0.997 | <0.001 |
| Guest age | −0.007 | 0.993 | 0.008 | −0.007 | 0.993 | 0.008 | −0.009 | 0.991 | <0.001 |
| Log distance | −0.618 | 0.539 | <0.001 | −0.520 | 0.595 | <0.001 | |||
| Kinship | 14.02 | 1.2E6 | <0.001 | 8.860 | 7045 | <0.001 | 8.435 | 4606 | <0.001 |
| Rate j hosts i | 19.99 | 4.8E8 | <0.001 | ||||||
| Kinship × Rate j hosts i | −64.86 | 7E-29 | 0.234 | ||||||
| AIC | 4,354 | 4,294 | 4,255 |
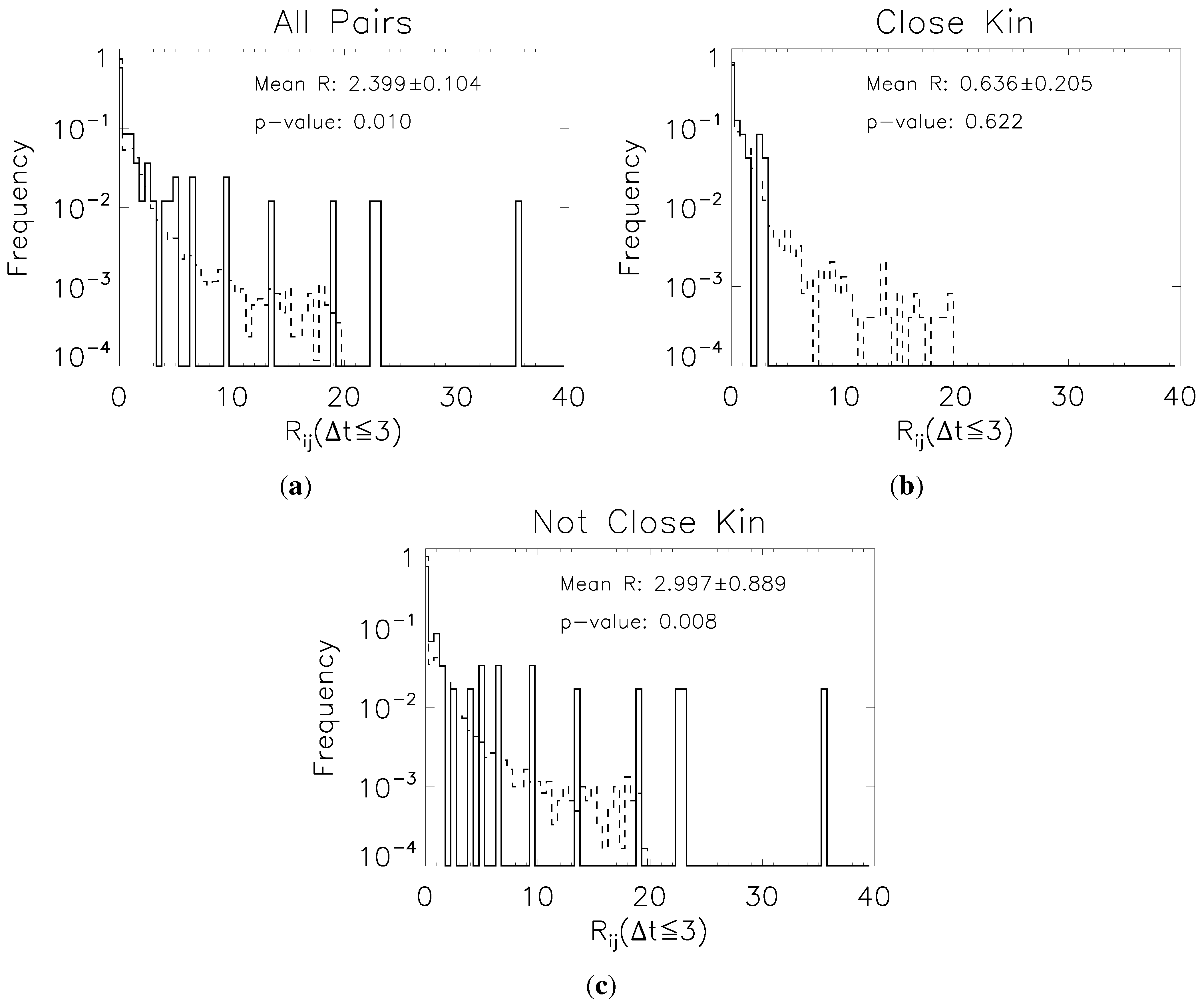
3.3. Time-Series Analysis


3.4. Discussion
4. Summary and Conclusion
Acknowledgments
Conflicts of Interest
References and Notes
- Iamele, G. Palabras Antiguas y Nuevas del Río Quiquibey en la Amazonia Boliviana; (in Spanish). Programa Regional de Apoyo a los Pueblos Indíginas de la Cuenca del Amazonas: La Paz, Bolivia, 2001. [Google Scholar]
- Silk, J.B. The Adaptive Value of Sociality. In The Evolution of Primate Societies; Mitani, J.C., Call, J., Kappeler, P.M., Palombit, R.A., Silk, J.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2012; pp. 554–564. [Google Scholar]
- Jackson, M.O.; Watts, A. The evolution of social and economic networks. J. Econ. Theory 2002, 106, 265–295. [Google Scholar] [CrossRef]
- Kappeler, P.M.; Barrett, L.; Blumstein, D.T.; Clutton-Brock, T.H. Constraints and flexibility in mammalian social behaviour: Introduction and synthesis. Philos. Trans. R. Soc. B 2013, 368, 20120337. [Google Scholar] [CrossRef] [PubMed]
- Doreian, P.; Stokman, F. Evolution of Social Networks; Routledge: Amsterdam, The Netherlands, 1997; Volume 1. [Google Scholar]
- Newman, M. Networks: An Introduction; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Flack, J.C. Multiple time-scales and the developmental dynamics of social systems. Philos. Trans. R. Soc. B 2012, 367, 1802–1810. [Google Scholar] [CrossRef] [PubMed]
- DeDeo, S.; Krakauer, D.C.; Flack, J.C. Inductive game theory and the dynamics of animal conflict. PLoS Comput. Biol. 2010, 6, e1000782. [Google Scholar] [CrossRef] [PubMed]
- DeDeo, S.; Krakauer, D.C.; Flack, J.C. Evidence of strategic periodicities in collective conflict dynamics. J. R. Soc. Interface 2011, 8, 1260–1273. [Google Scholar] [CrossRef] [PubMed]
- Mowat, L. Cassava and Chicha: Bread and Beer of the Amazonian Indians; Shire Publications: Aylesbury, UK, 1989. [Google Scholar]
- Hamilton, W.D. The genetical evolution of social behaviour. I. J. Theor. Biol. 1964, 7, 1–16. [Google Scholar] [CrossRef]
- Trivers, R.L. The evolution of reciprocal altruism. Q. Rev. Biol. 1971, 46, 35–57. [Google Scholar] [CrossRef]
- Gurven, M. To give and to give not: The behavioral ecology of human food transfers. Behav. Brain Sci. 2004, 27, 543–559. [Google Scholar] [CrossRef]
- West, S.A.; Griffin, A.S.; Gardner, A. Evolutionary explanations for cooperation. Curr. Biol. 2007, 17, R661–R672. [Google Scholar] [CrossRef] [PubMed]
- Langergraber, K.E. Cooperation among Kin. In The Evolution of Primate Societies; Mitani, J.C., Call, J., Kappeler, P.M., Palombit, R.A., Silk, J.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2012; pp. 491–513. [Google Scholar]
- Griffin, A.S.; West, S.A. Kin discrimination and the benefit of helping in cooperatively breeding vertebrates. Science 2003, 302, 634–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourke, A.F. The validity and value of inclusive fitness theory. Proc. R. Soc. B 2011, 278, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Service, E.R. Primitive Social Organization; Random House: New York, NY, USA, 1962. [Google Scholar]
- Hames, R. Garden labor exchange among the Ye’kwana. Ethol. Sociobiol. 1987, 8, 259–284. [Google Scholar] [CrossRef]
- Hooper, P.L. The Structure of Energy Production and Redistribution Among Tsimane’ Forager-Horticulturalists; University of New Mexico: Albuquerque, NM, USA, 2011. [Google Scholar]
- Koster, J. Interhousehold meat sharing among Mayangna and Miskito horticulturalists in Nicaragua. Hum. Nat. 2011, 22, 394–415. [Google Scholar] [CrossRef] [PubMed]
- Mauss, M. The Gift: The Form and Reason for Exchange in Archaic Societies. W.W. Norton: New York, NY, USA, 1990. [Google Scholar]
- Axelrod, R.; Hamilton, W.D. The evolution of cooperation. Science 1981, 211, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Gurven, M. The evolution of contingent cooperation. Curr. Anthropol. 2006, 47, 185–192. [Google Scholar] [CrossRef]
- Reciprocity might also simply refer to the frequency with which the social partners nominated by an individual also nominate that individual in turn; here we are interested in reciprocity in networks in which directed edges represent tangible flows of benefits, goods, or services between nodes.
- Rao, A.R.; Bandyopadhyay, S. Measures of reciprocity in a social network. Sankhyā: Indian J. Stat. Ser. A 1987, 49, 141–188. [Google Scholar]
- Wasserman, S.; Faust, K. Social Network Analysis: Methods and Applications; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Jaeggi, A.V.; Gurven, M. Reciprocity explains food sharing in humans and other primates independent of kin selection and tolerated scrounging: A phylogenetic meta-analysis. Proc. R. Soc. B 2013, 280, 20131615. [Google Scholar] [CrossRef] [PubMed]
- Gilby, I.C. Cooperation among Non-Kin: Reciprocity, Markets, and Mutualism. In The Evolution of Primate Societies; Mitani, J.C., Call, J., Kappeler, P.M., Palombit, R.A., Silk, J.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2012; pp. 514–530. [Google Scholar]
- Jana, R.; Bandyopadhyay, S.; Choudhuri, A.K. Reciprocity among farmers in farming system research: Application of social network analysis. J. Hum. Ecol. 2013, 41, 45–51. [Google Scholar]
- Silk, J.B. Cooperation without Counting: The Puzzle of Friendship. In Genetic and Cultural Evolution of Cooperation; Hammerstein, P., Ed.; MIT: Cambridge, MA, USA, 2003; pp. 37–54. [Google Scholar]
- Hruschka, D.J. Friendship: Development, Ecology, and Evolution of a Relationship; University of California Press: Berkeley, CA, USA, 2010. [Google Scholar]
- Kasper, C.; Fitzherbert, E.; Borgerhoff Mulder, M. Who helps and why? Cooperative networks in Mpimbwe; Working Paper; University of California Davis: Davis, CA, USA, 2013. [Google Scholar]
- Hames, R.; McCabe, C. Meal sharing among the Ye’kwana. Hum. Nat. 2007, 18, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hemelrijk, C.K. Support for being groomed in long-tailed macaques, Macaca fascicularis. Anim. Behav. 1994, 48, 479–481. [Google Scholar] [CrossRef]
- Melis, A.P.; Hare, B.; Tomasello, M. Do chimpanzees reciprocate received favours? Anim. Behav. 2008, 76, 951–962. [Google Scholar] [CrossRef]
- Koyama, N.; Caws, C.; Aureli, F. Interchange of grooming and agonistic support in chimpanzees. Int. J. Primatol. 2006, 27, 1293–1309. [Google Scholar] [CrossRef]
- Jaeggi, A.V.; de Groot, E.; Stevens, J.M.; van Schaik, C.P. Mechanisms of reciprocity in primates: Testing for short-term contingency of grooming and food sharing in bonobos and chimpanzees. Evol. Hum. Behav. 2012, 34, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Schino, G.; Aureli, F. Reciprocal altruism in primates: Partner choice, cognition, and emotions. Adv. Stud. Behav. 2009, 39, 45–69. [Google Scholar]
- Frank, R.E.; Silk, J.B. Impatient traders or contingent reciprocators? Evidence for the extended time-course of grooming exchanges in baboons. Behaviour 2009, 146, 1123–1135. [Google Scholar] [CrossRef]
- Gomes, C.M.; Mundry, R.; Boesch, C. Long-term reciprocation of grooming in wild West African chimpanzees. Proc. R. Soc. B 2009, 276, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Sabbatini, G.; Vizioli, A.D.B.; Visalberghi, E.; Schino, G. Food transfers in capuchin monkeys: An experiment on partner choice. Biol. Lett. 2012, 8, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Tiddi, B.; Aureli, F.; di Sorrentino, E.P.; Janson, C.H.; Schino, G. Grooming for tolerance? Two mechanisms of exchange in wild tufted capuchin monkeys. Behav. Ecol. 2011, 22, 663–669. [Google Scholar] [CrossRef]
- McElreath, R.; Boyd, R. Mathematical Models of Social Evolution: A Guide for the Perplexed; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- McGlothlin, J.W.; Moore, A.J.; Wolf, J.B.; Brodie, E.D., III. Interacting phenotypes and the evolutionary process. III. Social evolution. Evolution 2010, 64, 2558–2574. [Google Scholar] [CrossRef] [PubMed]
- Akçay, E.; van Cleve, J. Behavioral responses in structured populations pave the way to group optimality. Am. Nat. 2012, 179, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Allen-Arave, W.; Gurven, M.; Hill, K. Reciprocal altruism, rather than kin selection, maintains nepotistic food transfers on an Ache reservation. Evol. Hum. Behav. 2008, 29, 305–318. [Google Scholar] [CrossRef]
- Nolin, D.A. Kin preference and partner choice. Hum. Nat. 2011, 22, 156–176. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.A.; Sigmund, K. Evolution of indirect reciprocity. Nature 2005, 437, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Macfarlan, S.J.; Remiker, M.; Quinlan, R. Competitive altruism explains labor exchange variation in a Dominican community. Curr. Anthropol. 2012, 53, 118–124. [Google Scholar] [CrossRef]
- Gurven, M.; Allen-Arave, W.; Hill, K.; Hurtado, M. It’s a wonderful life: Signaling generosity among the Ache of Paraguay. Evol. Hum. Behav. 2000, 21, 263–282. [Google Scholar] [CrossRef]
- Bowles, S.; Gintis, H. The evolution of strong reciprocity: Cooperation in heterogeneous populations. Theor. Popul. Biol. 2010, 65, 17–28. [Google Scholar] [CrossRef]
- Bowles, S.; Gintis, H. A Cooperative Species: Human Reciprocity and its Evolution; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Smith, E.A.; Bird, R.L.B. Turtle hunting and tombstone opening: Public generosity as costly signaling. Evol. Hum. Behav. 2000, 21, 245–261. [Google Scholar] [CrossRef]
- Gintis, H.; Smith, E.A.; Bowles, S. Costly signaling and cooperation. J. Theor. Biol. 2001, 213, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Gurven, M.; Stieglitz, J.; Hooper, P.L.; Gomes, C.; Kaplan, H. From the womb to the tomb: The role of transfers in shaping the evolved human life history. Exp. Gerontol. 2012, 47, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Gurven, M.; Kaplan, H.; Zelada Supa, A. Mortality experience of Tsimane Amerindians of Bolivia: Regional variation and temporal trends. Am. J. Hum. Biol. 2007, 19, 376–398. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T.; Atkinson, E.; Sinnwell, J.; Matsumoto, M.; Schaid, D.; McDonnell, S. kinship2: Pedigree Functions, R Package Version 1.5.0; 2013. Available online: http://CRAN.R-project.org/package=kinship2 (accessed on 1 July 2013).
- The R Core Team. R: A Language and Environment for Statistical Computing, version 3.0.1; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 1 September 2013).
- Csárdi, G.; Nepusz, T. igraph: The igraph Software Package for Complex Network Research, R Package version 0.6.5; 2013. Available online: http://igraph.sf.net (accessed 1 September 2013).
- Bates, D.; Maechler, M.; Bolker, B. lme4: Linear Mixed-Effects Models Using S4 Classes, R Package version 0.999999-2; 2013. Available online: http://CRAN.R-project.org/package=lme4 (accessed 1 September 2013).
- If it were the case that family i hosting family j on a particular day made it more difficult to host another family k on the same day (e.g., due to limited space or chicha), then an alternative specification of Equation (1), such as the multinomial logistic, might be preferred in order to represent the negative interdependence of the two dyadic interactions. However, since two or more families (up to a maximum of 10) were hosted on 106 out of 193 risk days that one family hosted at least one other family, the multinomial specification—which forces an either-or choice between families to host—would be inappropriate to apply to the current dataset. We thank one of the reviewers for these insights.
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Hox, J. Multilevel Analysis: Techniques and Applications; Routledge Academic: New York, NY, USA, 2010. [Google Scholar]
- Pearl, J. Causality, 2nd ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- The frequencies of hosting and attendance are also associated with household distance from village center (β = -0.816, p = 0.239 for hosts; β = -1.003, p = 0.001 for guests) and mean relatedness to other families in the community (i.e., node strength in the kinship network; β = 29.58, p = 0.119 for hosts; β = 23.75, p < 0.001 for guests); these relationships, however, are more directly captured (and statistically obviated: p > 0.6) by the dyad-specfic distance and relatedness terms.
- Press, W. Numerical Recipes: The Art of Scientific Computing, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Blurton Jones, N.G. A selfish origin for human food sharing: Tolerated theft. Ethol. Sociobiol. 1984, 5, 1–3. [Google Scholar] [CrossRef]
- Gurven, M.; Kaplan, H. Determinants of time allocation across the lifespan: A theoretical model and an application to the Machiguenga and Piro of Peru. Hum. Nat. 2006, 17, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Bowles, S.; Polania-Reyes, S. Economic incentives and social preferences: Substitutes or complements? J. Econ. Lit. 2012, 50, 368–425. [Google Scholar] [CrossRef]
A. Appendix: Further Details on Statistical Estimation of Conditional-Action Reciprocity
A.1. Correction for Seasonality
A.2. Correction for Reporting
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hooper, P.L.; DeDeo, S.; Caldwell Hooper, A.E.; Gurven, M.; Kaplan, H.S. Dynamical Structure of a Traditional Amazonian Social Network. Entropy 2013, 15, 4932-4955. https://doi.org/10.3390/e15114932
Hooper PL, DeDeo S, Caldwell Hooper AE, Gurven M, Kaplan HS. Dynamical Structure of a Traditional Amazonian Social Network. Entropy. 2013; 15(11):4932-4955. https://doi.org/10.3390/e15114932
Chicago/Turabian StyleHooper, Paul L., Simon DeDeo, Ann E. Caldwell Hooper, Michael Gurven, and Hillard S. Kaplan. 2013. "Dynamical Structure of a Traditional Amazonian Social Network" Entropy 15, no. 11: 4932-4955. https://doi.org/10.3390/e15114932
APA StyleHooper, P. L., DeDeo, S., Caldwell Hooper, A. E., Gurven, M., & Kaplan, H. S. (2013). Dynamical Structure of a Traditional Amazonian Social Network. Entropy, 15(11), 4932-4955. https://doi.org/10.3390/e15114932