Searching for Conservation Laws in Brain Dynamics—BOLD Flux and Source Imaging

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Do Conservation Laws Rule the Brain?
2.1. The Importance of Conservation Laws
2.2. Possible Hints for Conservation Laws Ruling the Brain
3. Theory
3.1. Continuity Equations
3.2. Estimation of Fluxes and Sources from BOLD Data
3.3. Generalizations
4. Mapping BOLD Fluxes and Sources in a Motor Imagery Experiment
4.1. Motivation
4.2. Subjects
4.3. MRI
4.4. fMRI Data Analysis
- (1)
- Conventional BOLD amplitude mapping: The linear correlation coefficient between the characteristic function and each preprocessed voxel time series was converted into z-values by Fisher’s z-transform [67]. This linear least-squares procedure models the case that the relationship between data and characteristic function is linear with unexplained variance expressed through normally distributed residuals.
- (2)
- Spatial interaction of BOLD amplitude mapping: For each voxel, the linear correlation coefficient between the characteristic function and the flux norm was computed and converted to z-values. A positive z-value means that during the task the absolute value of the flux as per Equation (1) increases. The procedure was repeated for the source estimate as per Equation (5). Again, any unexplained variance is modeled by normally distributed residuals.
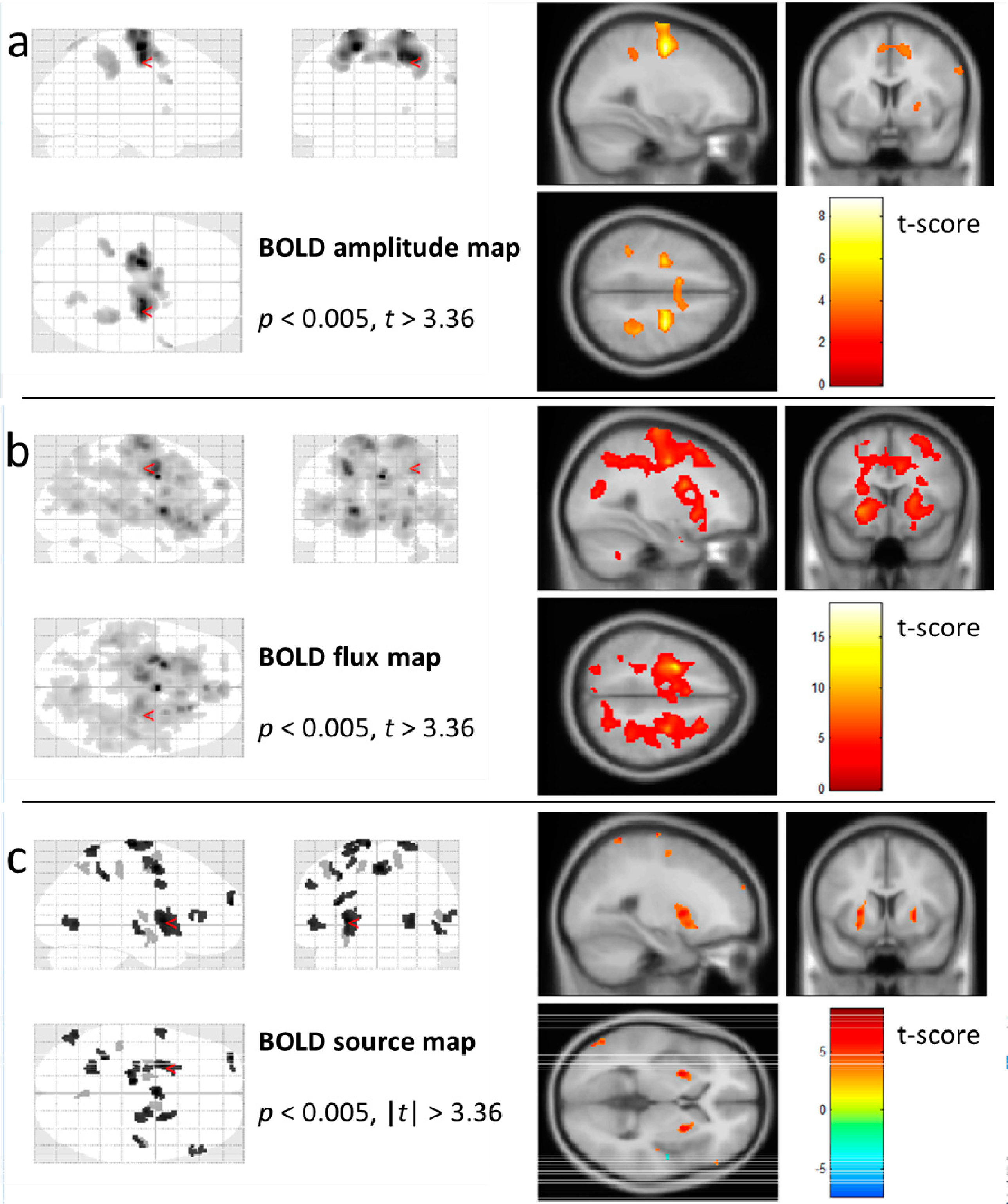
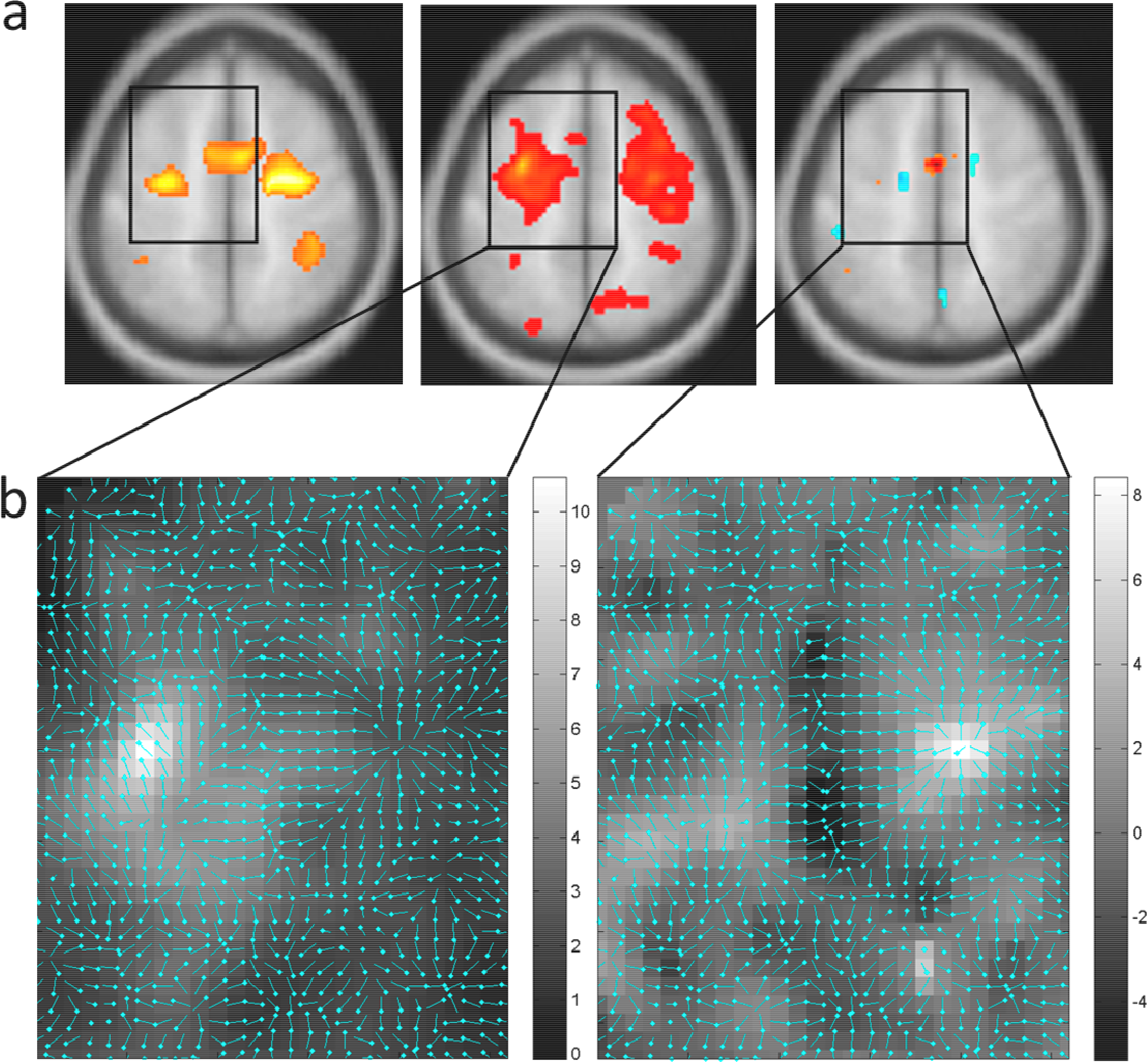
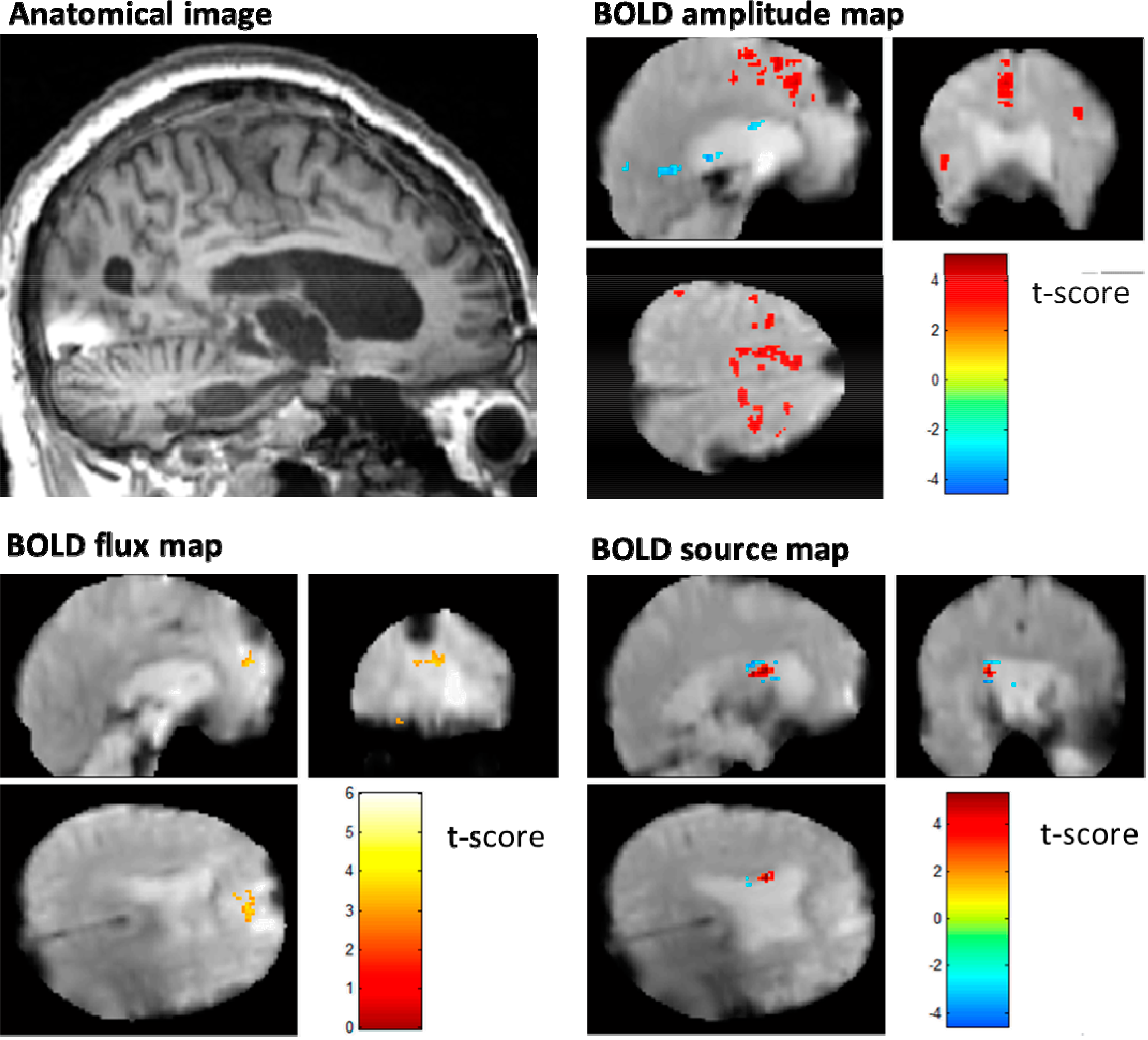
5. Results
5.1. BOLD Amplitude Mapping
5.2. BOLD Flux Mapping
5.3. BOLD Source Mapping
6. Discussion
6.1. Implications towards Conservation Laws Affecting Neuronal Dynamics
6.2. BOLD Flux Imaging and Model-Free Approaches
6.3. Putamen
6.4. Relation to Resting State fMRI
6.5. Differential Operators, Smoothing, and Nonlinear Transformations
6.6. Possible Clinical Applications
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Davis, T.L.; Kwong, K.K.; Weisskoff, R.M.; Rosen, B.R. Calibrated functional MRI: Mapping the dynamics of oxidative metabolism. Proc. Natl. Acad. Sci. USA 1998, 95, 1834–1839. [Google Scholar]
- Buxton, R.B.; Wong, E.C.; Frank, L.R. Dynamics of blood flow and oxygenation changes during brain activation: The balloon model. Magn. Reson. Med 1998, 39, 855–864. [Google Scholar]
- Buxton, R.B.; Uludag, K.; Dubowitz, D.J.; Liu, T.T. Modeling the hemodynamic response to brain activation. Neuroimage 2004, 23, S220–S233. [Google Scholar]
- Voss, H.U.; Ballon, D.J.; Domingos, A.I. Neuronal and hemodynamic source modeling of optogenetic BOLD signals. IEEE Signal Processing in Medicine and Biology Symposium, Brooklyn, New York, NY, USA, 10 December 2011; pp. 1–6.
- Voss, H.U.; Domingos, A.I. Analysis of coexisting neuronal populations in optogenetic and conventional BOLD data. IEEE Signal Processing in Medicine and Biology Symposium, Brooklyn, NY, USA, 1 December 2012; pp. 1–6.
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med 1995, 34, 537–541. [Google Scholar]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; van Essen, D.C.; Raichle, M.E. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci 2007, 8, 700–711. [Google Scholar]
- Friston, K.J.; Ashburner, J.T.; Kiebel, S.J.; Nichols, T.E.; Penny, W.D. Statistical Parametric Mapping: The Analysis of Functional Brain Images; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Sporns, O. Networks of the Brain; MIT Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Salvador, R.; Suckling, J.; Coleman, M.R.; Pickard, J.D.; Menon, D.; Bullmore, E. Neurophysiological architecture of functional magnetic resonance images of human brain. Cereb. Cortex 2005, 15, 1332–1342. [Google Scholar]
- Honey, C.J.; Kotter, R.; Breakspear, M.; Sporns, O. Network structure of cerebral cortex shapes functional connectivity on multiple time scales. Proc. Natl. Acad. Sci. USA 2007, 104, 10240–10245. [Google Scholar]
- Achard, S.; Salvador, R.; Whitcher, B.; Suckling, J.; Bullmore, E. A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. J. Neurosci 2006, 26, 63–72. [Google Scholar]
- Jeannerod, M.; Frak, V. Mental imaging of motor activity in humans. Curr. Opin. Neurobiol 1999, 9, 735–739. [Google Scholar]
- Cross, M.; Greenside, H. Pattern Formation and Dynamics in Nonequilibrium Systems; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Prigogine, I. Introduction to Thermodynamics of Irreversible Processes, 3d ed; Interscience Publishers: New York, NY, USA, 1968. [Google Scholar]
- Haddad, W.M. Temporal Asymmetry, Entropic Irreversibility, and Finite-Time Thermodynamics: From Parmenides-Einstein Time-Reversal Symmetry to the Heraclitan Entropic Arrow of Time. Entropy 2012, 14, 407–455. [Google Scholar]
- Tschoegl, N.W. Fundamentals of Equilibrium and Steady-State Thermodynamics; Elsevier Science: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Siegel, G.J. Basic Neurochemistry: Molecular, Cellular, and Medical Aspects, 7th ed; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Krieger, S.N.; Streicher, M.N.; Trampel, R.; Turner, R. Cerebral blood volume changes during brain activation. J. Cereb. Blood. Flow. Metab 2012, 32, 1618–1631. [Google Scholar]
- Yang, L.; Xenos, M.; Zhang, L.; Linninger, A.A. Prediction and Control of Blood Flow in the Human Brain Vascular Network. J. Undergrad. Res 2007, 1, 71–77. [Google Scholar]
- Zhang, D.; Raichle, M.E. Disease and the brain’s dark energy. Nat. Rev. Neurol 2010, 6, 15–28. [Google Scholar]
- Laughlin, S.B. Energy as a constraint on the coding and processing of sensory information. Curr. Opin. Neurobiol 2001, 11, 475–480. [Google Scholar]
- Shmuel, A.; Yacoub, E.; Pfeuffer, J.; van de Moortele, P.F.; Adriany, G.; Hu, X.; Ugurbil, K. Sustained negative BOLD, blood flow and oxygen consumption response and its coupling to the positive response in the human brain. Neuron 2002, 36, 1195–1210. [Google Scholar]
- Pasley, B.N.; Inglis, B.A.; Freeman, R.D. Analysis of oxygen metabolism implies a neural origin for the negative BOLD response in human visual cortex. Neuroimage 2007, 36, 269–276. [Google Scholar]
- Stefanovic, B.; Warnking, J.M.; Pike, G.B. Hemodynamic and metabolic responses to neuronal inhibition. Neuroimage 2004, 22, 771–778. [Google Scholar]
- Shmuel, A.; Augath, M.; Oeltermann, A.; Logothetis, N.K. Negative functional MRI response correlates with decreases in neuronal activity in monkey visual area V1. Nat. Neurosci 2006, 9, 569–577. [Google Scholar]
- Bianciardi, M.; Fukunaga, M.; van Gelderen, P.; de Zwart, J.A.; Duyn, J.H. Negative BOLD-fMRI signals in large cerebral veins. J. Cereb. Blood Flow Metab 2011, 31, 401–412. [Google Scholar]
- Kannurpatti, S.S.; Biswal, B.B. Negative functional response to sensory stimulation and its origins. J. Cereb. Blood Flow Metab 2004, 24, 703–712. [Google Scholar]
- Harel, N.; Lee, S.P.; Nagaoka, T.; Kim, D.S.; Kim, S.G. Origin of negative blood oxygenation level-dependent fMRI signals. J. Cereb. Blood Flow Metab 2002, 22, 908–917. [Google Scholar]
- Goense, J.; Merkle, H.; Logothetis, N.K. High-resolution fMRI reveals laminar differences in neurovascular coupling between positive and negative BOLD responses. Neuron 2012, 76, 629–639. [Google Scholar]
- Archer, J.S.; Abbott, D.F.; Waites, A.B.; Jackson, G.D. fMRI “deactivation” of the posterior cingulate during generalized spike and wave. Neuroimage 2003, 20, 1915–1922. [Google Scholar]
- Schridde, U.; Khubchandani, M.; Motelow, J.E.; Sanganahalli, B.G.; Hyder, F.; Blumenfeld, H. Negative BOLD with large increases in neuronal activity. Cereb. Cortex 2008, 18, 1814–1827. [Google Scholar]
- Sotero, R.C.; Trujillo-Barreto, N.J. Biophysical model for integrating neuronal activity, EEG, fMRI and metabolism. Neuroimage 2008, 39, 290–309. [Google Scholar]
- Shih, Y.Y.I.; Wey, H.Y.; De La Garza, B.H.; Duong, T.Q. Striatal and cortical BOLD, blood flow, blood volume, oxygen consumption, and glucose consumption changes in noxious forepaw electrical stimulation. J. Cereb. Blood Flow Metab 2011, 31, 832–841. [Google Scholar]
- Lee, J.H.; Durand, R.; Gradinaru, V.; Zhang, F.; Goshen, I.; Kim, D.S.; Fenno, L.E.; Ramakrishnan, C.; Deisseroth, K. Global and local fMRI signals driven by neurons defined optogenetically by type and wiring. Nature 2010, 465, 788–792. [Google Scholar]
- Domingos, A.I.; Vaynshteyn, J.; Voss, H.U.; Ren, X.; Gradinaru, V.; Zang, F.; Deisseroth, K.; de Araujo, I.E.; Friedman, J. Leptin regulates the reward value of nutrient. Nat. Neurosci 2011, 14, 1562–1568. [Google Scholar]
- Moraschi, M.; DiNuzzo, M.; Giove, F. On the origin of sustained negative BOLD response. J. Neurophysiol 2012, 108, 2339–2342. [Google Scholar]
- Lauritzen, M. Reading vascular changes in brain imaging: is dendritic calcium the key? Nat. Rev. Neurosci 2005, 6, 77–85. [Google Scholar]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar]
- Singh, K.D.; Fawcett, I.P. Transient and linearly graded deactivation of the human default-mode network by a visual detection task. Neuroimage 2008, 41, 100–112. [Google Scholar]
- Voss, H.U.; Heier, L.A.; Schiff, N.D. Multimodal imaging of recovery of functional networks associated with reversal of paradoxical herniation after cranioplasty. Clin. Imaging 2011, 35, 253–258. [Google Scholar]
- Mantini, D.; Perrucci, M.G.; Del Gratta, C.; Romani, G.L.; Corbetta, M. Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. USA 2007, 104, 13170–13175. [Google Scholar]
- Esposito, F.; Scarabino, T.; Hyvarinen, A.; Himberg, J.; Formisano, E.; Comani, S.; Tedeschi, G.; Goebel, R.; Seifritz, E.; Di Salle, F. Independent component analysis of fMRI group studies by self-organizing clustering. Neuroimage 2005, 25, 193–205. [Google Scholar]
- De Luca, M.; Beckmann, C.F.; De Stefano, N.; Matthews, P.M.; Smith, S.M. fMRI resting state networks define distinct modes of long-distance interactions in the human brain. Neuroimage 2006, 29, 1359–1367. [Google Scholar]
- Casanova, M.F.; El-Baz, A.; Switala, A. Laws of conservation as related to brain growth, aging, and evolution: symmetry of the minicolumn. Front. Neuroanat 2011, 5, 66. [Google Scholar]
- Charron, S.; Koechlin, E. Divided Representation of Concurrent Goals in the Human Frontal Lobes. Science 2010, 328, 360–363. [Google Scholar]
- Ophir, E.; Nass, C.; Wagner, A.D. Cognitive control in media multitaskers. Proc. Natl. Acad. Sci. USA 2009, 106, 15583–15587. [Google Scholar]
- Mandeville, J.B.; Marota, J.J.A.; Ayata, C.; Zaharchuk, G.; Moskowitz, M.A.; Rosen, B.R.; Weisskoff, R.M. Evidence of a cerebrovascular postarteriole windkessel with delayed compliance. J. Cereb. Blood Flow Metabol 1999, 19, 679–689. [Google Scholar]
- Friston, K.J.; Mechelli, A.; Turner, R.; Price, C.J. Nonlinear responses in fMRI: The balloon model, volterra kernels, and other hemodynamics. Neuroimage 2000, 12, 466–477. [Google Scholar]
- Friston, K.J.; Holmes, A.P.; Worsley, K.J.; Poline, J.P.; Frith, C.D.; Frackowiak, R.S.J. Statistical Parametric Maps in Functional Imaging: A General Linear Approach. Hum. Brain. Mapp 1995, 2, 189–210. [Google Scholar]
- Press, W.H. Numerical Recipes in C: The Art of Scientific Computing; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Guillot, A.; Collet, C. The Neurophysiological Foundations of Mental and Motor Imagery; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Boly, M.; Coleman, M.R.; Davis, M.H.; Hampshire, A.; Bor, D.; Moonen, G.; Maquet, P.A.; Pickard, J.D.; Laureys, S.; Owen, A.M. When thoughts become action: an fMRI paradigm to study volitional brain activity in non-communicative brain injured patients. Neuroimage 2007, 36, 979–992. [Google Scholar]
- Munzert, J.; Zentgraf, K.; Stark, R.; Vaitl, D. Neural activation in cognitive motor processes: comparing motor imagery and observation of gymnastic movements. Exp. Brain. Res 2008, 188, 437–444. [Google Scholar]
- Porro, C.A.; Francescato, M.P.; Cettolo, V.; Diamond, M.E.; Baraldi, P.; Zuiani, C.; Bazzocchi, M.; di Prampero, P.E. Primary motor and sensory cortex activation during motor performance and motor imagery: A functional magnetic resonance imaging study. J. Neurosci 1996, 16, 7688–7698. [Google Scholar]
- Ross, J.S.; Tkach, J.; Ruggieri, P.M.; Lieber, M.; Lapresto, E. The mind’s eye: Functional MR imaging evaluation of golf motor imagery. Am. J. Neuroradiol 2003, 24, 1036–1044. [Google Scholar]
- Roth, M.; Decety, J.; Raybaudi, M.; Massarelli, R.; Delon-Martin, C.; Segebarth, C.; Morand, S.; Gemignani, A.; Decorps, M.; Jeannerod, M. Possible involvement of primary motor cortex in mentally simulated movement: A functional magnetic resonance imaging study. Neuroreport 1996, 7, 1280–1284. [Google Scholar]
- Szameitat, A.J.; Shen, S.; Conforto, A.; Sterr, A. Cortical activation during executed, imagined, observed, and passive wrist movements in healthy volunteers and stroke patients. Neuroimage 2012, 62, 266–280. [Google Scholar] [Green Version]
- Szameitat, A.J.; Shen, S.; Sterr, A. Motor imagery of complex everyday movements. An fMRI study. Neuroimage 2007, 34, 702–713. [Google Scholar]
- Voss, H.U.; Helekar, S.A.; Schiff, N.D. Local spatial synchronization fMRI indicates functional specialization of putamen in motor imagery. Proc. Hum. Brain. Mapp 2014. submitted for publication. [Google Scholar]
- Yoo, S.S.; O’Leary, H.M.; Lee, J.H.; Chen, N.K.; Panych, L.P.; Jolesz, F.A. Reproducibility of trial-based functional MRI on motor imagery. Int. J. Neurosci 2007, 117, 215–227. [Google Scholar]
- Bardin, J.C.; Fins, J.J.; Katz, D.I.; Hersh, J.; Heier, L.A.; Tabelow, K.; Dyke, J.P.; Ballon, D.J.; Schiff, N.D.; Voss, H.U. Dissociations between behavioural and functional magnetic resonance imaging-based evaluations of cognitive function after brain injury. Brain 2011, 134, 769–782. [Google Scholar]
- Owen, A.M.; Coleman, M.R.; Boly, M.; Davis, M.H.; Laureys, S.; Pickard, J.D. Detecting awareness in the vegetative state. Science 2006, 313, 1402–1402. [Google Scholar]
- Owen, A.M.; Coleman, M.R. Functional MRI in disorders of consciousness: Advantages and limitations. Curr. Opin. Neurol 2007, 20, 632–637. [Google Scholar]
- Fisher, R.A. Frequency distribution of the values of the correlation coefficient in samples from an indefinitely large population. Biometrika 1914, 10, 507–521. [Google Scholar]
- Huettel, S.A.; Song, A.W.; McCarthy, G. Functional Magnetic Resonance Imaging, 2nd ed; Sinauer Associates: Sunderland, UK, 2008. [Google Scholar]
- Friston, K.J.; Stephan, K.E.; Lund, T.E.; Morcom, A.; Kiebel, S. Mixed-effects and fMRI studies. Neuroimage 2005, 24, 244–252. [Google Scholar]
- Monti, M.M.; Vanhaudenhuyse, A.; Coleman, M.R.; Boly, M.; Pickard, J.D.; Tshibanda, L.; Owen, A.M.; Laureys, S. Willful modulation of brain activity in disorders of consciousness. New Engl. J. Med 2010, 362, 579–589. [Google Scholar]
- Harrison, L.M.; Penny, W.; Daunizeau, J.; Friston, K.J. Diffusion-based spatial priors for functional magnetic resonance images. Neuroimage 2008, 41, 408–423. [Google Scholar]
- Tabelow, K.; Polzehl, J.; Voss, H.U.; Spokoiny, V. Analyzing fMRI experiments with structural adaptive smoothing procedures. Neuroimage 2006, 33, 55–62. [Google Scholar]
- Galizia, C.G.; Lledo, P.-M. Neurosciences: From Molecule to Behavior: A University Textbook; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Rosenberg, G.A. Molecular Physiology Metabolism of the Nervous System: A Clinical Perspective; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Nicholls, J.G. From Neuron to Brain, 5th ed; Sinauer Associates: Sunderland, UK, 2012. [Google Scholar]
- Gonzalez-Castillo, J.; Saad, Z.S.; Handwerker, D.A.; Inati, S.J.; Brenowitz, N.; Bandettini, P.A. Whole-brain, time-locked activation with simple tasks revealed using massive averaging and model-free analysis. Proc. Natl. Acad. Sci. USA 2012, 109, 5487–5492. [Google Scholar]
- Adler, A.; Finkes, I.; Katabi, S.; Prut, Y.; Bergman, H. Encoding by Synchronization in the Primate Striatum. J. Neurosci 2013, 33, 4854–4866. [Google Scholar]
- Goldman, R.I.; Stern, J.M.; Engel, J.; Cohen, M.S. Simultaneous EEG and fMRI of the alpha rhythm. Neuroreport 2002, 13, 2487–2492. [Google Scholar]
- Voss, H.U.; Schiff, N.D. MRI of neuronal network structure, function, and plasticity. Prog. Brain Res 2009, 175, 483–496. [Google Scholar]
- Park, H.J.; Friston, K. Structural and Functional Brain Networks: From Connections to Cognition. Science 2013, 342, 579–587. [Google Scholar]
- Romeney, B.M.T.H. Front-End Vision Multi-Scale Image Analysis: Multi-Scale Computer Vision Theory and Applications, Written in Mathematica; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Voss, H.U.; Kolodner, P.; Abel, M.; Kurths, J. Amplitude equations from spatiotemporal binary-fluid convection data. Phys. Rev. Lett 1999, 83, 3422–3425. [Google Scholar]
- Giacino, J.T.; Ashwal, S.; Childs, N.; Cranford, R.; Jennett, B.; Katz, D.I.; Kelly, J.P.; Rosenberg, J.H.; Whyte, J.; Zafonte, R.D.; et al. The minimally conscious state—Definition and diagnostic criteria. Neurology 2002, 58, 349–353. [Google Scholar]
- Posner, J.B.; Saper, C.B.; Schiff, N.D.; Plum, F. Plum and Posner’s Diagnosis of Stupor and Coma; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Stender, J.; Gosseries, O.; Bruno, M.-A.; Vanessa, C.-V.; Vanhaudenhuyse, A.; Demertzi, A.; Chatelle, C.; Thonnard, M.; Thibaut, A.; et al. Diagnostic precision of PET imaging and functional MRI in disorders of consciousness: A clinical validation study. Lancet 2014. [Google Scholar] [CrossRef]
- Plum, F.; Posner, J. The Locked in Syndrome. Br. Med. J 1987, 294, 1163–1163. [Google Scholar]
- Schiff, N.D.; Plum, F.; Rezai, A.R. Developing prosthetics to treat cognitive disabilities resulting from acquired brain injuries. Neurolog. Res 2002, 24, 116–124. [Google Scholar]
- Schiff, S.J. Neural Control: The Emerging Intersection between Control Theory and Neuroscience; MIT Press: Cambridge, MA, 2011. [Google Scholar]
- Schiff, N.D.; Rezai, A.R.; Plum, F. A neuromodulation strategy for rational therapy of complex brain injury states. Neurol. Res 2000, 22, 267–272. [Google Scholar]
- Schiff, N.D.; Giacino, J.T.; Kalmar, K.; Victor, J.D.; Baker, K.; Gerber, M.; Fritz, B.; Eisenberg, B.; O’Connor, J.; Kobylarz, E.J.; et al. Behavioural improvements with thalamic stimulation after severe traumatic brain injury. Nature 2007, 448, 600–603. [Google Scholar]
- Sauer, T.D.; Schiff, S.J. Data assimilation for heterogeneous networks: The consensus set. Phys. Rev. E 2009, 79, 051909. [Google Scholar]
- Schiff, N.D. Measurements and models of cerebral function in the severely injured brain. J. Neurotrauma 2006, 23, 1436–1449. [Google Scholar]
- Schiff, N.D. Modeling the minimally conscious state: Measurements of brain function and therapeutic possibilities. Bound. Conscious.: Neurobiol. Neuropathol 2005, 150, 473–493. [Google Scholar]
- Coleman, M.R.; Menon, D.K.; Fryer, T.D.; Pickard, J.D. Neurometabolic coupling in the vegetative and minimally conscious states: Preliminary findings. J. Neurol. Neurosurg. Psychiatry 2005, 76, 432–434. [Google Scholar]



© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Voss, H.U.; Schiff, N.D. Searching for Conservation Laws in Brain Dynamics—BOLD Flux and Source Imaging. Entropy 2014, 16, 3689-3709. https://doi.org/10.3390/e16073689
Voss HU, Schiff ND. Searching for Conservation Laws in Brain Dynamics—BOLD Flux and Source Imaging. Entropy. 2014; 16(7):3689-3709. https://doi.org/10.3390/e16073689
Chicago/Turabian StyleVoss, Henning U., and Nicholas D. Schiff. 2014. "Searching for Conservation Laws in Brain Dynamics—BOLD Flux and Source Imaging" Entropy 16, no. 7: 3689-3709. https://doi.org/10.3390/e16073689
APA StyleVoss, H. U., & Schiff, N. D. (2014). Searching for Conservation Laws in Brain Dynamics—BOLD Flux and Source Imaging. Entropy, 16(7), 3689-3709. https://doi.org/10.3390/e16073689