
Single to Two Cluster State Transition of Primary Motor Cortex 4-posterior (MI-4p) Activities in Humans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction


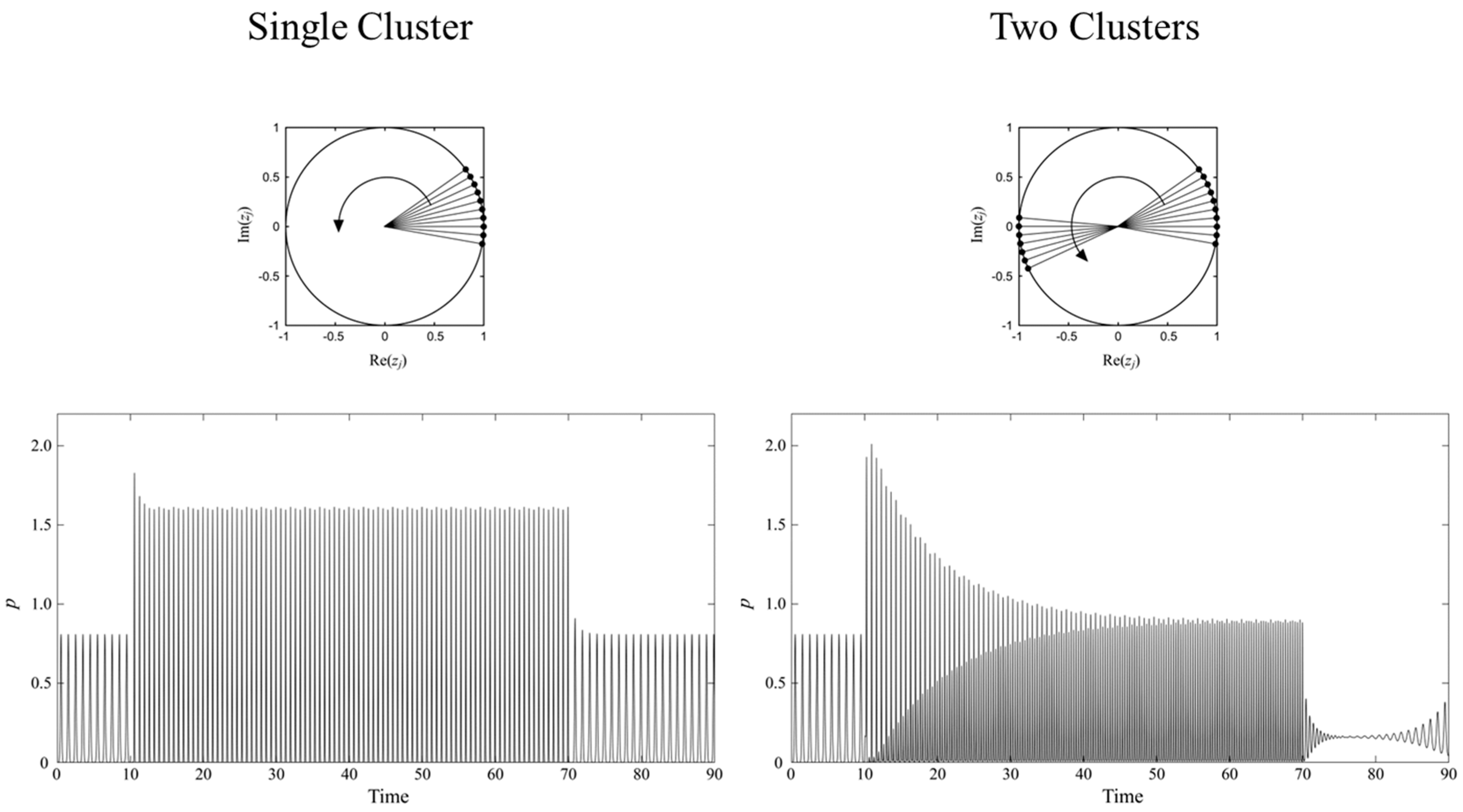
2. Reproduction of the Tass Entrainment Model
3. Functional MRI
3.1. Subjects
3.2. Data Acquisition and Processing
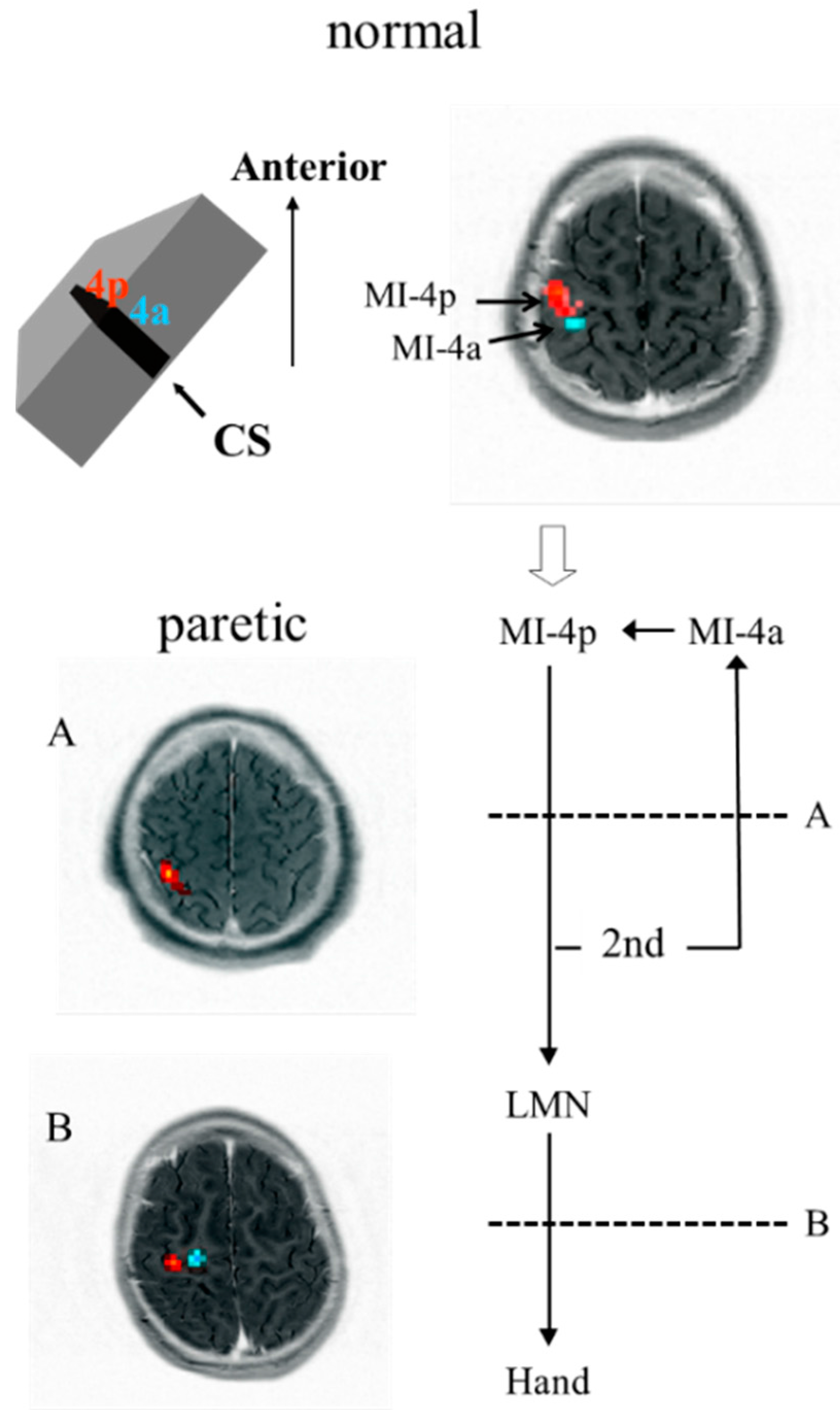
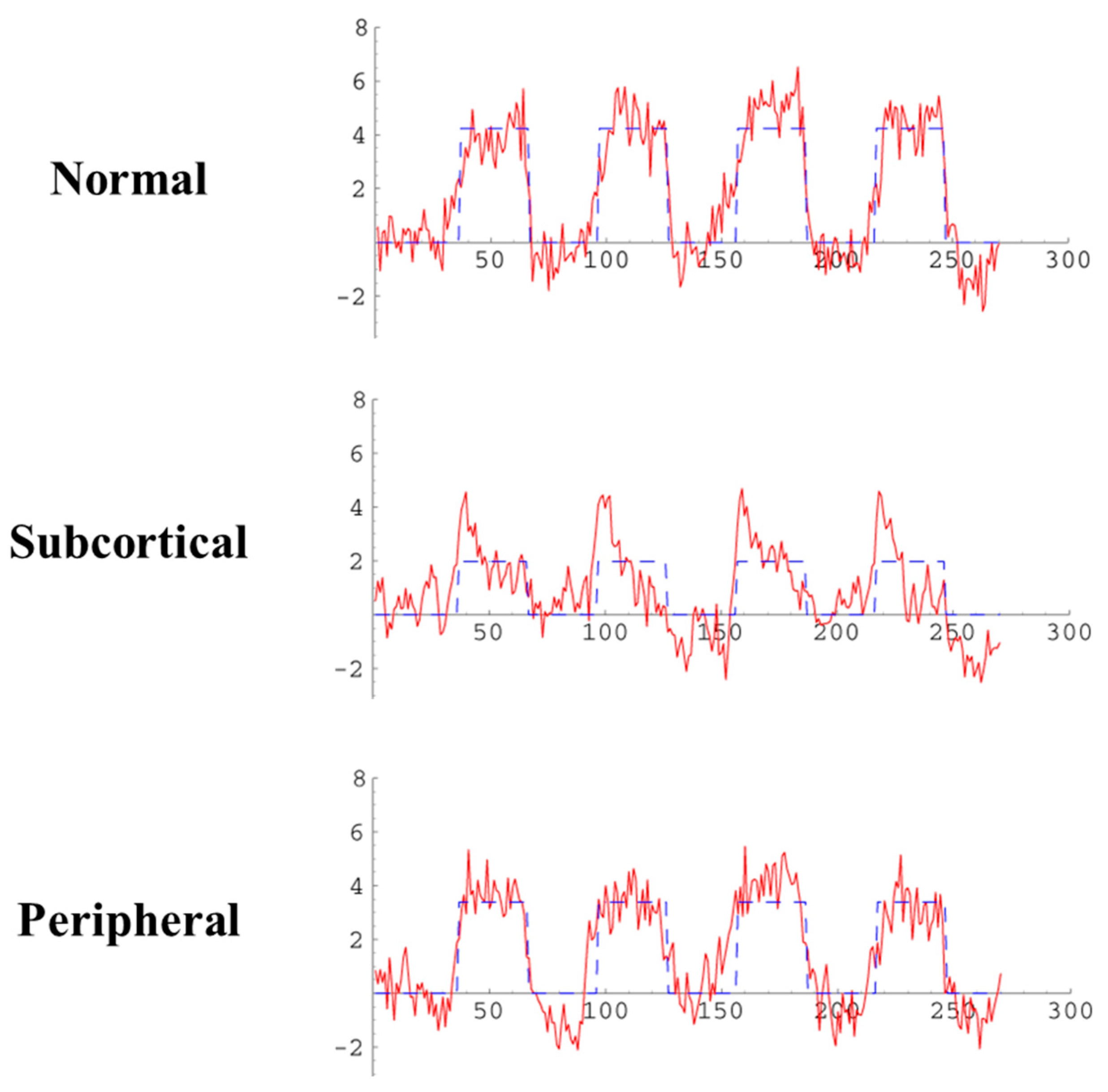
3.3. Results


4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
- Q = 0.4
- K1 = 1, K2 = K3 = K4 = 0
- C1 = C2 = C3 = C4 = 0
- ω = 3π
- I1 = 4, I2= I3 = I4 = 0 for single cluster
- I2 = 4, I1 = I3 = I4 = 0 for two clusters
- γ1 = γ2 = γ3 = γ4 = 0
References
- Strick, P.L.; Preston, B. Two representations of the hand in area 4 of a primate. I. Motor output organization. J. Neurophysiol. 1982, 48, 139–149. [Google Scholar] [PubMed]
- Strick, P.L.; Preston, B. Two representations of the hand in area 4 of a primate. II. Somatosensory input organization. J. Neurophysiol. 1982, 48, 150–159. [Google Scholar] [PubMed]
- Geyer, S.; Ledberg, A.; Schleicher, A.; Kinomura, S.; Schormann, T.; Bürgel, U.; Klingberg, T.; Larsson, J.; Zilles, K.; Roland, P.E. Two different areas within the primary motor cortex of man. Nature 1996, 382, 805–807. [Google Scholar] [CrossRef] [PubMed]
- Nakada, T.; Fujii, Y.; Suzuki, K.; Kwee, I.L. High-field (3.0T) functional MRI sequential epoch analysis: An example for motion control analysis. Neurosci. Res. 1998, 32, 355–362. [Google Scholar] [CrossRef]
- Nakada, T.; Suzuki, K.; Fujii, Y.; Matsuzawa, H.; Kwee, I.L. Independent component-cross correlation-sequential epoch (ICS) analysis of high-field fMRI time series: direct visualization of dual representation of the primary motor cortex in human. Neurosci. Res. 2000, 37, 237–244. [Google Scholar] [CrossRef]
- Haken, H. Principles of Brain Functioning; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Benabid, A.L.; Chabardes, S.; Mitrofanis, J.; Pollak, P. Deep brain stimulation of the subthalamic nucleus for the treatment of Parkinson’s disease. Lancet Neurol. 2009, 8, 67–81. [Google Scholar] [CrossRef]
- Perlmutter, J.S.; Mink, J.W. Deep brain stimulation. Ann. Rev. Neurosci. 2006, 29, 229–257. [Google Scholar] [CrossRef] [PubMed]
- Tass, P.A. Phase Resetting in Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Nakada, T. Myths and truths in functional MRI: A basic guide for practitioners. Magn. Reson. Med. Sci. 2002, 1, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Brunnstrom, S. Motor testing procedures in hemiplegia: Based on sequential recovery stages. Phys. Ther. 1966, 46, 357–375. [Google Scholar] [PubMed]
- McKeown, M.J.; Makeig, S.; Brown, G.G.; Jung, T.-P.; Kindermann, S.S.; Kindermann, R.S.; Bell, A.J.; Sejnowski, T.J. Analysis of fMRI data by blind separation into independent spatial components. Hum. Brain Map. 1995, 6, 160–188. [Google Scholar] [CrossRef]
- Comon, P. Independent component analysis: A new concept? Signal Process. 1994, 36, 287–314. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.J.; Sejnowski, T.J. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995, 7, 1129–1159. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S.; Jung, T.P.; Ghahremani, D.; Bell, B.J.; Sejnowski, T.J. Blind separation of auditory event-related brain responses into independent components. Proc. Natl. Acad. Sci. USA 1997, 94, 10979–10984. [Google Scholar] [CrossRef] [PubMed]
- Amari, S. National gradient works efficiently in learning. Neural Comput. 1998, 10, 251–276. [Google Scholar] [CrossRef]
- Singer, W.; Gray, C.M. Visual feature integration and the temporal correlation hypothesis. Annu. Rev. Neurosci. 1995, 18, 555–586. [Google Scholar] [CrossRef] [PubMed]
- Winfree, A.T. The Geometry of Biological Time; Springer: Berlin/Heidelberg, Germany, 1980. [Google Scholar]
- Nakada, T.; Kwee, I.L. Oculopalatal myoclonus. Brain 1986, 109, 431–441. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakada, K.; Suzuki, K.; Nakada, T. Single to Two Cluster State Transition of Primary Motor Cortex 4-posterior (MI-4p) Activities in Humans. Entropy 2015, 17, 7596-7607. https://doi.org/10.3390/e17117596
Nakada K, Suzuki K, Nakada T. Single to Two Cluster State Transition of Primary Motor Cortex 4-posterior (MI-4p) Activities in Humans. Entropy. 2015; 17(11):7596-7607. https://doi.org/10.3390/e17117596
Chicago/Turabian StyleNakada, Kazunori, Kiyotaka Suzuki, and Tsutomu Nakada. 2015. "Single to Two Cluster State Transition of Primary Motor Cortex 4-posterior (MI-4p) Activities in Humans" Entropy 17, no. 11: 7596-7607. https://doi.org/10.3390/e17117596
APA StyleNakada, K., Suzuki, K., & Nakada, T. (2015). Single to Two Cluster State Transition of Primary Motor Cortex 4-posterior (MI-4p) Activities in Humans. Entropy, 17(11), 7596-7607. https://doi.org/10.3390/e17117596