
Self-Organized Patterns Induced by Neimark-Sacker, Flip and Turing Bifurcations in a Discrete Predator-Prey Model with Lesie-Gower Functional Response



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Model and Stability Analysis
2.1. A Discrete Predator-Prey Model
2.2. Fixed Points and Stability
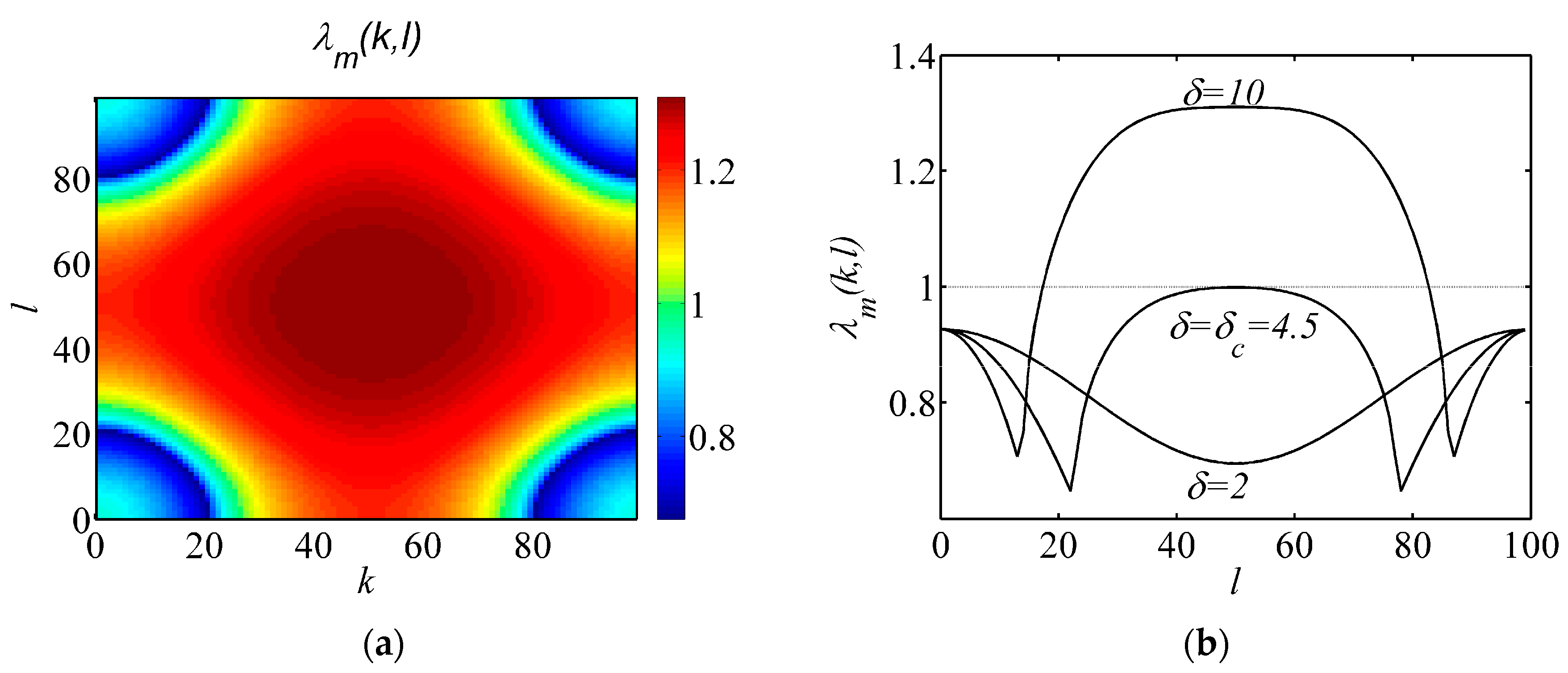
2.3. Bifurcation Analysis
3. Numerical Simulations
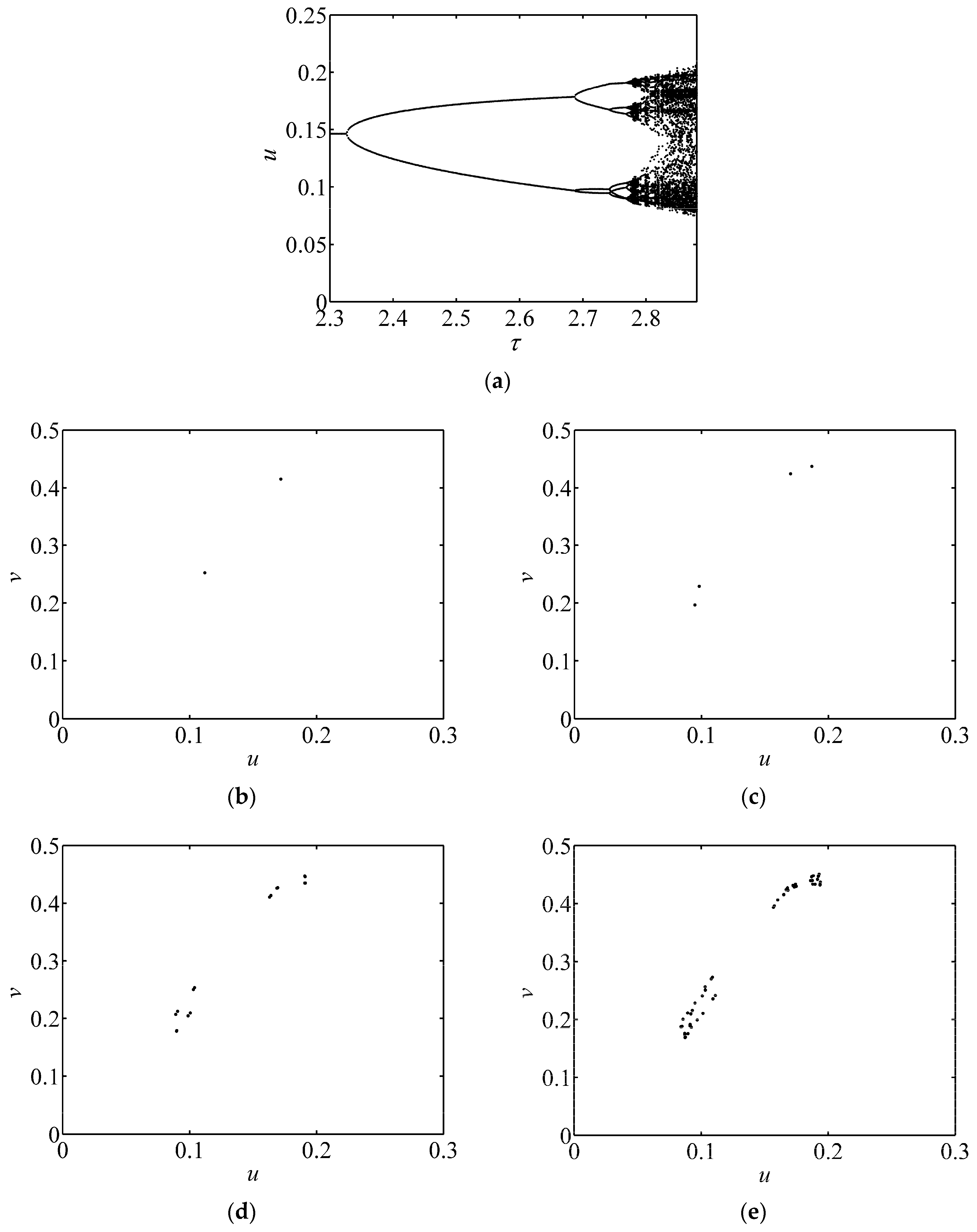
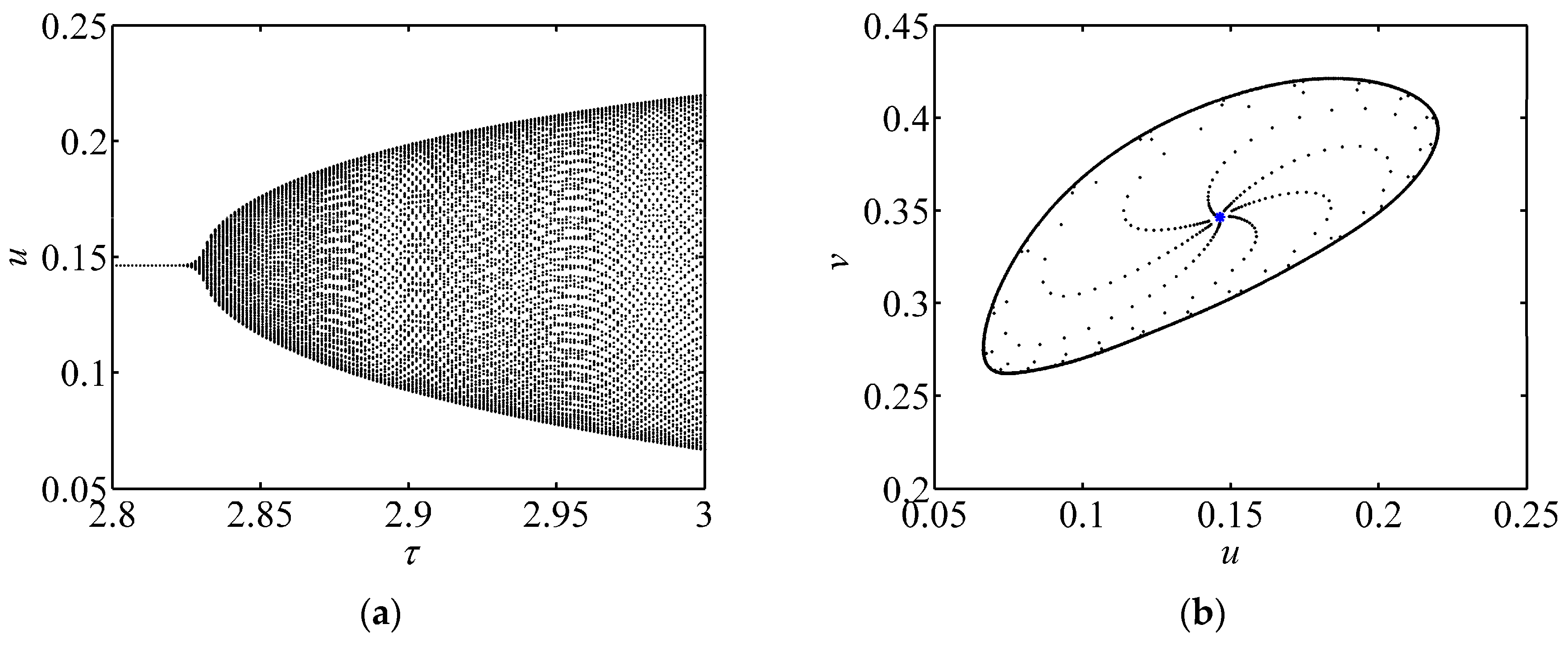
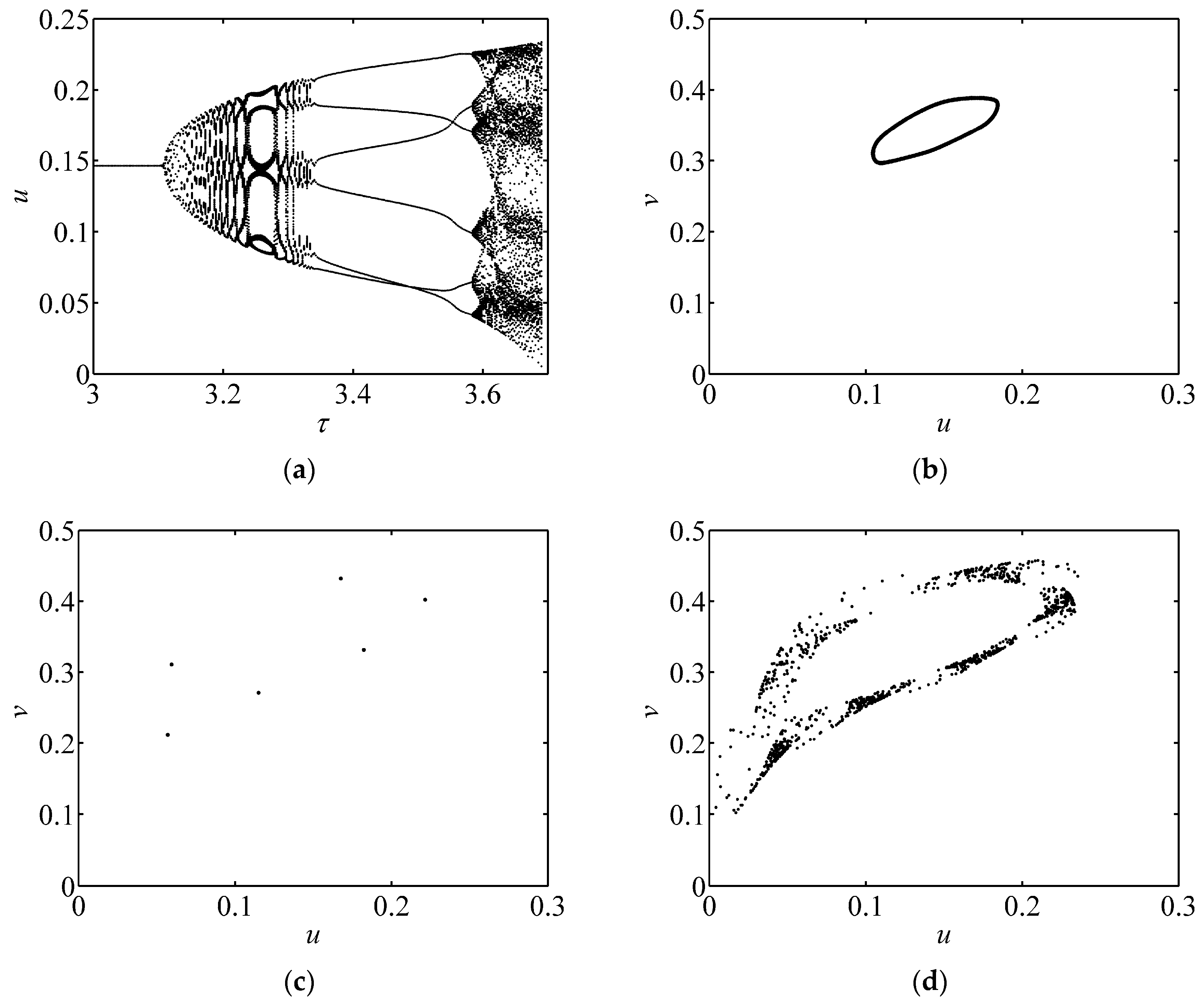
3.1. Bifurcation Diagram and Phase Portrait
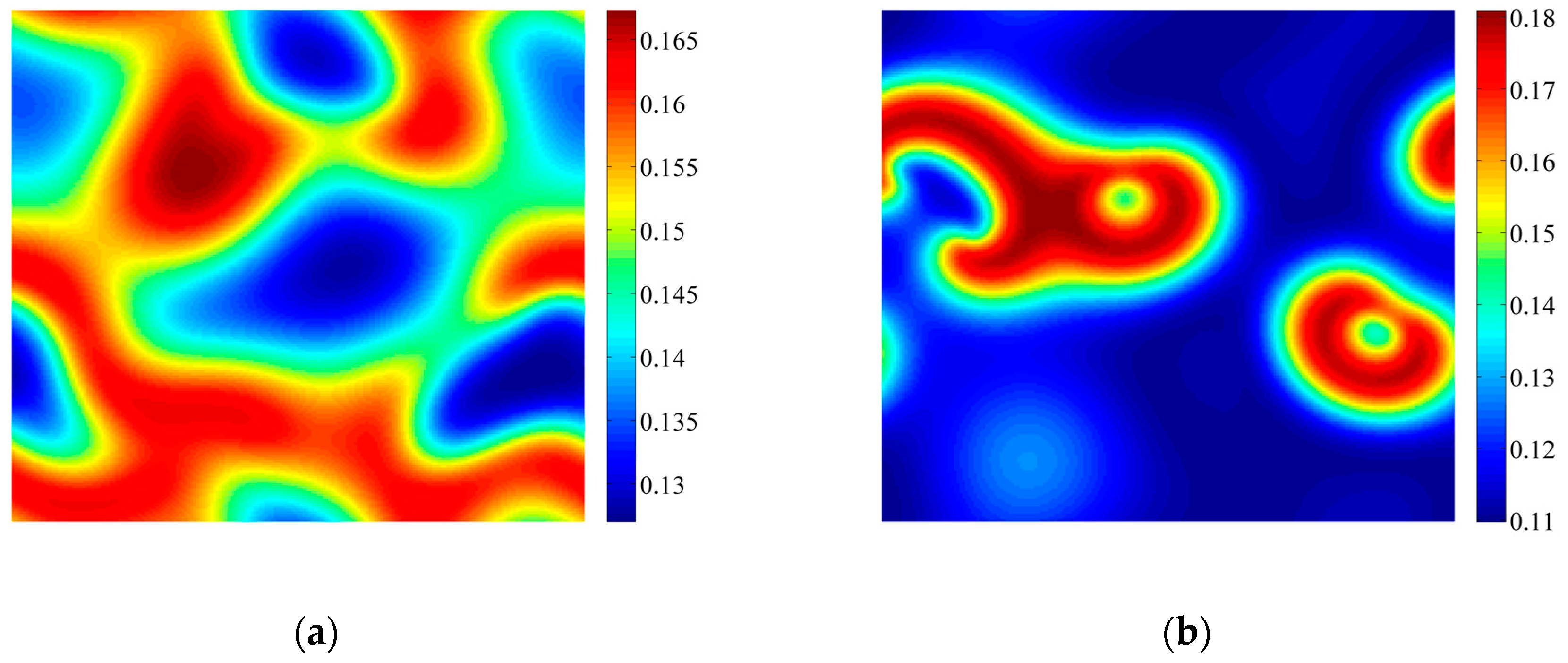
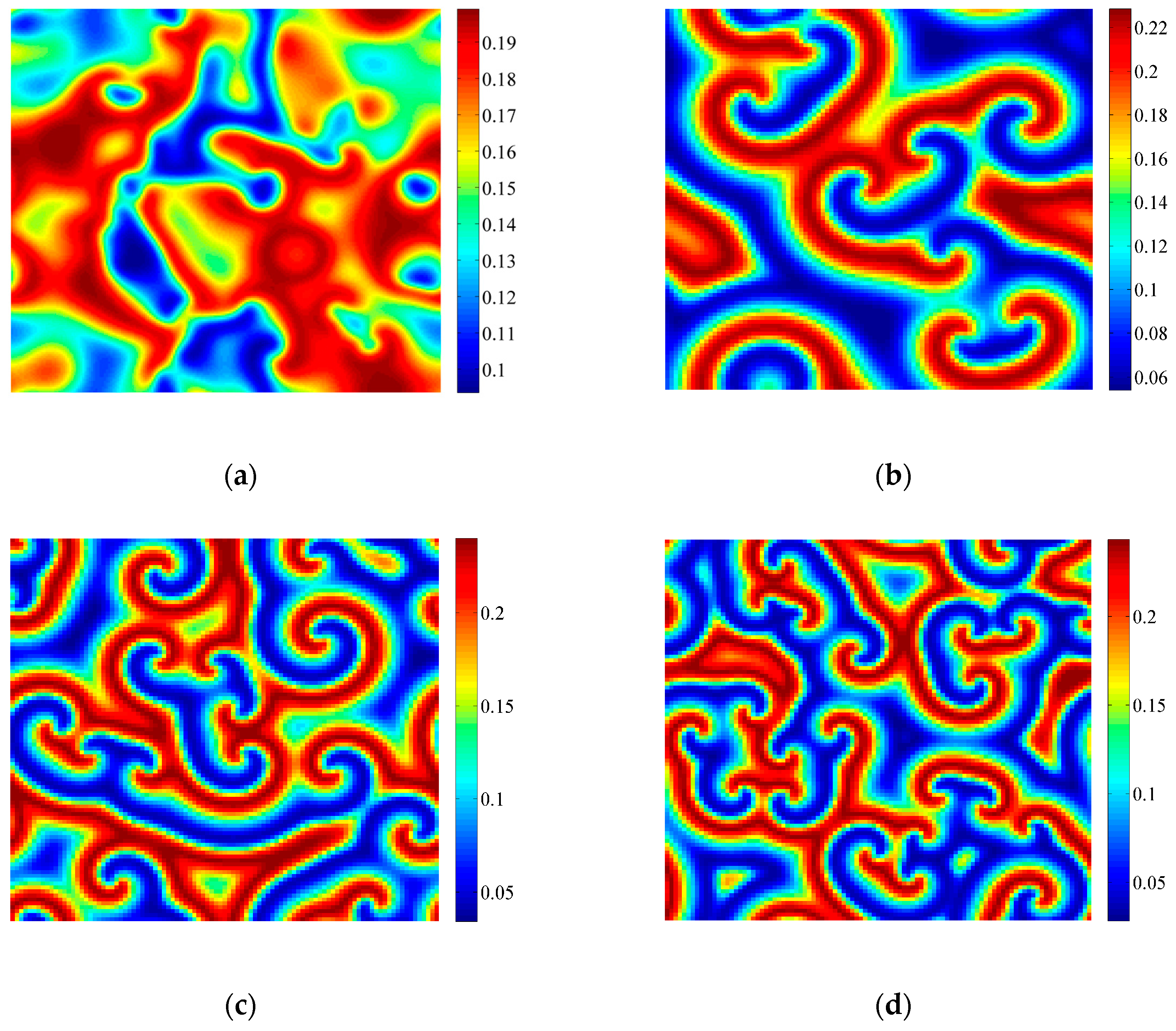
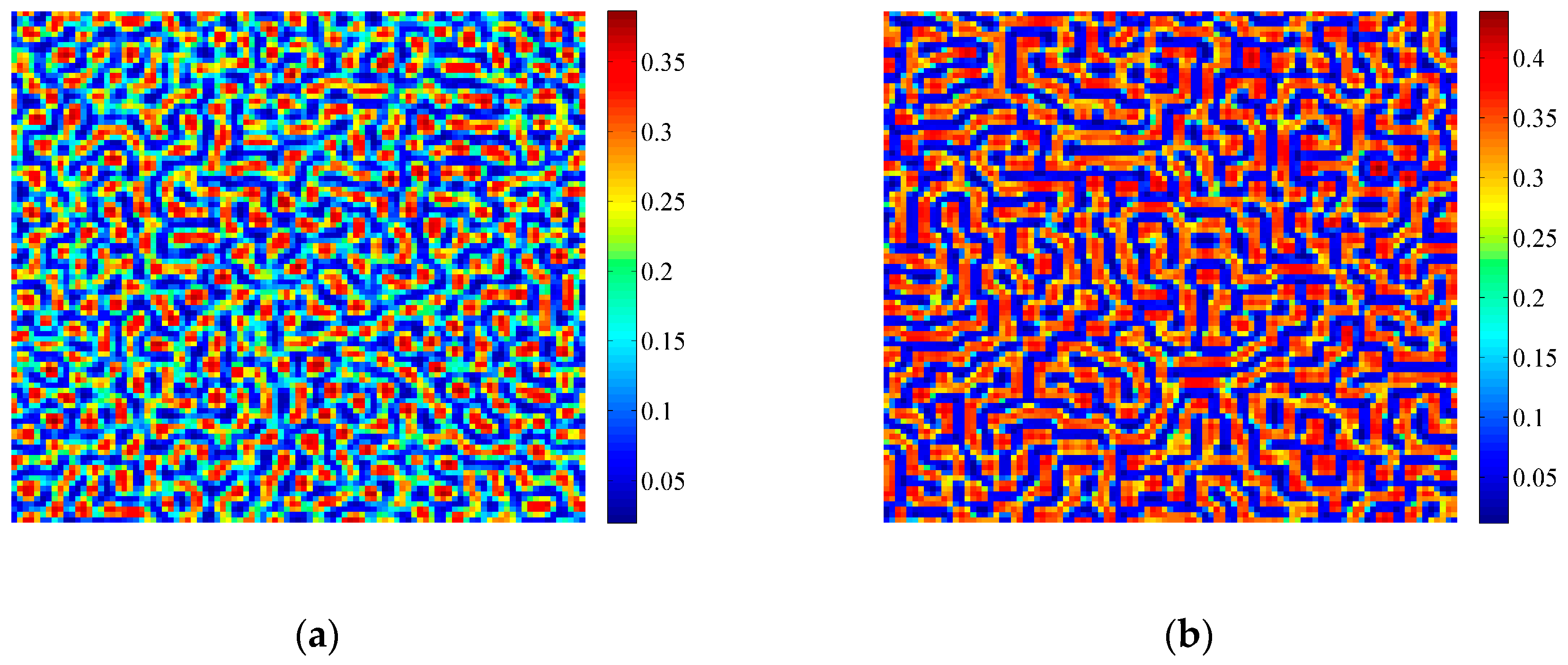
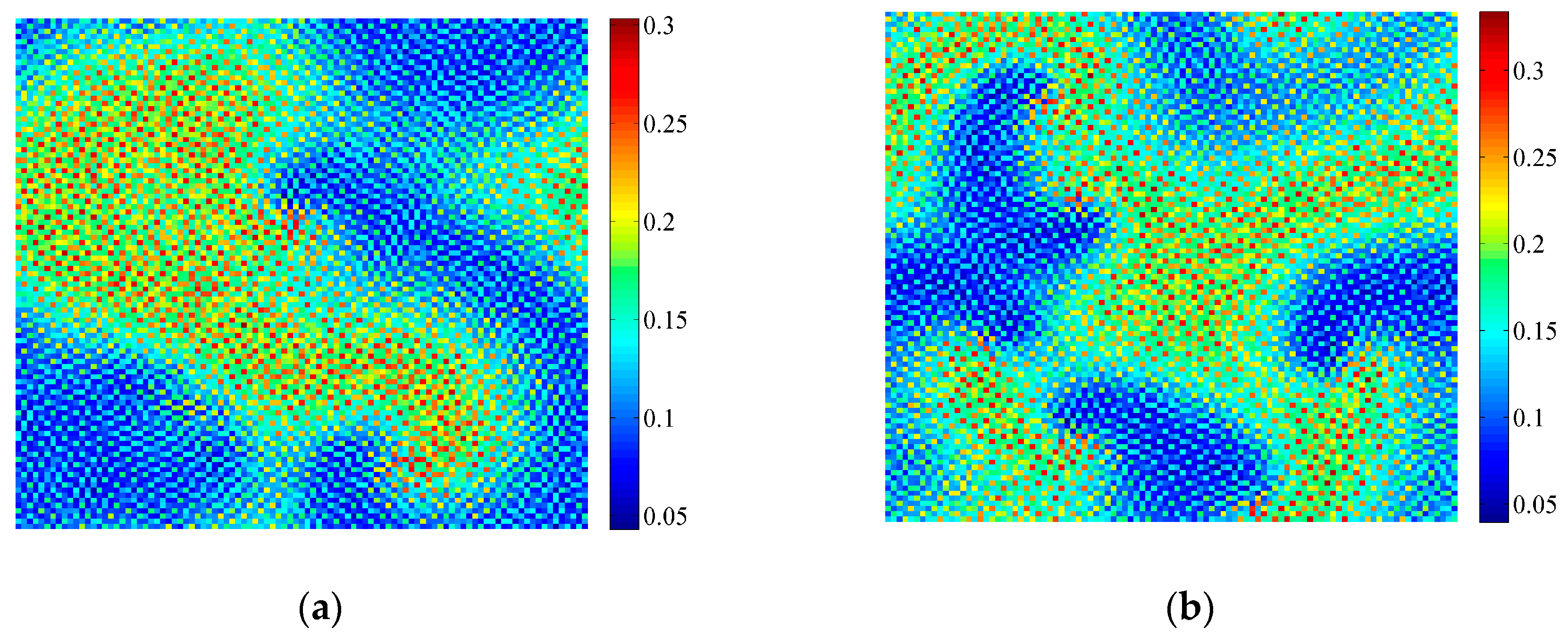
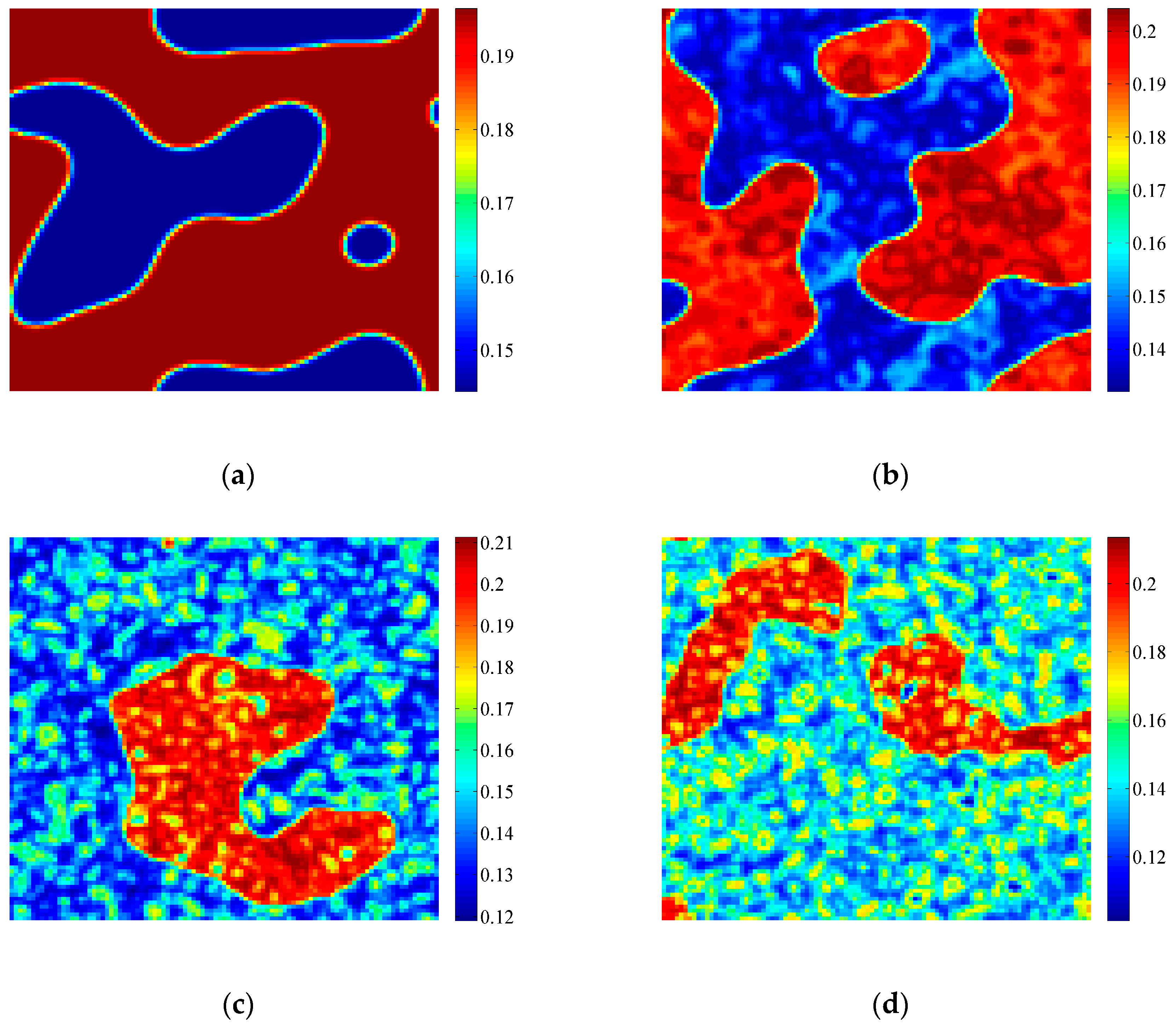
3.2. Formation of Self-Organized Patterns
4. Discussion and Conclusions
- The discrete predator-prey model with Lesie-Gower functional response can generate many complex dynamics including three types of bifurcations, which are flip bifurcation, Neimark-Sacker bifurcation and Turing bifurcation.
- A variety of self-organized patterns can be formed through the discrete predator-prey model with Lesie-Gower functional response and the above three bifurcations. These patterns consist of spots, transitional patterns from spots to spirals, spirals, spirals coupled with mosaics, labyrinths, and many other complex patterns generated by flip bifurcation.
- Among the studies on predator-prey models with Lesie-Gower functional response, this research may develop a special perspective to interpret the how self-organized patterns are generated.
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
Appendix A.1. Flip Bifurcation Analysis
Appendix A.2. Neimark-Sacker Bifurcation Analysis
Appendix A.3. Turing Bifurcation Analysis
References
- Taylor, A.D. Metapopulations, dispersal, and predator-prey dynamics: An overview. Ecology 1990, 71, 429–433. [Google Scholar] [CrossRef]
- Briggs, C.J.; Hoopes, M.F. Stabilizing effects in spatial parasitoid-host and predator-prey models: A review. Theor. Popul. Boil. 2004, 65, 299–315. [Google Scholar] [CrossRef]
- Hu, D.; Cao, H. Bifurcation and chaos in a discrete-time predator-prey system of Holling and Leslie type. Commun. Nonlinear Sci. 2015, 22, 702–715. [Google Scholar] [CrossRef]
- Yang, R. Hopf bifurcation analysis of a delayed diffusive predator-prey system with non-constant death rate. Chaos Solitons Fract. 2015, 81, 224–232. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, T.; Dai, L. Nonlinear dynamic analysis and characteristics diagnosis of seasonally perturbed predator-prey systems. Commun. Nonlinear Sci. 2015, 22, 407–419. [Google Scholar] [CrossRef]
- Kondo, S.; Miura, T. Reaction-diffusion model as a framework for understanding biological pattern formation. Science 2010, 329, 1616–1620. [Google Scholar] [CrossRef] [PubMed]
- Rao, F.; Wang, W.; Li, Z. Spatiotemporal complexity of a predator-prey system with the effect of noise and external forcing. Chaos Solitons Fract. 2009, 41, 1634–1644. [Google Scholar] [CrossRef]
- Guin, L.N. Spatial patterns through Turing instability in a reaction-diffusion predator-prey model. Math. Comput. Simul. 2015, 109, 174–185. [Google Scholar] [CrossRef]
- Cobbold, C.A.; Lutscher, F.; Sherratt, J.A. Diffusion-driven instabilities and emerging spatial patterns in patchy landscapes. Ecol. Complex. 2015, 24, 69–81. [Google Scholar] [CrossRef]
- Cai, Y.; Zhao, C.; Wang, W. Spatiotemporal complexity of a Leslie-Gower predator-prey model with the weak Alee effect. J. Appl. Math. 2013, 2013, 535746. [Google Scholar] [CrossRef]
- Dilão, R. Turing instabilities and patterns near a Hopf bifurcation. Appl. Math. Comput. 2005, 64, 391–414. [Google Scholar] [CrossRef]
- Chang, L.; Sun, G.Q.; Wang, Z.; Jin, Z. Rich dynamics in a spatial predator-prey model with delay. Appl. Math. Comput. 2015, 256, 540–550. [Google Scholar] [CrossRef]
- Abid, W.; Yafia, R.; Aziz-Alaoui, M.A.; Bouhafa, H.; Abichou, A. Diffusion driven instability and Hopf bifurcation in spatial predator-prey model on a circular domain. Appl. Math. Comput. 2015, 260, 292–313. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, L.; Xue, Y.; Jin, Z. Spatiotemporal pattern formation of Beddington-DeAngelis-type predator-prey model. arXiv, 2008; arXiv:0801.0797. [Google Scholar]
- Klausmeier, C.A. Regular and irregular patterns in semiarid vegetation. Science 1999, 284, 1826–1828. [Google Scholar] [CrossRef] [PubMed]
- Meron, E.; Gilad, E.; von Hardenberg, J.; Shachak, M.; Zarmi, Y. Vegetation patterns along a rainfall gradient. Chaos Solitons Fract. 2004, 19, 367–376. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, X.; Huang, X. Hopf bifurcation and spatial patterns of a delayed biological economic system with diffusion. Appl. Math. Comput. 2015, 266, 462–480. [Google Scholar] [CrossRef]
- Wang, Q.; Fan, M.; Wang, K. Dynamics of a class of nonautonomous semi-ratio-dependent predator-prey systems with functional responses. J. Math. Anal. Appl. 2003, 278, 443–471. [Google Scholar] [CrossRef]
- Leslie, P.H. Some further notes on the use of matrices in population mathematics. Biometrika 1948, 35, 213–245. [Google Scholar] [CrossRef]
- Leslie, P.H.; Gower, J.C. The properties of a stochastic model for the predator-prey type of interaction between two species. Biometrika 1960, 47, 219–234. [Google Scholar] [CrossRef]
- Wang, W.; Lin, Y.; Zhang, L.; Rao, F.; Tan, Y. Complex patterns in a predator-prey model with self and cross-diffusion. Commun. Nonlinear Sci. 2011, 16, 2006–2015. [Google Scholar] [CrossRef]
- Zhang, X.C.; Sun, G.Q.; Jin, Z. Spatial dynamics in a predator-prey model with Beddington-DeAngelis functional response. Phys. Rev. E 2012, 85, 021924. [Google Scholar] [CrossRef] [PubMed]
- Xue, L. Pattern formation in a predator-prey model with spatial effect. Phys. A Stat. Mech. Appl. 2012, 391, 5987–5996. [Google Scholar] [CrossRef]
- Haque, M. Existence of complex patterns in the Beddington-DeAngelis predator-prey model. Math. Biosci. 2012, 239, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Inter-Science: New York, NY, USA, 1969. [Google Scholar]
- Mistro, D.C.; Rodrigues, L.A.D.; Petrovskii, S. Spatiotemporal complexity of biological invasion in a space-and time-discrete predator-prey system with the strong Allee effect. Ecol. Complex. 2012, 9, 16–32. [Google Scholar] [CrossRef]
- Han, Y.T.; Han, B.; Zhang, L.; Xu, L.; Li, M.F.; Zhang, G. Turing instability and wave patterns for a symmetric discrete competitive Lotka-Volterra system. WSEAS Trans. Math. 2011, 10, 181–189. [Google Scholar]
- Aziz-Alaoui, M.A.; Daher-Okiye, M. Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling type II schemes. Appl. Math. Lett. 2003, 16, 1069–1075. [Google Scholar] [CrossRef]
- Daher-Okiye, M.; Aziz-Alaoui, M.A. On the dynamics of a predator-prey model with the Holling-Tanner functional response. In Mathematical Modelling & Computing in Biology and Medicine: 5th ESMTB Conference 2002; Capasso, V., Ed.; Società Editrice Esculapio: Bologna, Italy, 2002; pp. 270–278. [Google Scholar]
- Korobeinikov, A. A Lyapunov function for Leslie-Gower predator-prey models. Appl. Math. Lett. 2001, 14, 697–699. [Google Scholar] [CrossRef]
- Nindjin, A.F.; Aziz-Alaoui, M.A.; Cadivel, M. Analysis of a predator-prey model with modified Leslie-Gower and Holling-type II schemes with time delay. Nonlinear Anal. Real. 2006, 7, 1104–1118. [Google Scholar] [CrossRef]
- Nindjin, A.F.; Aziz-Alaoui, M.A. Persistence and global stability in a delayed Leslie-Gower type three species food chain. J. Math. Anal. Appl. 2008, 340, 340–357. [Google Scholar] [CrossRef]
- Aziz-Alsoui, M.A. Study of a Leslie Gower-type tritrophic population. Chaos Solitons Fract. 2002, 14, 1275–1293. [Google Scholar] [CrossRef]
- Letellier, C.; Asis-Alaoui, M.A. Analysis of the dynamics of a realistic ecological model. Chaos Solitons Fract. 2002, 13, 95–107. [Google Scholar] [CrossRef]
- Letellier, C.; Aguirre, L.; Maquet, J.; Aziz-Alaoui, M.A. Should all the species of a food chain be counted to investigate the global dynamics. Chaos Solitons Fract. 2002, 13, 1099–1113. [Google Scholar] [CrossRef]
- Camara, B.I.; Aziz-Alaoui, M.A. Turing and Hopf patterns formation in a predator-prey model with Leslie-Gower-type functional response. Dyn. Contin. Discret. Impuls. Syst. B 2009, 16, 479–488. [Google Scholar]
- Punithan, D.; Kim, D.K.; McKay, R.I.B. Spatio-temporal dynamics and quantification of daisy world in two-dimensional coupled map lattices. Ecol. Complex. 2012, 12, 43–57. [Google Scholar] [CrossRef]
- Rodrigues, L.A.D.; Mistro, D.C.; Petrovskii, S. Pattern formation in a space- and time-discrete predator-prey system with a strong Allee effect. Theor. Ecol. 2012, 5, 341–362. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, H.; Yang, H.; Wang, N.; Zhang, F. Complex patterns in a space- and time-discrete predator-prey model with Beddington-DeAngelis functional response. Commun. Nonlinear Sci. Numer. Simul. 2017, 43, 182–199. [Google Scholar] [CrossRef]
- Maionchi, D.O.; Reis, S.F.D.; Aguiar, M.A.M.D. Chaos and pattern formation in a spatial tritrophic food chain. Ecol. Model. 2006, 191, 291–303. [Google Scholar] [CrossRef]
- Gurney, W.S.C.; Veitch, A.R.; Cruickshank, I.; McGeachin, G. Circles and spirals: Population persistence in a spatially explicit predator-prey model. Ecology 1998, 79, 2516–2530. [Google Scholar] [CrossRef]
- Guckenheimer, J.; Holmes, P. Nonlinear Oscillations, Dynamical Systems and Bifurcations of Vector Fields; Springer: New York, NY, USA, 1983. [Google Scholar]
- Turing, A. The chemical basis of morphogenesis. Philos. Trans. R. Soc. Lond. B 1952, 237, 37–72. [Google Scholar] [CrossRef]
- Bai, L.; Zhang, G. Nontrivial solutions for a nonlinear discrete elliptic equation with periodic boundary conditions. Appl. Math. Comput. 2009, 210, 321–333. [Google Scholar] [CrossRef]









© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Zhang, H.; Ma, S.; Meng, T.; Huang, T.; Yang, H. Self-Organized Patterns Induced by Neimark-Sacker, Flip and Turing Bifurcations in a Discrete Predator-Prey Model with Lesie-Gower Functional Response. Entropy 2017, 19, 258. https://doi.org/10.3390/e19060258
Zhang F, Zhang H, Ma S, Meng T, Huang T, Yang H. Self-Organized Patterns Induced by Neimark-Sacker, Flip and Turing Bifurcations in a Discrete Predator-Prey Model with Lesie-Gower Functional Response. Entropy. 2017; 19(6):258. https://doi.org/10.3390/e19060258
Chicago/Turabian StyleZhang, Feifan, Huayong Zhang, Shengnan Ma, Tianxiang Meng, Tousheng Huang, and Hongju Yang. 2017. "Self-Organized Patterns Induced by Neimark-Sacker, Flip and Turing Bifurcations in a Discrete Predator-Prey Model with Lesie-Gower Functional Response" Entropy 19, no. 6: 258. https://doi.org/10.3390/e19060258
APA StyleZhang, F., Zhang, H., Ma, S., Meng, T., Huang, T., & Yang, H. (2017). Self-Organized Patterns Induced by Neimark-Sacker, Flip and Turing Bifurcations in a Discrete Predator-Prey Model with Lesie-Gower Functional Response. Entropy, 19(6), 258. https://doi.org/10.3390/e19060258