1. Introduction
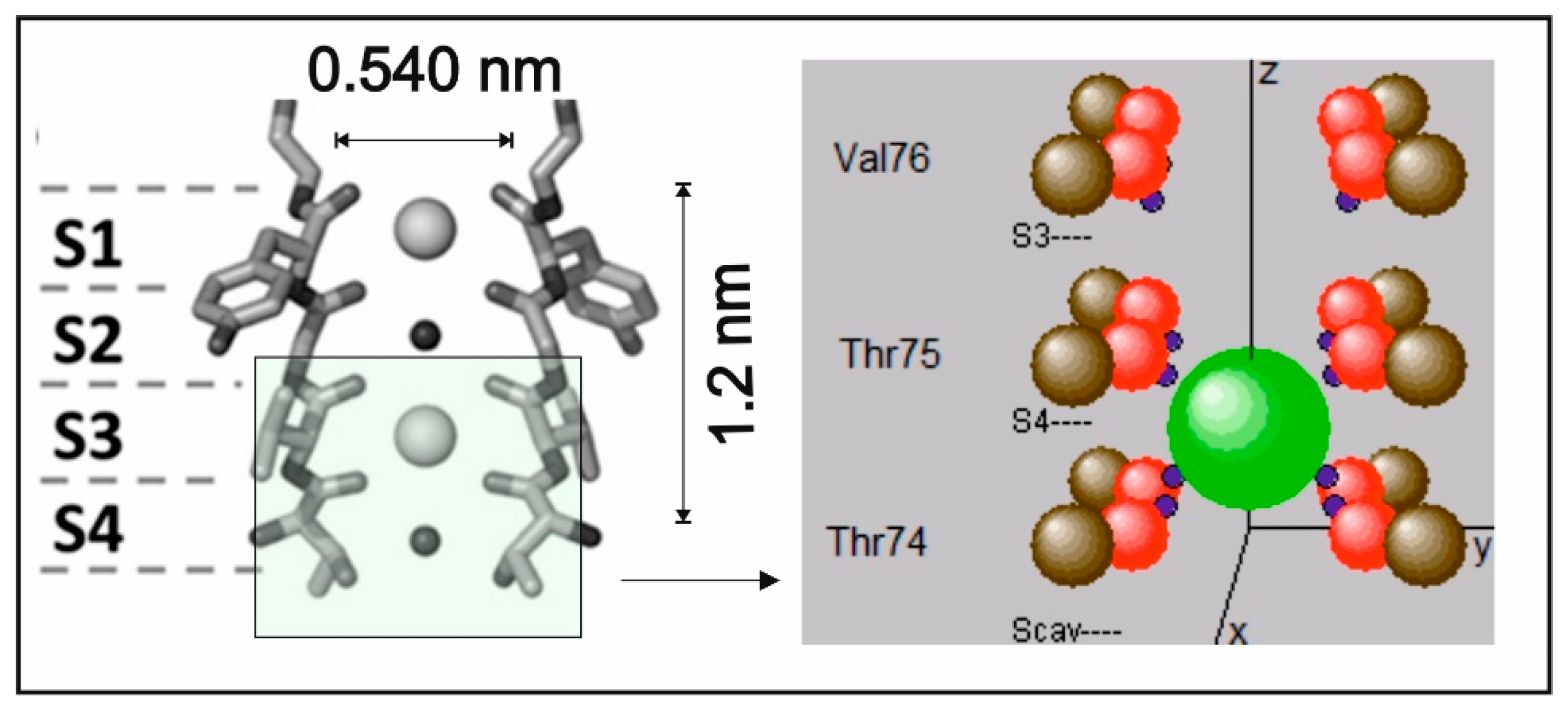
Selective translocation of ions bound to charges across the plasma membrane of cells provides the physical background for the generation and propagation of electrical membrane signals in excitable cells, particularly in nerve cells. The molecules organizing this translocation are provided by membrane-integrated channel proteins, which control the access of ions to permeation (“gating”) in response to changes in transmembrane potentials (“voltage-gating”) and allow very fast ion conduction without loss of selectivity towards certain ion species [
1,
2]. An unprecedented series of studies of these proteins has been initiated after the elucidation of the atomic resolution crystal structure of the prototypic
S. lividans K
+ channel by MacKinnon et al. [
3]. It turned out that the critical domain of the protein that can combine fast transduction close to the diffusion limit with selective preference for the intrinsic ion species is provided by the narrow selectivity filter (SF) of the protein [
4]. In particular, an evolutionary highly conserved sequence of amino acids, the TVGYG (Thr75, Val76, Gly77, Tyr78, Gly79) motive lining the filter region, allows for an inward orientation of backbone carbonyls with oxygen-bound lone pair electrons interacting with the positively charged alkali ions (see
Figure 1). This delicate arrangement involving glycine (Gly79, Gly77) residues serving as “surrogate D-amino acids” [
5] can offer a unique “interaction topology”, mimicking the ions’ hydration shells prior to entering the filter pore. The interactions are realized by short-range attractive (filter atoms) and repulsive (between ion) Coulombic forces [
2,
6].
The initial picture of ion conduction states was built on an alternating sequence of ions and water molecules (e.g., KwKw) passing through the filter with four equally spaced ion binding sites (labeled as S0–S4, as seen from the extra to the intracellular side) [
4]. However, in the course of molecular dynamics (MD) studies this view has become considerably relaxed (e.g., by the observation that different ion permeation mechanisms may coexist energetically [
7], pairwise water-ion hopping mechanisms can occur [
8], the selectivity filter by itself could play a role in gating [
9], and fast permeation involves a direct Coulombic “knock-on” without intermittent water [
10]). Even more important are observations from MD studies calculating potentials of mean force (PMF) within the filter, which demonstrate that the potential barriers for ion translocation are simply too high (>5 kT at 300 K) to be in line with experimental conductance as predicted by the Nernst–Planck equation [
11]. This seems to be a reflection of an enduring problem within the structure-function relations of purely classical MD simulations at the atomic scale and marks the point where the intention of our present contribution becomes significant.
Generally, the short range Coulombic forces coordinating the atoms in the filter reflect quantum-mechanical effects [
2]. As argued before, this requires some quantum dynamics to account for the observed atomic behavior within their molecular environment [
12,
13,
14]. In our previous work, we have suggested a role for a quantum physical description of ion motion through the selectivity domain of K
+ channel proteins. In these studies, we suggested evidence that at least two important features behind ion permeation and gating dynamics can follow naturally if quantum properties are inserted into the underlying equations of motion. First, inserting quantum interference terms into the canonical version of action potential (AP) initiation can reproduce the fast onset characteristic of APs as seen in experimental recordings of cortical neurons [
13]. Second, we have demonstrated evidence for different quantum oscillatory effects within the filter’s atomic environment, which discriminate intrinsic (e.g., K
+) from extrinsic (e.g., Na
+) filter occupations in K
+-type channels [
14].
In the present paper, we go beyond classical MD simulations by treating the motion of the ion itself in a quantum mechanical (QM) context. Because the de Broglie wavelength of thermal ions at 310 K (~0.025 nm) amounts to up to 10% of the spacing in the periodic structure of Coulomb potentials (~0.3 nm) in the nano-pore model of the selectivity filter, the ions wave packet is found to spread out over a certain region. The associated wave dynamics have a coherent short-lived and significant effect on the Coulomb interaction with the surrounding carbonyl charges. Based on first principle methods and solving a non-linear version of the Schrödinger equation, we find that the quantum trajectory of an ion through the filter is accompanied by different time-dependent phase velocities that can exert a favorable effect on the passage of ions through the confining potential landscape of the filter. We suggest that this observation from a combined QM-MD calculation can possibly explain fast conductance without compromising selectivity in the filter of ion channels. We shall discuss the way this favorable effect is exerted and to what extent it lowers the effective potential barriers for ion translocations. Phrased loosely, it is found that the front part of the particle wave function paves the path for the remaining wave components to “sneak through” the open doors of the confining potentials. Due to the involved barriers and masses, this process is different from “particle tunneling” (although it is naturally considered in the solution of the Schrödinger equation calculated with the Crank–Nicolson formalism [
15]). Yet, another finding is remarkable within the context of the enduring debate about quantum coherence times in biological organizations: The quantum characteristic for the present effect not just builds on but also requires very short decoherence times (around 1 ps), a scale that is well within the expected range at biological temperatures [
16].
Finally, it should be mentioned why we deal with a “non-local effect”, as expressed in the title of this paper: Most frequently, the term “non-locality” in QM states refers to a spatial separation between observables preserving a QM correlation (entanglement) between different modes (i.e., pertaining to different sub-systems behaving as one system). Here, we deal with only one technical QM mode or system (i.e., the ion). However, the present finding, that a short and coherent “spread” or “smear” over space of the particle’s mass-bounded charge, according to its QM wave function, can have a strong effect on the dynamic behavior within its environmental potentials implies a functional role for a “non-local property” of a single mode (or system).
2. Methods
Our intention was to observe the K
+ ion during the transition from site S4 to site S3 in the selectivity filter of the KcsA channel (
Figure 1). Therefore, the simulation included the carbonyl groups of Thr74, Thr75, and Val76, as shown in
Figure 1 (right). The backbone carbons were positioned at the widely used coordinates of Guidoni and Garofoli [
17,
18]. The carbonyl oxygen–carbon bond was set initially to 0.123 nm bond length and, in the force-free, unperturbed situation, pointed straight to the central axis (the
z-axis of the coordinate system). Oxygen atoms were allowed to oscillate within horizontal and vertical bending modes, excluding stretching. The effective spring constant and the damping factor of this oscillation were adjusted to the values of typical thermal frequencies in the range of a few THz and to the expected dissipation of vibrational energy into the protein backbone structure after a few oscillation periods. Although considered in the implementation of the program, the short time interval in the present study did not necessitate setting thermal random kicks from backbone atoms to carbonyl atoms (
Table A1). The degrees of freedom for the motion of a single K
+ ion were constrained to the central
z-axis of the selectivity filter. This allowed us to implement all calculations on a quad-core computer within a reasonable processing time and does not influence or restrict the conclusions to be drawn from the present results.
The Coulomb type interaction potential between two particles located at
r1,
r2 and charges
q1,
q2, including a repulsion term with a characteristic distance
rcut (the distance where the electron shells start to overlap), and
ε0 the vacuum dielectric constant, is as follows:
where “point-charges” are located at the center of the particle. K
+ ions carry unit charge and carbonyl-bound C atoms carry partial charges, usually set to +0.38 units. We assigned two-point charges to oxygen atoms; one at the center of the atom, and the second one representing the effective charge center of the lone pair electrons coordinating the K
+ ions and/or water dipoles. The partial charge relocation between the lone pairs and the central O positions was chosen as one of the dynamic variables that determines the depth of the ion-trapping potential (
Table A1).
The classical part of the present MD simulation is based on Verlet’s algorithm applied to Lennard-Jones molecules [
19,
20]. The QM model applies to the motional behavior of the K
+ ion particle waves and was obtained from a non-linear Schrödinger equation (NLSE) (see Equation (2)), with an initial Gaussian wave packet set to an adjustable width and an adjustable mean ion velocity along the
z-axis of the filter. The range of these settings is given in
Appendix A in
Table A1. It is assumed that the wave packet experiences a potential at every instant of time
t, which depends on the position of all other particles at this time. Together with the potential term in Equation (1), this can be described by the following NLSE:
where
mk denotes the mass of the K
+ ion and
rk its position vector along the
z-axis of the filter. Due to the tetrameric lining of the observed motive (
Figure 1), summation over the potential term runs over 12 backbone atomic positions for the carbon, oxygen, and lone pair centers within the Thr74, Thr75, and Val76 lining amino acids shown in
Figure 1 (right). As the atomic positions
ri change in time and are influenced by the position of other atoms, as well as by the probability distribution of the K
+ ion, the situation entails non-linearity in the Schrödinger equation (see also
Appendix B). In the above Equation (2), this functional dependence of
ri on
ψ is explicitly indicated. The linear gradient “
g” expresses the transmembrane electric potential and
zk the z-component of the vector
rk. The parameters that determine the shape and scaling of interaction potentials (i.e., the geometrical embedding) were adjusted to previous models of the KcsA channel [
8,
11]. This implied initial values for
of 0.13 nm, with the charge separation distance of the lone pair electrons from an oxygen center being 1.4 times the radius of the oxygen atom, leading to an average of 0.0825 nm from the oxygen atom’s center. The partial charges of an oxygen atom were split to contain a fraction of 30% at central locations and 70% in the lone pair charge point location (
Table A1).
It should be noted that the NLSE given by Equation (2) restricts the motion of the K+ ion to the z-axis and does not include an expansion of the wave packet perpendicular to this axis. This restriction was necessary to keep the computational time within reasonable limits. In addition, the narrow extension and the symmetry of the filter lining in the pore cause sideways forces to mostly cancel each other along the filter’s z-axis. For the further formal description, this implies that the position vector of the K+ ion is essentially given by its z-component , because its x- and y-components are always zero.
Prior to solving the SE given by Equation (2), we have to take into account the experience of forces by the surrounding carbonyl C and O atoms due to the interacting K
+ wave. At this stage, we assume the backbone C atoms to be rigid (see
Section 4) but allow for two bending modes of O atoms while keeping the CO distance constant. The differential
dz along the
z-axis of the wave packet is then found to exert a differential force
as follows:
The total force
acting from K
+ on this O atom is then obtained by integrating over the range defined along
z (additional forces acting on this O atom are a restoring force and a decelerating force, as well as attraction/repulsion of the surrounding C and O atoms, see
Appendix C.) This will subsequently change the locations
ri of the O atoms and thereby the potential term in the SE acting back on the evolution of the wave packet. The effect of Equation (3) introduces a non-linearity into the SE as shown in Equation (2). The resulting non-linear Schrödinger equation (NLSE) is formally similar but causally different from the description of Bose–Einstein condensation (BEC) at ultra-cold temperatures [
21]. This is because under BEC conditions, the probability distribution of the wave function enters into the Hamiltonian itself, while in our case it is the effect integrated over time, as shown in
Appendix B. We solved Equation (2) together with Equation (3) in very small-time steps by the Crank-Nicolson method [
15] to keep track of the QM phase factor but sampled the positional changes of the O atoms in larger time steps (values given in
Appendix A, additional explanations on the NLSE derivation in
Appendix B).
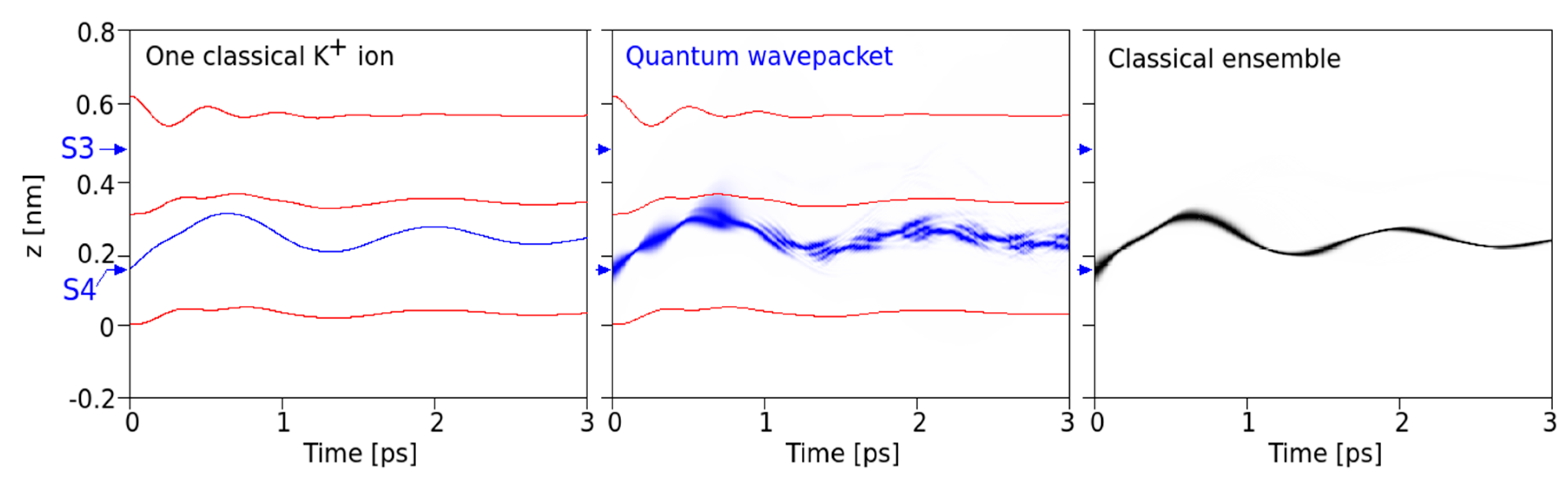
We simulated the behavior of classical ensembles to compare with the above-described quantum behavior in
Figure 2,
Figure 3,
Figure 4 and
Figure 5. In these simulations, classical particles were set into motion at 10
2 different starting positions. The positions were generated equidistantly within three times the full 1/e-width of the initial quantum wave packet and weighted with the probability density of this packet. At each initial location 10
2, particles were set into motion with velocities, again sampled equidistantly from within three times the full 1/e-width of the initial Gaussian momentum distribution of the QM wave packet.
The numerical implementation of the present methods was designed to offer an interactive control window that allowed us to change the settings given in the
Appendix A for path integration according to Equations (2) and (3), and it delivered the following graphs for time-dependent wave functions and probability plots (
Figure 2 and
Figure 3).
4. Discussion
We have investigated the motional behavior of a single K+ ion between two transition sites in the nano-pore selectivity filter motive of the KcsA channel from two different perspectives: A quantum mechanical simulation implemented by a non-linear version of the Schrödinger equation and a corresponding classical ensemble behavior under identical initial and interaction terms. The non-linear Schrödinger model (NLSE) integrates the solution of the wave equation into its interaction potentials, with all surrounding charges modulating the probability distribution of the wave at a given instance of time. This offers a kind of recursive approach, taking account of mutual interactions of confining Coulomb forces and the QM wave equation, a situation that seems more realistic than the calculation of potentials of mean force (PMF) from classical MD at the atomic scale. The methods were implemented in Java. An executable version is available upon request, as well as a source citation agreement (this requires prior installation of the Java Runtime Environment).
First, our results provide a comparison between classical and QM implementations, which reveal a high degree of similarity in the overall, time-dependent behavior during a several pico-second time interval (
Figure 2). The observed similarity is particularly obvious for the ensemble behavior of ions and certainly signals a high level of consistency of the implemented methods. Besides this similarity at the “caged” state, as shown in
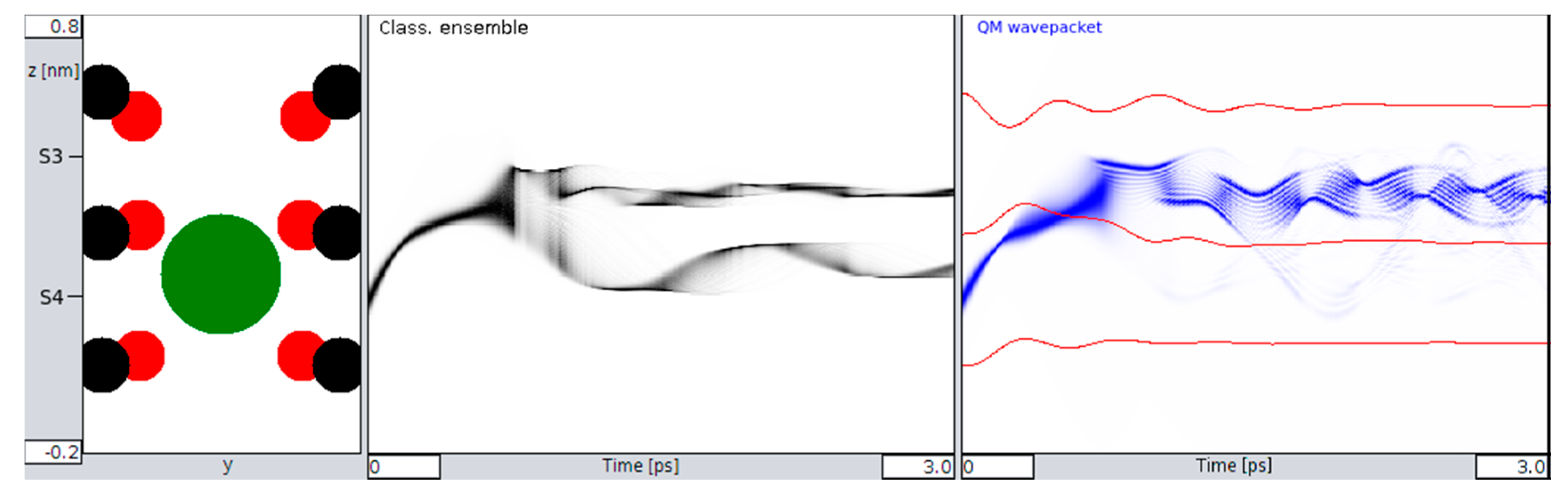
Figure 2, we also observe small but significant differences in the time evolution of QM and classical ions prior to transitions from S4 to S3 that bear the “seeds” for subsequent QM classical dissociations during the passage from one binding site to the next site. Following this initial situation, the site transition favored by higher onset velocities marks a clear difference between the behavior of the QM wave packet and the classical ensemble of ions. At a critical velocity of ions, the classical probability splits into ½ of its population, with 50% crossings to S3 and the rest remaining in S4 (
Figure 3 and
Figure 4). Within the same time, and under the same initial conditions, the QM packet, however, can cross to S3 almost completely. An increase in efficiency to cross the barriers of about 50% could be expected to increase permeation rates at larger scales by a similar amount (i.e., within a complete filter occupancy and including intermittent water molecules in the filter domain). Compared with previous reports about the discrepancy of calculated energetic barrier heights from PMF methods (between 5 and 10 kcal/mol) with observed and “effective permeation heights” as required by Nernst–Planck estimates of <~3 kcal/mol at 300 K [
11], the ratio of this difference lays well within the range of permeation enhancement as predicted here. In other words, assigning a short (1 ps) coherent quantum mechanical property to the atoms motion interacting with the carbonyl derived forcefields can explain fast conduction speeds, whereas purely classical models cannot.
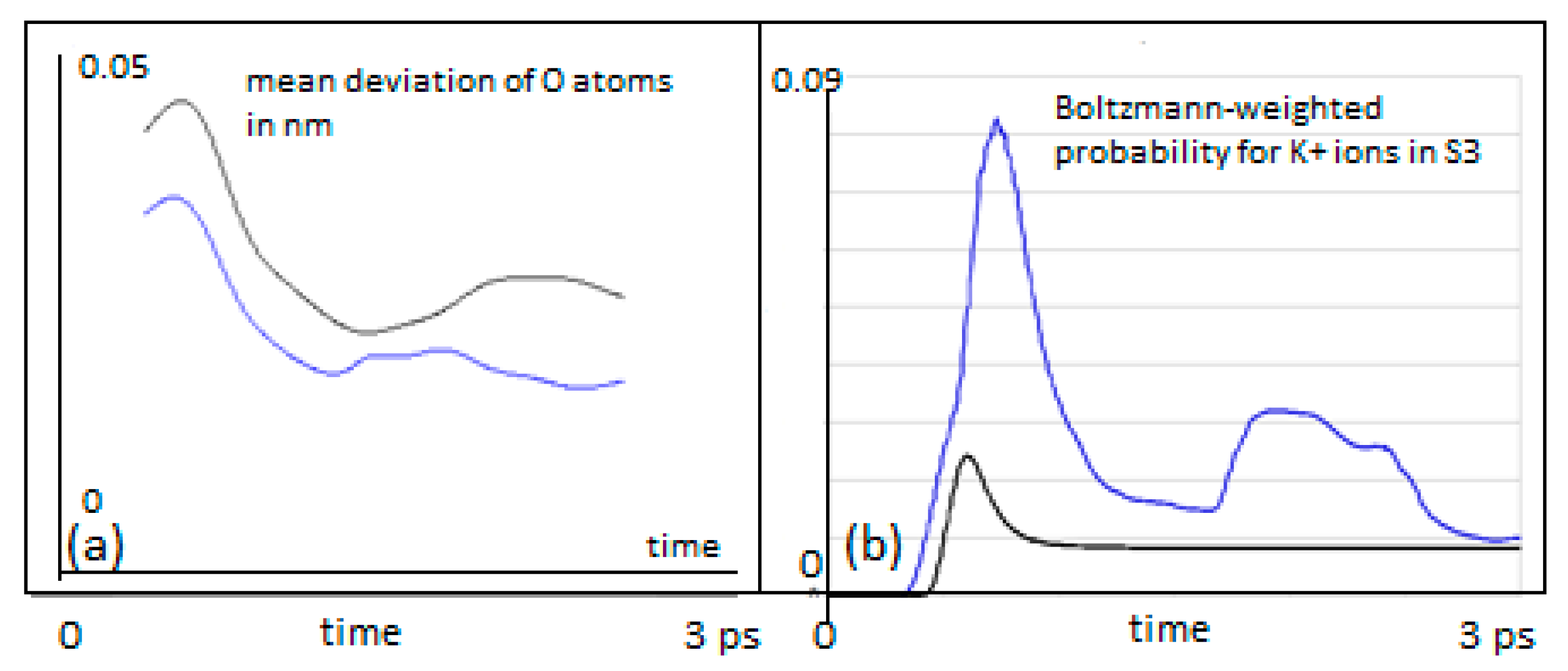
It must be granted that the present report focuses on a small and short time-scaled view on filter dynamics. This may somehow insufficiently sample the more complete conformational pattern underlying filter conduction states. However, the main findings of the present study strongly suggest that a small scaled and ultra-short-lived quantum state of the permeating ion is not just sufficient but also indispensable to explain fast conduction without compromising selectivity in the filter. The observations demonstrated in
Figure 4 and
Figure 5 indicate that due to the dispersion of the wave function of a “quantum ion”, the coordinating Coulomb forces from surrounding oxygen charges become dissociated to different parts of the wave function. This effect reduces the effective barrier that the ion has to cross. In a metaphorical view, it looks like the faster front part of the particle wave can open the door to the barrier, allowing the slower tail parts to sneak through this door. Due to the engaged height and width of the barrier, this effect is not typical “leap-frogging” (passing over), nor “quantum-tunneling”, although there is some formal resemblance to the latter. The resemblance is that the potential energy of the separating barrier is also a function of the state variable of the system. But, as opposed to tunneling, the system actually “manipulates” the barrier to be able to cross it. It is perhaps best described as “
quantum sneaking” through potential barriers.
The present study takes into account physiological temperatures with respect to ion motion and velocities. At this stage, the effects of these temperatures on thermally induced protein backbone and carbonyl vibrations are not included. Our numerical implementations do offer the extension to thermal carbonyl atomic motions during the temporal evolution of the K+ wave packet. We presently implement these fluctuations by repetitions of K+ evolutions during time-varying random fluctuations of the surrounding O and C atoms. We expect, however, that the main findings reported here about the difference between a classical ensemble and the K+ wave packet will remain largely resistant to these thermal vibrations. The reason is that the main effect found here is due to the spatial dispersion of the QM wave packet, which in turn dynamically spreads the interacting force directions of the surrounding and coordinating charges. In the pure classical case, these forces would permanently be directed towards the center of the moving ion. It is just this distinction that allows for what we called “quantum sneaking” in the discussion above.
Finally, our observations render an important role for decoherence and the quantum to classical transition as predicted in an earlier paper by one of the authors [
22]. Fast decoherence of the ions wave function after about 1 ps, a time when almost all of its probability distribution has penetrated the barrier, leads to classical behavior, which can be seen to avoid the return to the previous location. So, decoherence actually “guides” the particle into one direction in the filter, and an oscillation between quantum and classical states cooperate in a directed transport through the potential landscape of the filter.
A possible role of the important and unique QM interaction, the environment-induced, dynamical destruction of quantum coherence deserves some further remarks in the present context. It was not our intention to study decoherence in our simulation explicitly, and we did not include the scattering elements and processes that induce decoherence in the evolving wave packet. In some previous work, we and a co-author of this group (V. Salari) have provided a list of scattering sources and interacting scattering events applicable to the same atomic configuration and dynamics of the KcsA filter model as used in the present study [
14,
16,
23]. The results of these studies suggest that we can expect decoherence times for K
+ ions in the filter model around one or a few pico-seconds at warm temperatures.
The intention of the present simulation was more focused on a potential functional role of decoherence during ion permeation by implementing a comparison between quantum and classical motions. As we can expect that decoherence of the QM wave packet will lead to a certain resemblance with a classical behavior in the course of time, comparisons as those shown in
Figure 4 and
Figure 5b can give us some inferential information about the time and the role of decoherence. It turns out, that short decoherence times, exactly within the range of the predictions mentioned from the scattering studies above (i.e., around 1 ps), could play a highly beneficial role for the successful transition from S4 to S3. A transition to classical behavior due to decoherence after this time would actually “stabilize” the ion’s location at S3 once it has reached this site. The original transition probability would still be at the level of the high QM initial transfer probability. We therefore conjecture that short coherent QM states, in the range of a few ps, are of an advantage for the observed high ion transfer rates without compromising ion coordination. Taken together, we suggest that the quantum dynamics behind the ion motion in the filter open the door through confining potentials, and decoherence guides the moving atoms through the specific path offered by the selectivity filter of channel proteins. To the best of our knowledge, this is one of the first reports about a decisive role of quantum decoherence for an ancient and highly conserved mechanism of membrane signaling in biology.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




