Entropic Equilibria Selection of Stationary Extrema in Finite Populations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
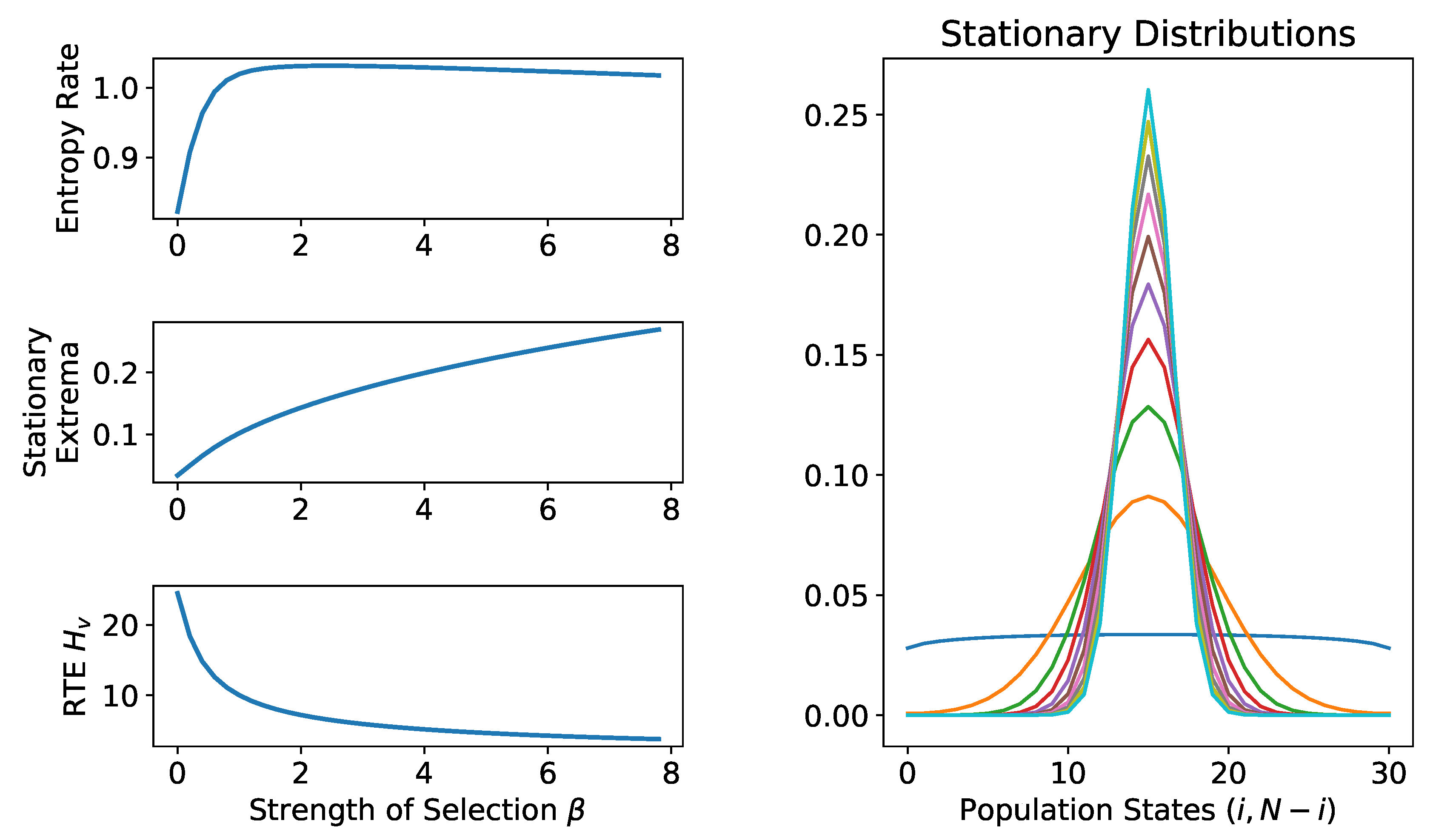
2.1. Stationary Distributions, Entropy Rates, and Random Trajectory Entropies
2.2. Applications
2.2.1. Moran Process with Mutation
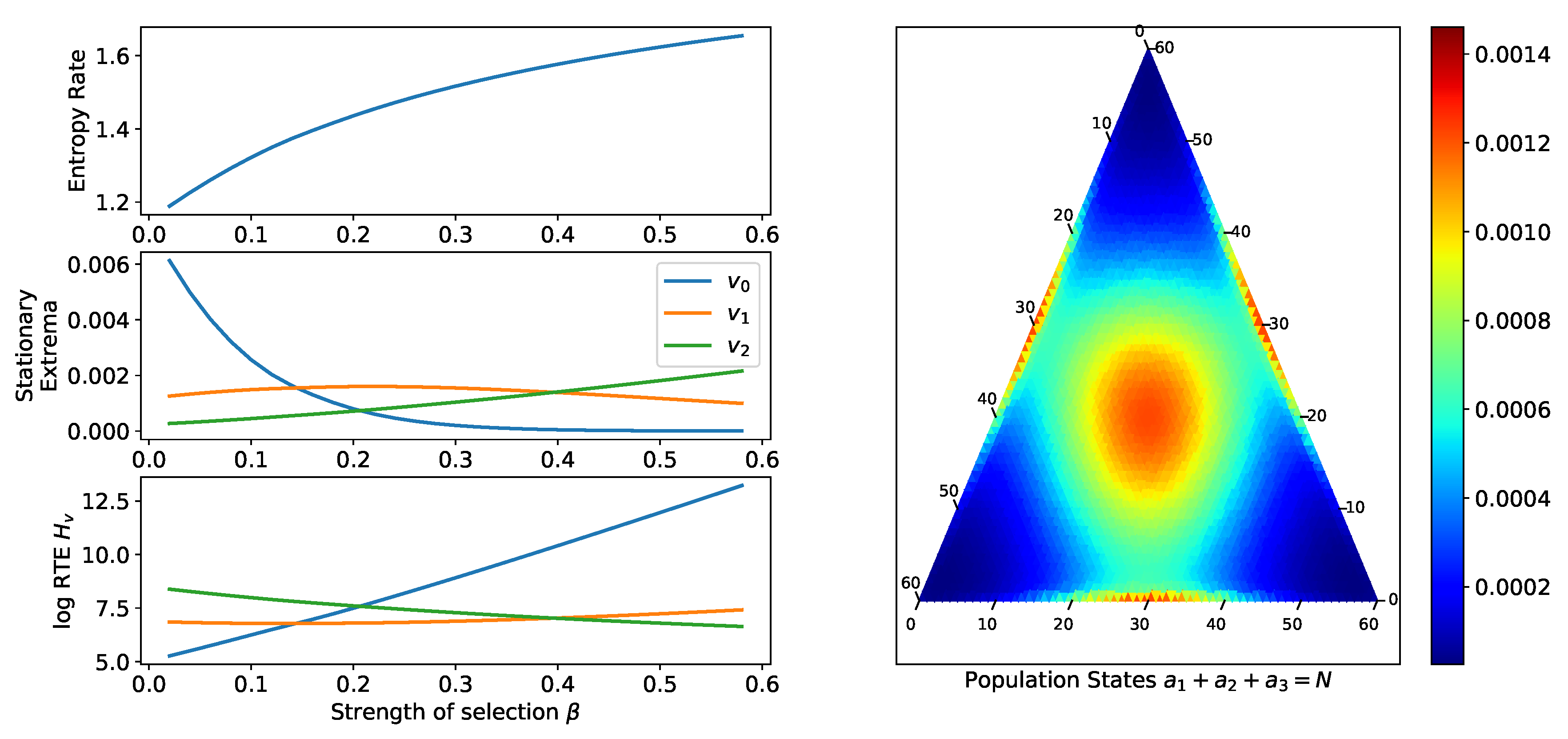
2.2.2. Comparison of Equilibria of a Single Process
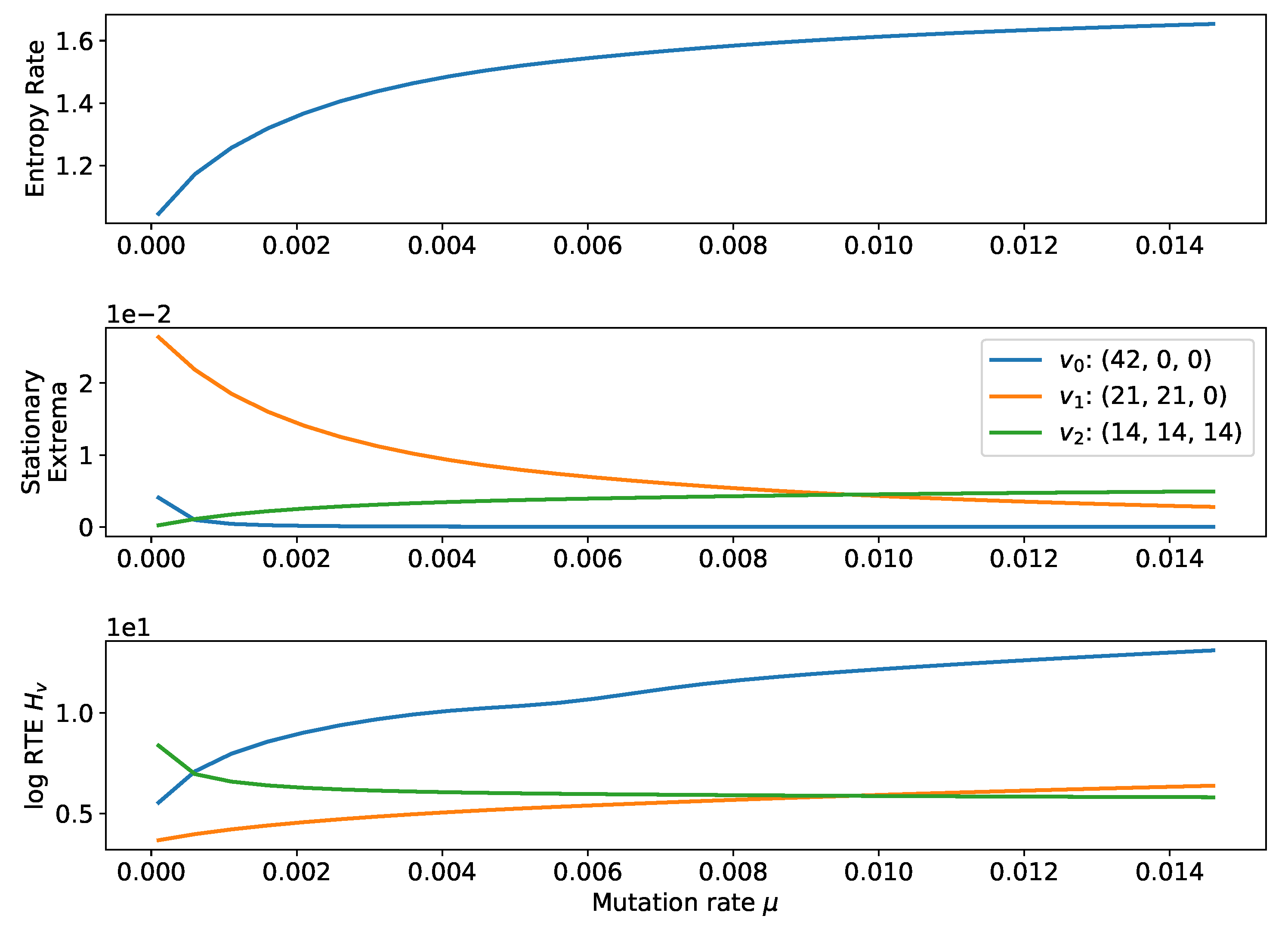
2.2.3. Small Mutation Limit
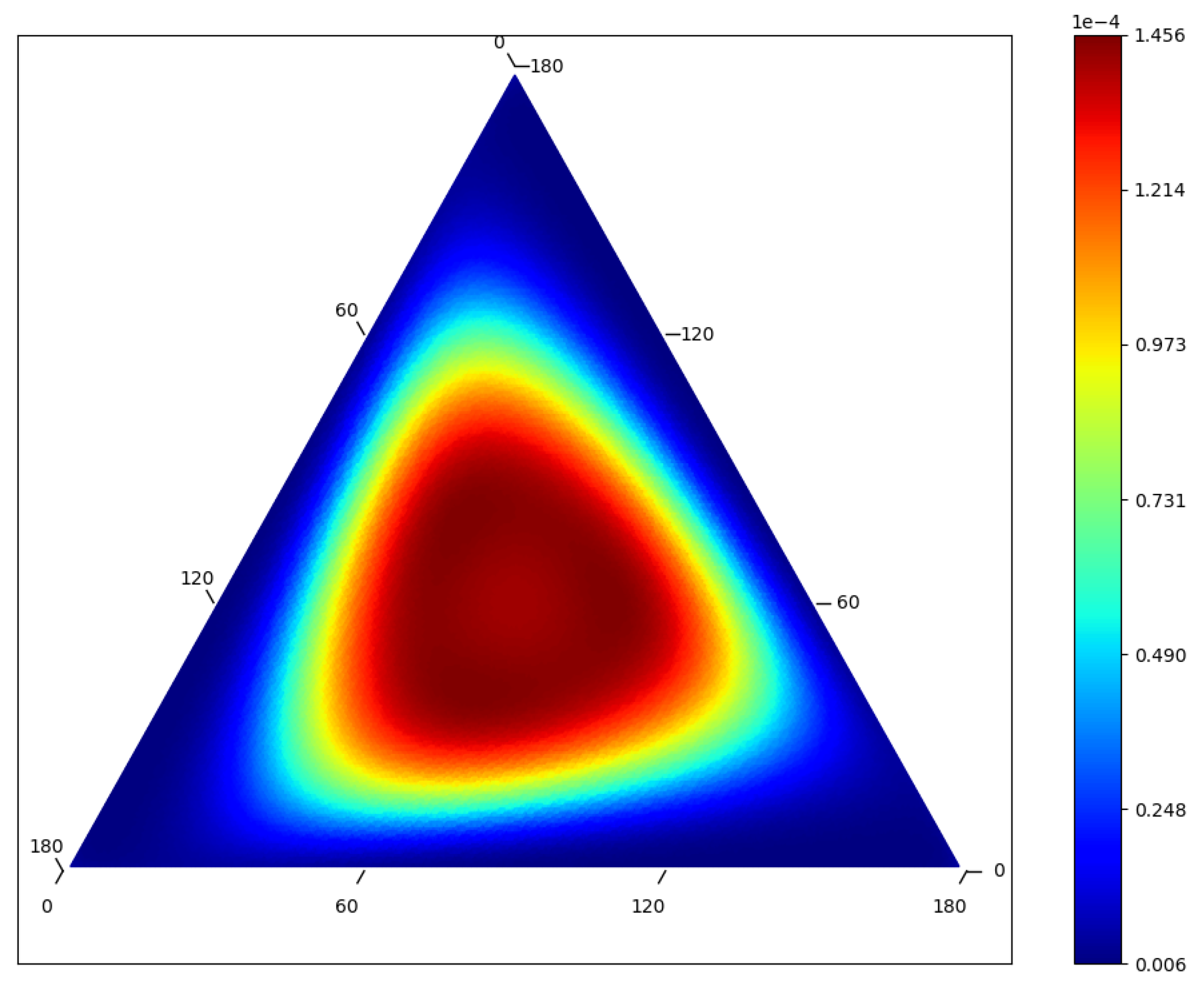
2.2.4. Large Populations and Neutral Landscapes
2.2.5. Comparison of Equilibria for Separate Processes on the Same States
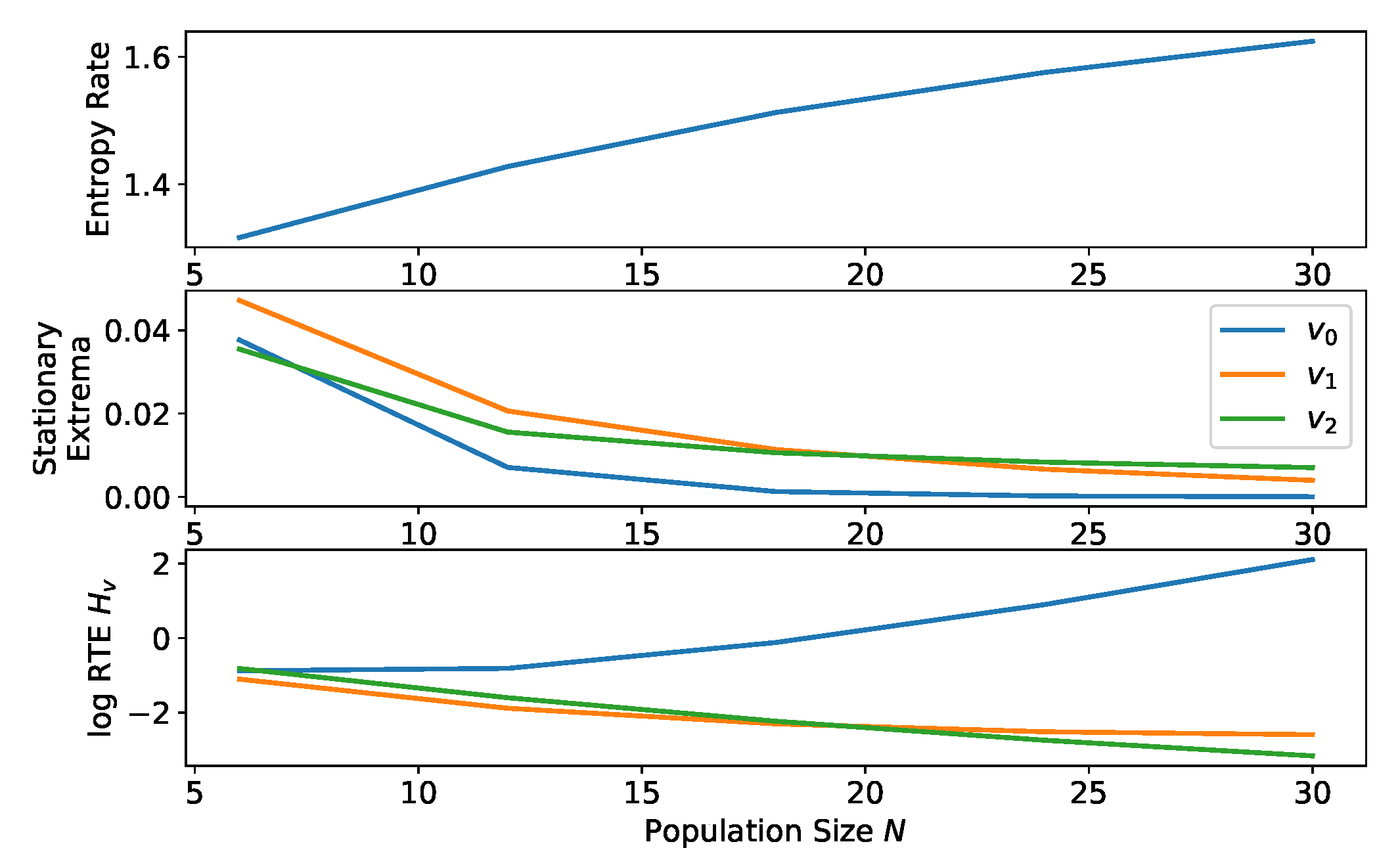
2.2.6. Comparison of Equilibria for Process with Varying Population Size
3. Discussion
4. Methods and Materials
- Given values for all relevant parameters (e.g., a fitness landscape, , N, ), generate the matrix of transition probabilities;
- Compute the stationary distribution of the process, which can be done efficiently (in memory and time) with a power iteration and sparse matrix implementation, or simply standard matrix multiplication for processes with small state spaces;
- With the stationary distribution and the matrix of transition probabilities, compute the entropy rate using Equation (1);
- For each state, compute the RTE using Equation (4) using the entropy rate and the stationary distribution.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harper, M.; Fryer, D. Stationary Stability for Evolutionary Dynamics in Finite Populations. Entropy 2016, 18, 316. [Google Scholar] [CrossRef]
- Smith, J.M. Evolution and The Theory of Games; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Ekroot, L.; Cover, T.M. The Entropy of Markov Trajectories. IEEE Trans. Inf. Theor. 1993, 39, 1418–1421. [Google Scholar] [CrossRef]
- Kafsi, M.; Grossglauser, M.; Thiran, P. The Entropy of Conditional Markov Trajectories. IEEE Trans. Inf. Theor. 2013, 59, 5577–5583. [Google Scholar] [CrossRef] [Green Version]
- Samuelson, L. Evolutionary Games and Equilibrium Selection; The MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Harsanyi, J.C.; Selten, R. A General Theory of Equilibrium Selection in Games; The MIT Press: Cambridge, MA, USA, 1988. [Google Scholar]
- Hordijk, A.; Ridder, A. Insensitive Bounds for The Stationary Distribution of Non-reversible Markov Chains. J. Appl. Probab. 1988, 25, 9–20. [Google Scholar] [CrossRef]
- Fudenberg, D.; Imhof, L.; Nowak, M.A.; Taylor, C. Stochastic Evolution As a Generalized Moran Process. Unpublished work. 2004. [Google Scholar]
- Claussen, J.C.; Traulsen, A. Non-Gaussian Fluctuations Arising from Finite Populations: Exact Results for The Evolutionary Moran Process. Phys. Rev. E 2005, 71, 025101. [Google Scholar] [CrossRef] [PubMed]
- Moran, P.A.P. The Statistical Processes of Evolutionary Theory. Am. J. Hum. Genet. 1962, 14, 438–439. [Google Scholar]
- Hofbauer, J.; Sigmund, K. Evolutionary Game Dynamics. Bull. Am. Math. Soc. 2003, 40, 479–519. [Google Scholar] [CrossRef]
- Weibull, J.W. Evolutionary Game Theory; The MIT Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Hofbauer, J.; Sigmund, K. Evolutionary Games and Population Dynamics; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Lynch, M. Evolution of The Mutation Rate. Trends Genet. 2010, 26, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Traulsen, A.; Hauert, C. Stochastic Evolutionary Game Dynamics. Rev. Nonlinear Dyn. Complex. 2009, 2, 25–61. [Google Scholar]
- Harper, M. Inherent Randomness of Evolving Populations. Phys. Rev. E 2014, 89, 032709. [Google Scholar] [CrossRef] [PubMed]
- Harper, M. Entropy Rates of The Multidimensional Moran Processes and Generalizations. arXiv, 2014; arXiv:1401.2713. [Google Scholar]
- Blume, L.E. The Statistical Mechanics of Strategic Interaction. Games Econ. Behav. 1993, 5, 387–424. [Google Scholar] [CrossRef]
- Harper, M. Stationary: Release 1.0.0. 2018. Available online: https://zenodo.org/record/1343125#.W3-HtbgRVPY (accessed on 23 May 2018).
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Harper, M.; Weinstein, B. Python-Ternary: Release 1.0.4. 2018. Available online: https://github.com/marcharper/python-ternary (accessed on 23 May 2018).





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harper, M.; Fryer, D. Entropic Equilibria Selection of Stationary Extrema in Finite Populations. Entropy 2018, 20, 631. https://doi.org/10.3390/e20090631
Harper M, Fryer D. Entropic Equilibria Selection of Stationary Extrema in Finite Populations. Entropy. 2018; 20(9):631. https://doi.org/10.3390/e20090631
Chicago/Turabian StyleHarper, Marc, and Dashiell Fryer. 2018. "Entropic Equilibria Selection of Stationary Extrema in Finite Populations" Entropy 20, no. 9: 631. https://doi.org/10.3390/e20090631
APA StyleHarper, M., & Fryer, D. (2018). Entropic Equilibria Selection of Stationary Extrema in Finite Populations. Entropy, 20(9), 631. https://doi.org/10.3390/e20090631