Image Entropy for the Identification of Chimera States of Spatiotemporal Divergence in Complex Coupled Maps of Matrices



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Preliminary Notes and the Objective
2.1. A Network of Coupled Maps
2.2. A Network of Coupled Map of Matrices
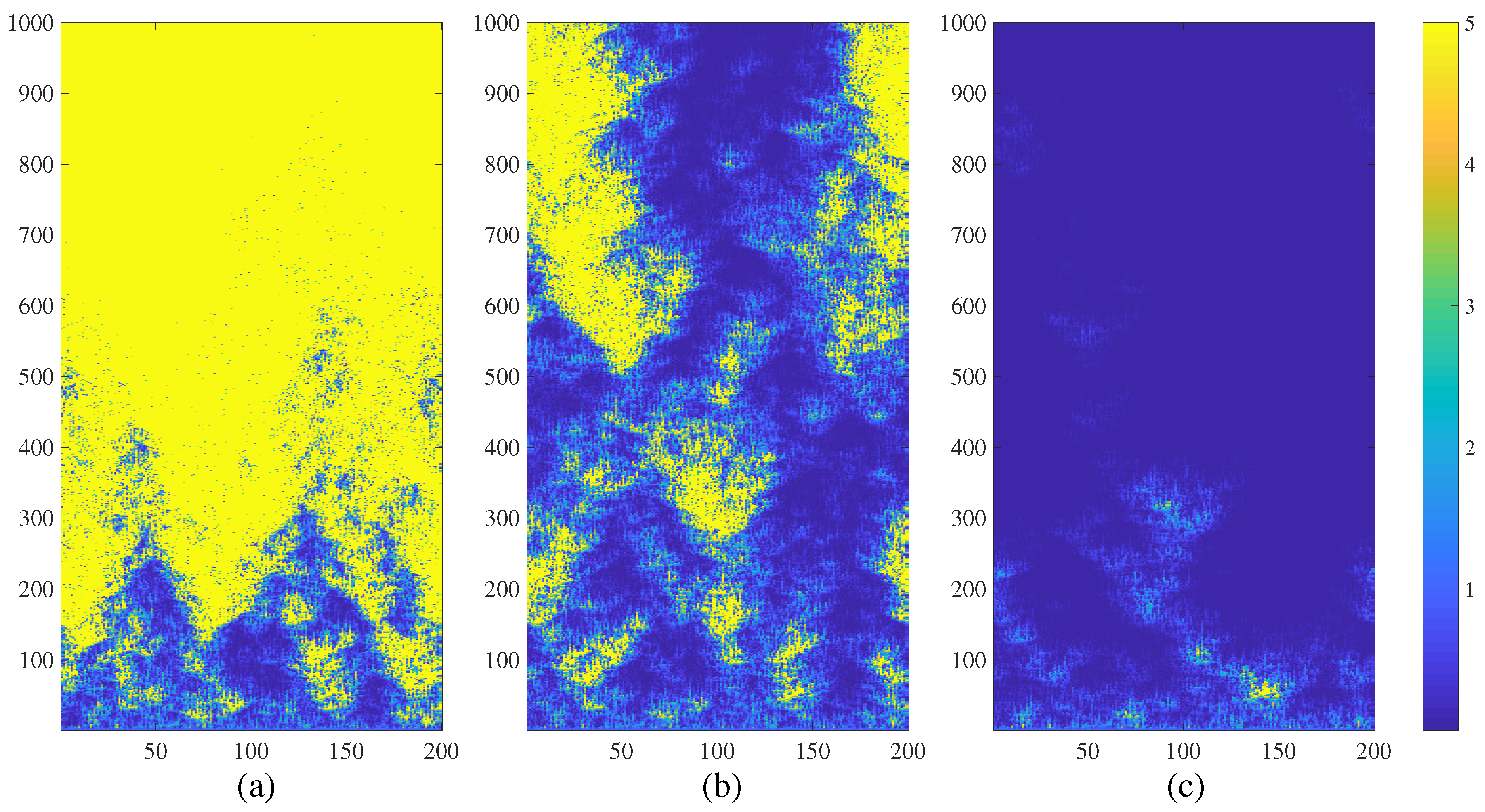
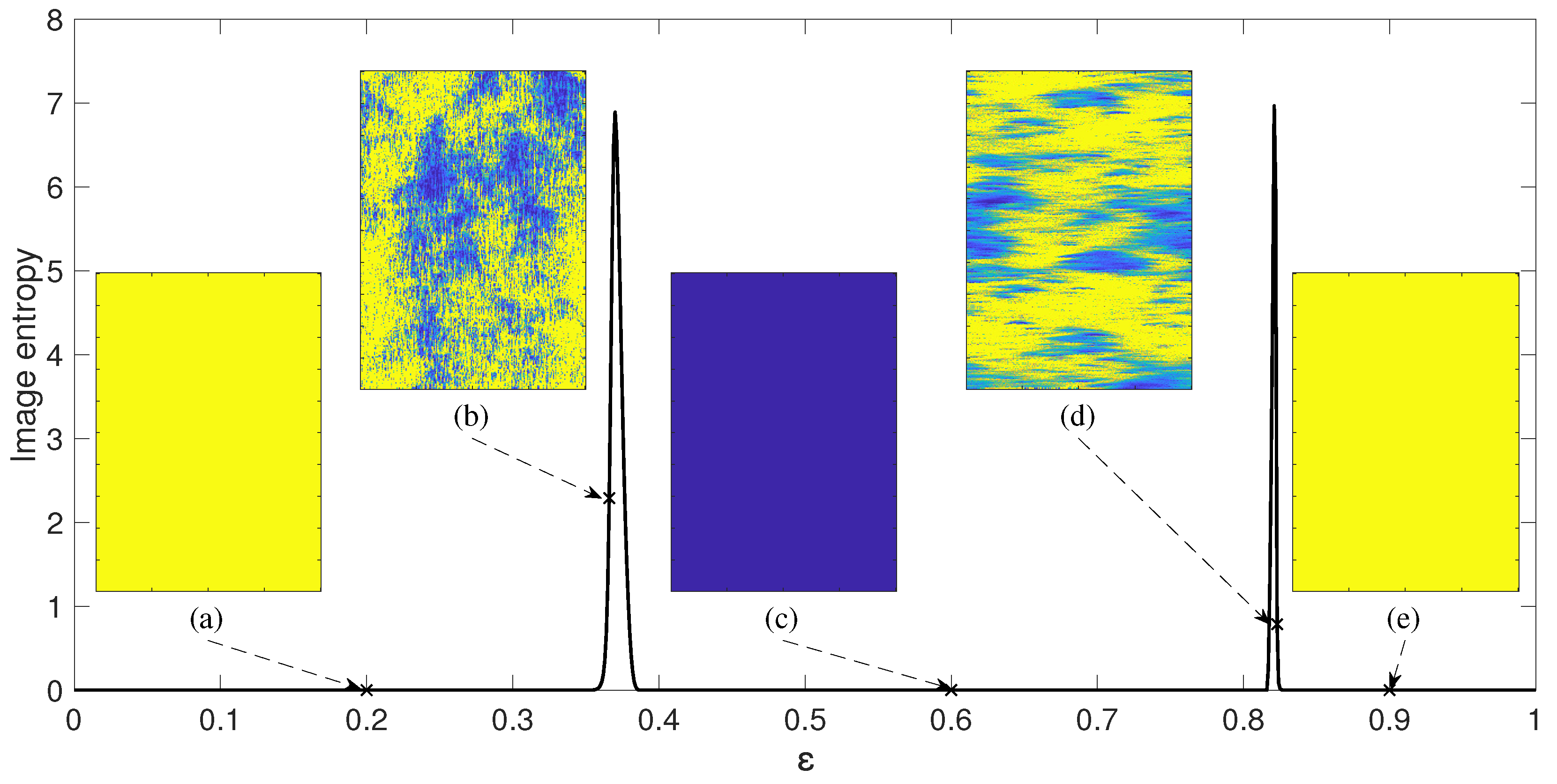
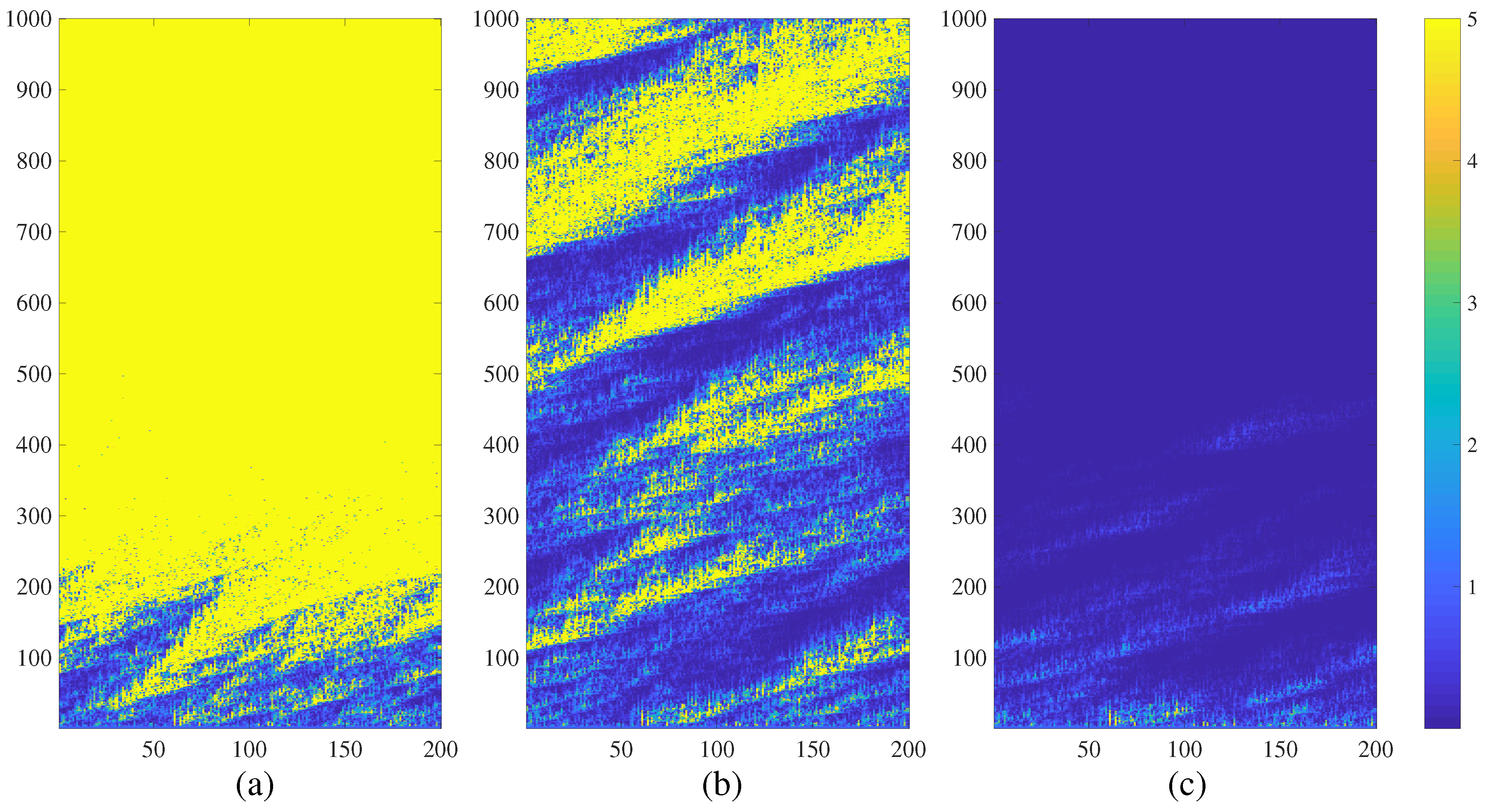
3. Chimera States of Spatiotemporal Divergence in Regular NCMMs
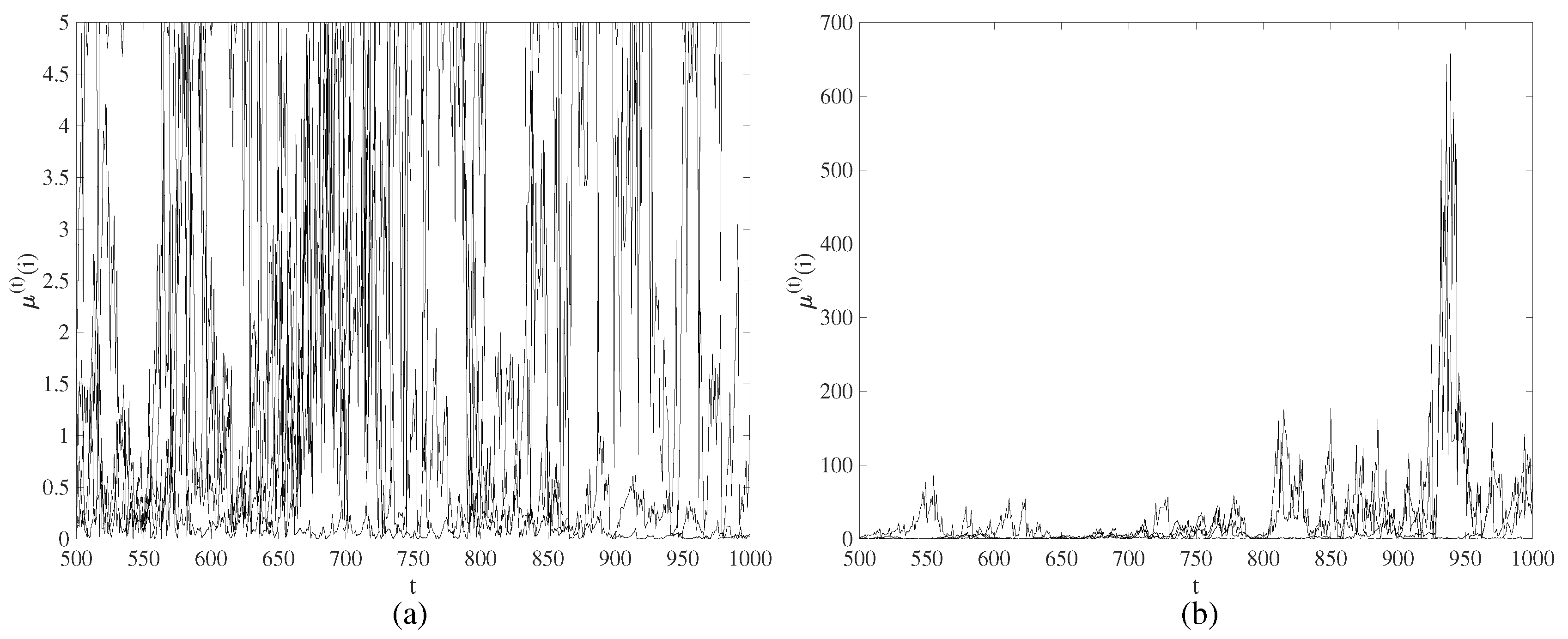
3.1. Spatiotemporal Divergence in a Regular NCMM
3.2. Chimera States of Spatiotemporal Divergence in a Regular Feed-Forward NCMM
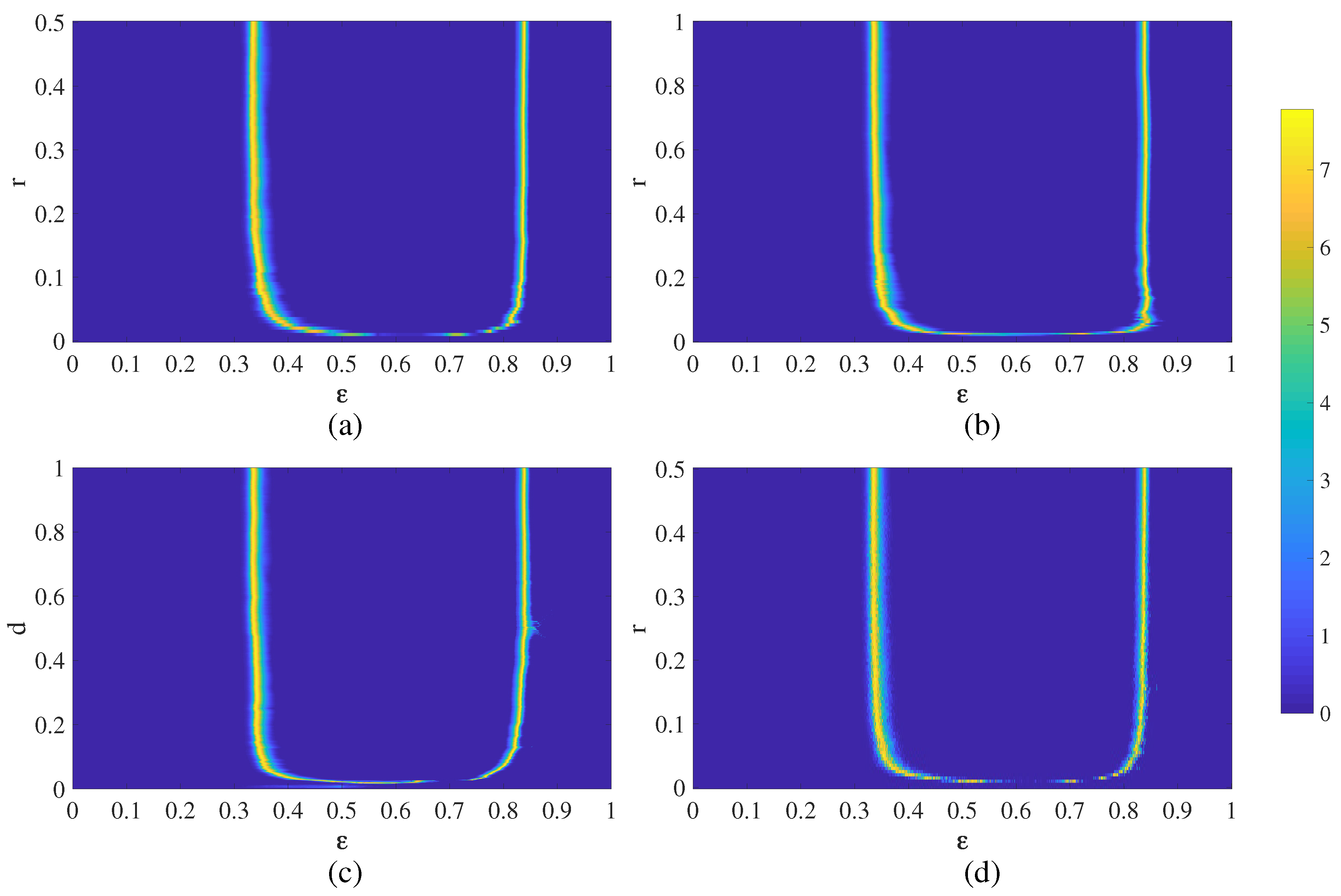
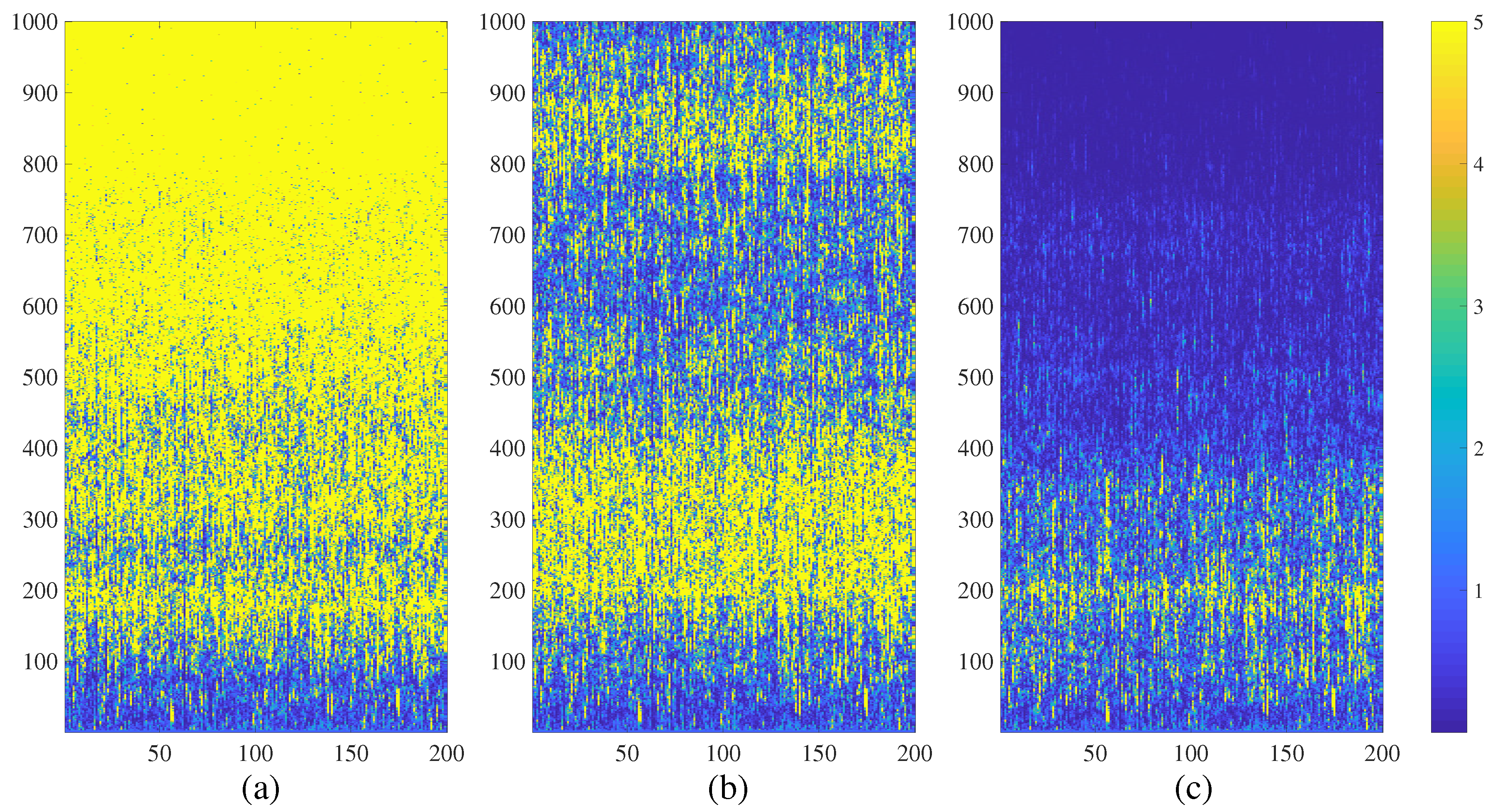
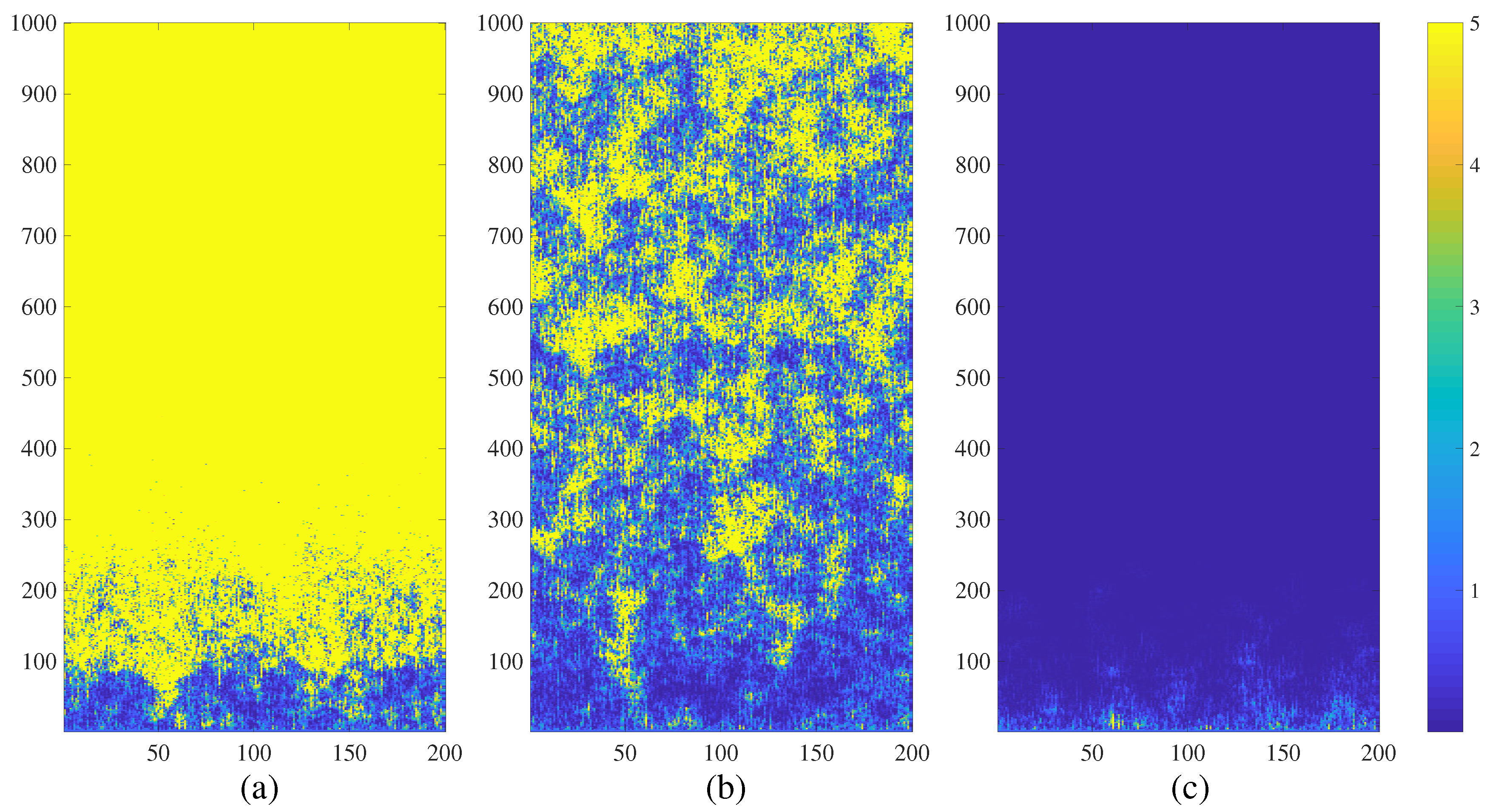
4. Chimera States of Spatiotemporal Divergence in a Complex NCMM
4.1. Chimera States of Spatiotemporal Divergence in the Erdős-Rényi NCMM
4.2. Chimera States of Spatiotemporal Divergence in the Small-World NCMM
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Kuramoto, Y.; Battogtokh, D. Coexistence of coherence and incoherence in nonlocally coupled phase oscillators. Nonlinear Phenom. Complex Syst. 2002, 5, 380–385. [Google Scholar]
- Zakharova, A.; Kapeller, M.; Schöll, E. Chimera Death: Symmetry Breaking in Dynamical Networks. Phys. Rev. Lett. 2014, 112, 154101. [Google Scholar] [CrossRef] [Green Version]
- Panaggio, M.J.; Abrams, D.M. Chimera states: Coexistence of coherence and incoherence in networks of coupled oscillators. Nonlinearity 2015, 28, 67–87. [Google Scholar] [CrossRef]
- Bukh, A.; Strelkova, G.; Anishchenko, V. Spiral wave patterns in a two-dimensional lattice of nonlocally coupled maps modeling neural activity. Chaos Soliton Fractals 2019, 120, 75–82. [Google Scholar] [CrossRef]
- Martens, E.A.; Thutupalli, S.; Fourrière, A.; Hallatschek, O. Chimera states in mechanical oscillator networks. Proc. Natl. Acad. Sci. USA 2013, 110, 10563–10567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, J.D.; Bansal, K.; Murphy, T.E.; Roy, R. Experimental observation of chimera and cluster states in a minimal globally coupled network. Chaos 2016, 26, 094801. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Bi, R.; Sun, Y.X.; Zhang, S.; Song, Q.Q. Chimera states in Gaussian coupled map lattices. Front. Phys. 2018, 13, 130502. [Google Scholar] [CrossRef]
- Xu, H.Y.; Wang, G.L.; Huang, L.; Lai, Y.C. Chaos in Dirac electron optics: Emergence of a relativistic quantum chimera. Phys. Rev. Lett. 2018, 120, 124101. [Google Scholar] [CrossRef]
- Nkomo, S.; Tinsley, M.R.; Showalter, K. Chimera States in Populations of Nonlocally Coupled Chemical Oscillators. Phys. Rev. Lett. 2013, 110, 244102. [Google Scholar] [CrossRef] [Green Version]
- Totz, J.F.; Rode, J.; Tinsley, M.R.; Showalter, K.; Engel, H. Spiral wave chimera states in large populations of coupled chemical oscillators. Nat. Phys. 2018, 14, 282–286. [Google Scholar] [CrossRef]
- Majhi, S.; Bera, B.K.; Ghosh, D.; Perc, M. Chimera states in neuronal networks: A review. Phys. Life Rev. 2018. [Google Scholar] [CrossRef]
- Hizanidis, J.; Kanas, V.G.; Bezerianos, A.; Bountis, T. Chimera states in networks of nonlocally coupled Hindmarsh-Rose neuron models. Int. J. Bifurc. Chaos 2014, 24, 1450030. [Google Scholar] [CrossRef]
- Shepelev, I.A.; Bukh, A.V.; Strelkova, G.I.; Vadivasova, T.E. Chimera states in ensembles of bistable elements with regular and chaotic dynamics. Nonlinear Dyn. 2017, 90, 2317–2330. [Google Scholar] [CrossRef]
- Malchow, A.K.; Omelchenko, I.; Schöll, E.; Hövel, P. Robustness of chimera states in nonlocally coupled networks of nonidentical logistic maps. Phys. Rev. E 2018, 98, 012217. [Google Scholar] [CrossRef]
- Bogomolov, S.A.; Slepnev, A.V.; Strelkova, G.I.; Schöll, E.; Anishchenko, V.S. Mechanisms of appearance of amplitude and phase chimera states in ensembles of nonlocally coupled chaotic systems. Commun. Nonlinear Sci. Numer. Simul. 2017, 43, 25–36. [Google Scholar] [CrossRef]
- Bukh, A.; Rybalova, E.; Semenova, N.; Strelkova, G.; Anishchenko, V. New type of chimera and mutual synchronization of spatiotemporal structures in two coupled ensembles of nonlocally interacting chaotic maps. Chaos Interdiscipl. J. Nonlin. Sci. 2017, 27, 111102. [Google Scholar] [CrossRef] [PubMed]
- Laing, C.R. Chimeras in networks with purely local coupling. Phys. Rev. E 2015, 92, 050904(R). [Google Scholar] [CrossRef] [PubMed]
- Clerc, M.G.; Coulibaly, S.; Ferré, M.A.; García-Ñustes, M.A.; Rojas, R.G. Chimera-type states induced by local coupling. Phys. Rev. E 2016, 93, 052204. [Google Scholar] [CrossRef] [Green Version]
- Kundu, S.; Majhi, S.; Bera, B.K.; Ghosh, D.; Lakshmanan, M. Chimera states in two-dimensional networks of locally coupled oscillators. Phys. Rev. E 2018, 97, 022201. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, L.; Krischer, K. Clustering as a Prerequisite for Chimera States in Globally Coupled Systems. Phys. Rev. Lett. 2015, 114, 034101. [Google Scholar] [CrossRef]
- zur Bonsen, A.; Omelchenko, I.; Zakharova, A.; Schöll, E. Chimera states in networks of logistic maps with hierarchical connectivities. Eur. Phys. J. B 2018, 91, 65. [Google Scholar] [Green Version]
- Omelchenko, I.; Provata, A.; Hizanidis, J.; Schöll, E.; Hövel, P. Robustness of chimera states for coupled FitzHugh-Nagumo oscillators. Phys. Rev. E 2015, 91, 022917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, M.A.; Goltsev, A.V. Distinct dynamical behavior in Erdős-Rényi networks, regular random networks, ring lattices, and all-to-all neuronal networks. Phys. Rev. E 2019, 99, 022303. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, J.; Omelchenko, I.; Zakharova, A.; Schöll, E. Chimera states in complex networks: Interplay of fractal topology and delay. Eur. Phys. J. Spec. Top. 2017, 226, 1883–1892. [Google Scholar] [CrossRef]
- Zhu, Y.; Zheng, Z.; Yang, J. Chimera states on complex networks. Phys. Rev. E 2014, 89, 022914. [Google Scholar] [CrossRef]
- Li, B.; Saad, D. Chimera-like states in structured heterogeneous networks. Chaos 2017, 27, 043109. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Zakharova, A.; Jalan, S. Non-identical multiplexing promotes chimera states. Chaos Soliton Fractals 2018, 106, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Hizanidis, J.; Kouvaris, N.E.; Zamora-López, G.; Díaz-Guilera, A.; Antonopoulos, C.G. Chimera-like States in Modular Neural Networks. Sci. Rep. 2016, 6, 19845. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.V.; Kundu, S.; Kirsanov, D.V.; Frolov, N.S.; Maksimenko, V.A.; Ghosh, D.; Dana, S.K.; Hramov, A.E. Multiscale interaction promotes chimera states in complex networks. Commun. Nonlinear Sci. 2019, 71, 118–129. [Google Scholar] [CrossRef]
- May, R.M. Simple mathematical models with very complicated dynamics. Nature 1976, 261, 459–467. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; He, Y.; Wang, X.Y. Spatiotemporal chaos in mixed linear-nonlinear two-dimensional coupled logistic map lattice. Phys. A 2018, 490, 148–160. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, H. Bifurcation, chaos and pattern formation in a space-and time-discrete predator-prey system. Chaos Soliton Fractals 2016, 91, 92–107. [Google Scholar] [CrossRef]
- Fernandez, B. Selective chaos of travelling waves in feedforward chains of bistable maps. arXiv 2018, arXiv:1811.08310. [Google Scholar]
- Guangqing, L.; Smidtaite, R.; Navickas, Z.; Ragulskis, M. The effect of explosive divergence in a coupled map lattice of matrices. Chaos Soliton Fractals 2018, 113, 308–313. [Google Scholar]
- Navickas, Z.; Smidtaite, R.; Vainoras, A. The logistic map of matrices. Discret. Cont. Dyn. B 2011, 3, 927–944. [Google Scholar]
- Miranda, G.H.B.; Machicao, J.; Bruno, O.M. Exploring spatio-temporal dynamics of cellular automata for pattern recognition in networks. Sci. Rep. 2016, 6, 37329. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Lu, Q.S. Spatiotemporal patterns and chaotic burst synchronization in a small-world neuronal network. Phys. A Stat. Mech. Its Appl. 2008, 387, 3719–3728. [Google Scholar] [CrossRef]
- Sakyte, E.; Ragulskis, M. Self-calming of a random network of dendritic neurons. Neurocomputing 2011, 74, 3912–3920. [Google Scholar] [CrossRef]
- Cross, M.C.; Hohenberg, P.C. Pattern formation outside of equilibrium. Rev. Mod. Phys. 1993, 65, 851. [Google Scholar] [CrossRef]
- Goldman, M.S. Memory without feedback in a neural network. Neuron. 2009, 61, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Zankoc, C.; Fanelli, D.; Ginelli, F.; Livi, R. Desynchronization and pattern formation in a noisy feed-forward oscillator network. Phys. Rev. E 2019, 99, 012303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, R.V.; Valverde, S. Information theory of complex networks: On evolution and architectural constraints. In Complex Networks; Springer: Berlin/Heidelberg, Germany, 2004; pp. 189–207. [Google Scholar]
- Erdős, P.; Rényi, A. On Random Graphs. Publ. Math. 1959, 6, 290–297. [Google Scholar]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’networks. Nature 1998, 393, 440. [Google Scholar] [CrossRef]
- Shinoda, K.; Kaneko, K. Chaotic Griffiths Phase with Anomalous Lyapunov Spectra in Coupled Map Networks. Phys. Rev. Lett. 2016, 117, 254101. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smidtaite, R.; Lu, G.; Ragulskis, M. Image Entropy for the Identification of Chimera States of Spatiotemporal Divergence in Complex Coupled Maps of Matrices. Entropy 2019, 21, 523. https://doi.org/10.3390/e21050523
Smidtaite R, Lu G, Ragulskis M. Image Entropy for the Identification of Chimera States of Spatiotemporal Divergence in Complex Coupled Maps of Matrices. Entropy. 2019; 21(5):523. https://doi.org/10.3390/e21050523
Chicago/Turabian StyleSmidtaite, Rasa, Guangqing Lu, and Minvydas Ragulskis. 2019. "Image Entropy for the Identification of Chimera States of Spatiotemporal Divergence in Complex Coupled Maps of Matrices" Entropy 21, no. 5: 523. https://doi.org/10.3390/e21050523
APA StyleSmidtaite, R., Lu, G., & Ragulskis, M. (2019). Image Entropy for the Identification of Chimera States of Spatiotemporal Divergence in Complex Coupled Maps of Matrices. Entropy, 21(5), 523. https://doi.org/10.3390/e21050523