
Algal Bloom Ties: Spreading Network Inference and Extreme Eco-Environmental Feedback



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Algal Blooms as the Epitome of Marine Ecosystem Health
1.2. Complex Marine Ecosystems
1.3. Ecological Patterns as Chlorophyll-a Spreading Networks
2. Materials and Methods
2.1. Datasets
2.2. Ecosystem Organization and Connectome
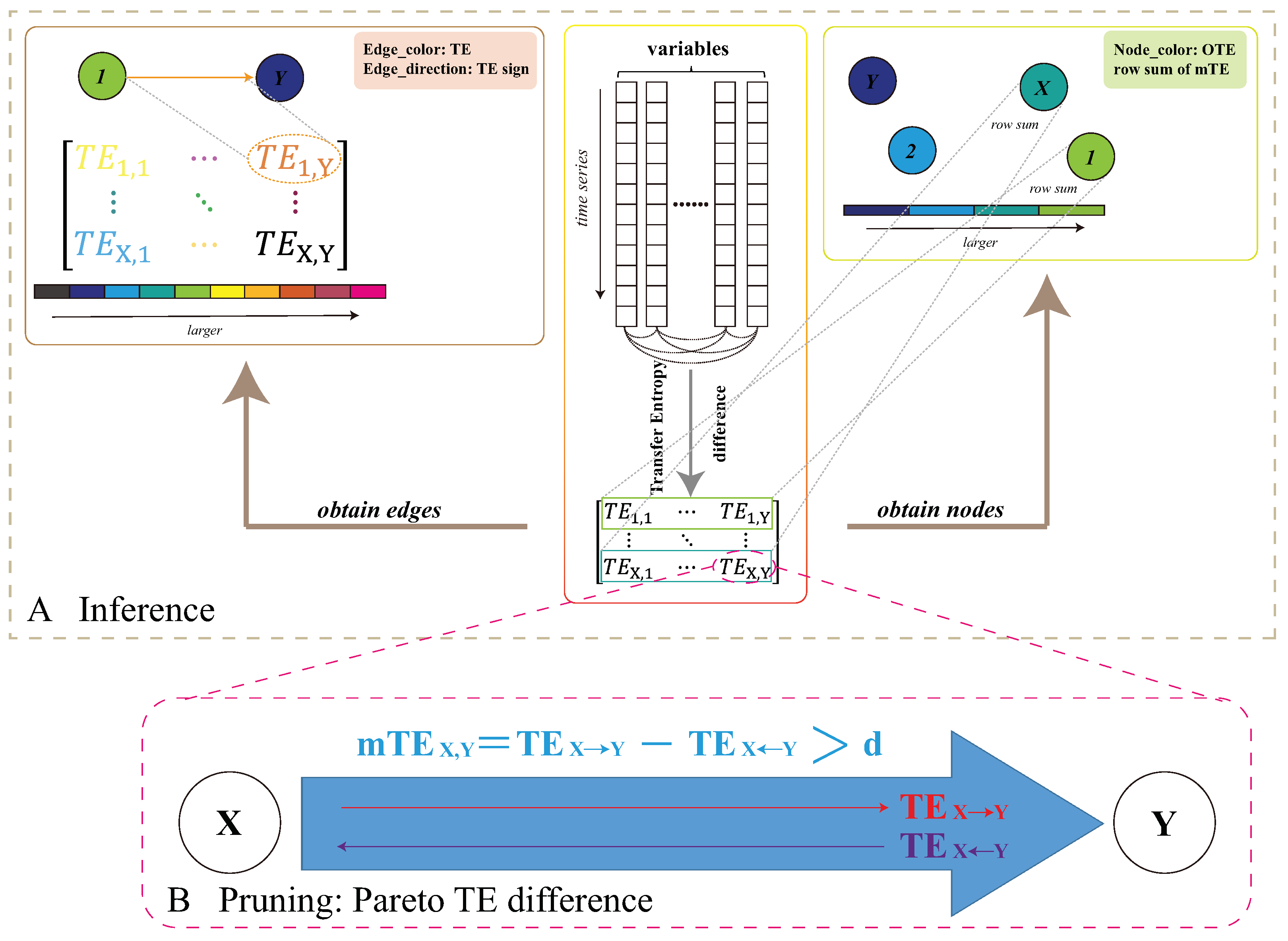
2.3. Eco-Environmental Network Inference
2.4. Eco-Environmental Factor Predictive Causality
3. Results and Discussion
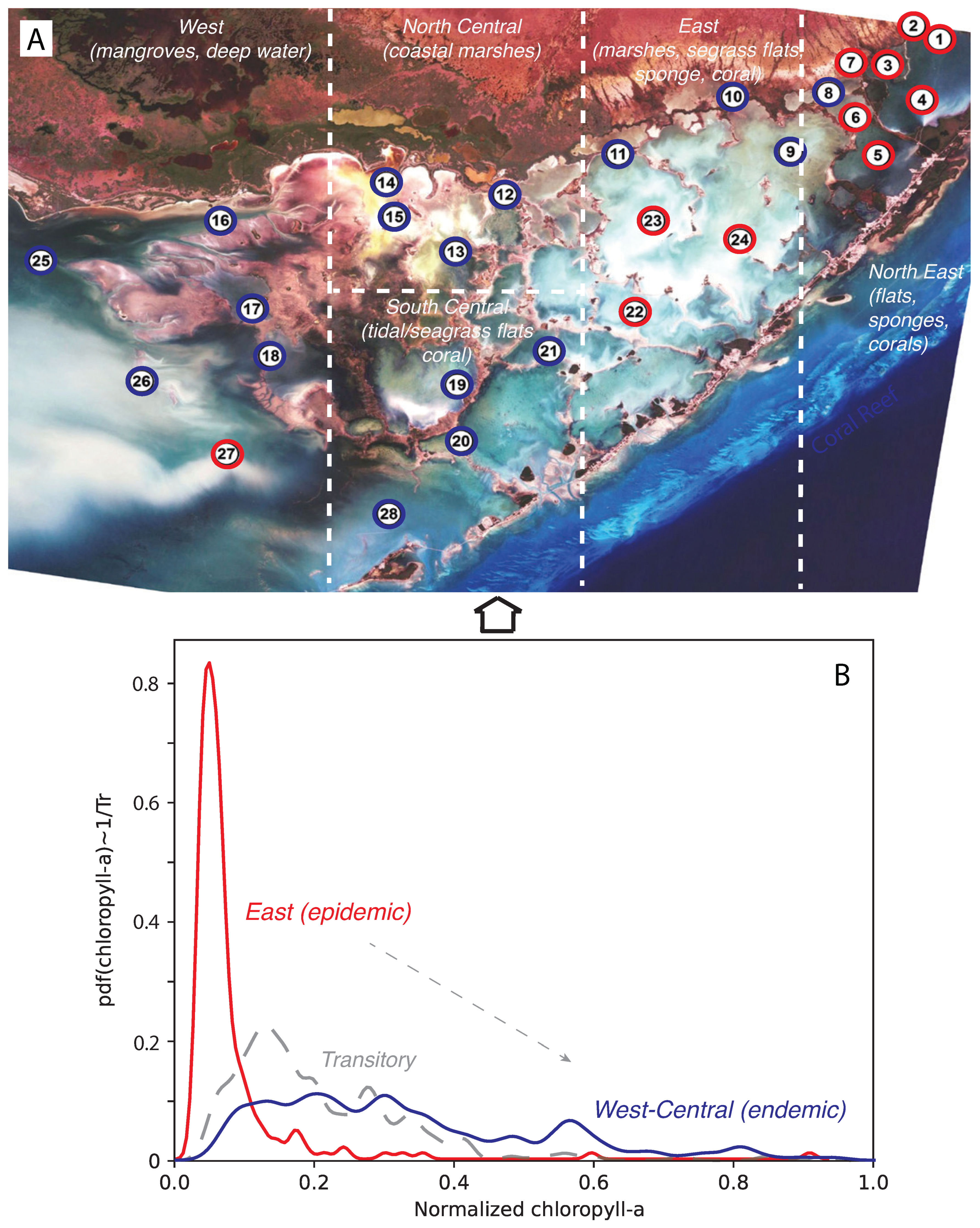
3.1. Spatio-Temporal Spreading and Fluctuations
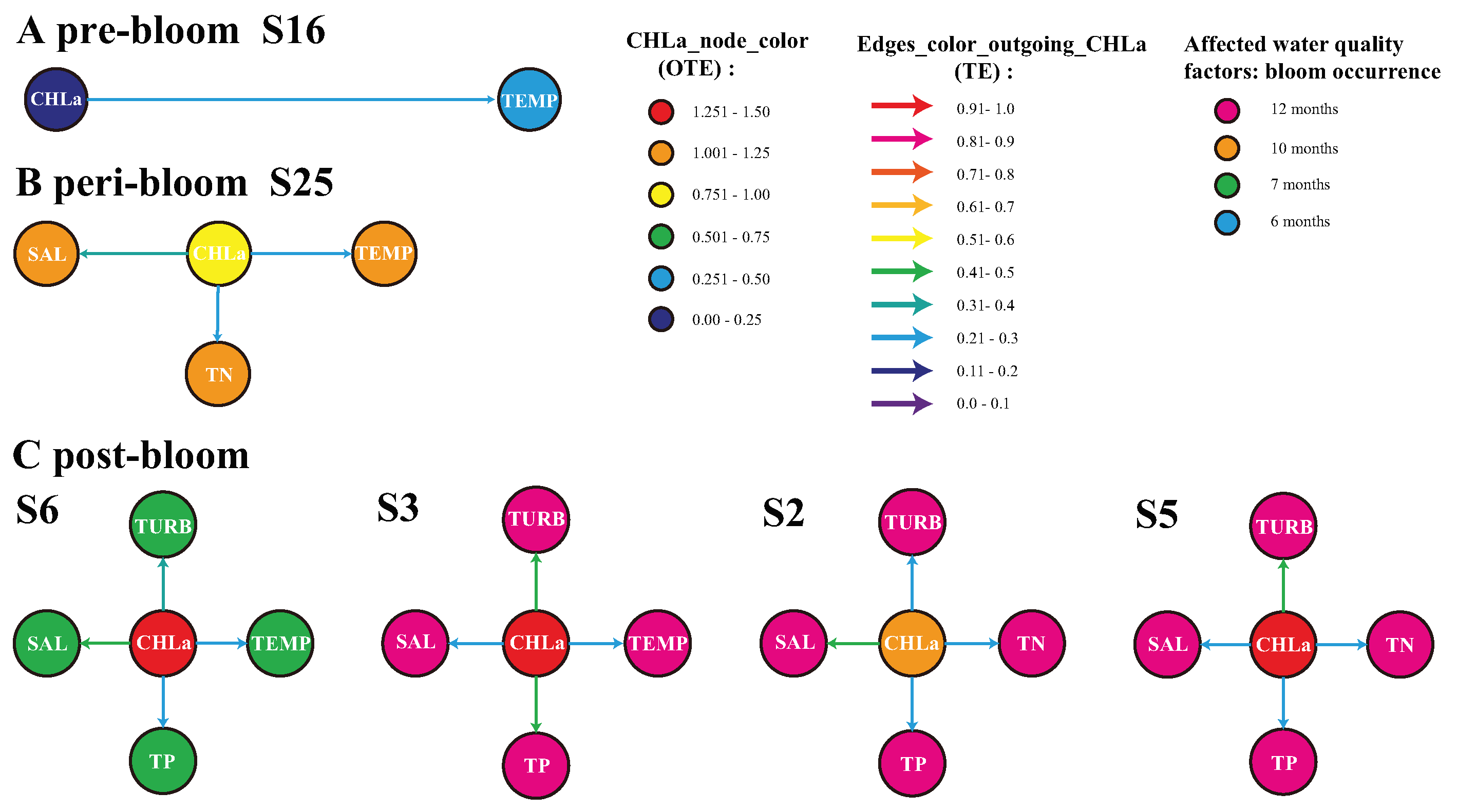
3.2. Water Quality Trends and Bloom Impacts
3.3. Bloom Intensity and Area Dependency
4. Conclusions
- We showed how CHLa patterns carry information regarding the underpinning ecohydrological networks (and associated spreading determinants, such as nutrients) that support ecosystem function and services. Salient Pareto interactions were defined via thresholding TE differences with a threshold of causal significance that was set to consider the top 20% of TEs (related to the tail of scale-free CHLa probability distribution function), i.e., necessary and sufficient interactions to predict the risk of bloom spreading.More generally, the discovery and inference of the ”ecosystem connectome” (as biogeochemical determinant and spreading networks) allows for the assessment of ecosystem health (quantified by the proximity to an optimal condition, such as the non-bloom state) as well as the investigation of causal determinants and their sources, proximity to ecosystem shifts and targeted ecohydrological controls.
- Through spatial analysis of bloom spreading networks, we showed how regions not previously involved in blooms (i.e., the highly biodiverse NE tidal-flat habitats with corals and sponges) were caused by large imbalances of CHLa in the western and central blooms, which were causally involved. The latter regions were characterized by CHLa that was more randomly distributed and a higher probability of CHLa extremes. This probabilistic structure, reflecting the spatial distribution of CHLa, is likely tipping eastern regions to similar bloom endemics. From the perspective of complex networks, this bloom event (2004–2006) evolved from a spatial network with a localized trigger area and a small-world topology to a random topology with long-range spatial diffusion.In 2005, when most stations were blooming, the spatial spreading network was scale-free (theoretically optimal in a purely topological and predictive sense [55,56]) with a random biogeochemical network, including CHLa (topologically suboptimal), which underpins the dichotomy between structural and functional networks for ecological risks.
- In terms of temporal dynamics, subsequent to the first bloom outbreak, persistent and recurring blooms were observed for several NE areas with long-lasting environmental impacts on turbidity and salinity aggravated by temperature increases. Bloom sources were related to central coastal marshes and, to a lower extent, mangrove habitats. We further showed that blooms were a recurring and persistent phenomenon over a long period of time with continuous outbreaks in interdependent regions. This led to higher energy dissipation and larger instability dictated by the more random distribution of CHLa, which was associated with a more uniform network with long-range connectivity regardless of habitats, likely leading to the loss of ecological heterogeneity.
- The analysis of biogeochemical factors affecting water quality showed that the occurrence of blooms could only affect small fluctuations of temperature at the beginning of the blooms; however, repeated bloom outbreaks largely affected other biogeochemical factors (such as salinity, turbidity and CHLa triggering hysteresis or memory effects) that are poorly systemically controllable due to the loss of vegetation and other keynote species.The concentration of CHLa can be influenced by temperature and salinity, and changes in the CHLa concentration can, in turn, have indirect effects on water temperature through various ecological processes. In some regions, facilitated by shallow-water habitats, a water temperature increase can stimulate phytoplankton growth and increase the concentration of CHLa. The increased CHLa can, in turn, absorb more sunlight, which can lead to local warming of the water.In the long term, the persistence of blooms, i.e., high CHLa, may also alter nutrient cycling as highlighted by other studies with the term “oceanic positive feedback mechanism” [11], and our model was able to infer this secondary causal pathway together with the primary one, where temperature change led to CHLa change and blooms. This underscores that bloom management should start from the source, otherwise blooms’ environmental impacts will gradually expand and become uncontrollable, thus, also affecting the ecosystem stability and resilience and settling into undesired ecological states.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Smayda, T.J. What is a bloom? A commentary. Limnol. Oceanogr. 1997, 42, 1132–1136. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Yang, S.; Zhao, D.; Hu, C.; Xu, W.; Anderson, D.M.; Li, Y.; Song, X.P.; Boyce, D.G.; Gibson, L.; et al. Coastal phytoplankton blooms expand and intensify in the 21st century. Nature 2023, 615, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lotze, H.K.; Worm, B.; Sommer, U. Propagule banks, herbivory and nutrient supply control population development and dominance patterns in macroalgal blooms. Oikos 2000, 89, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Cosper, E.M.; Dennison, W.C.; Carpenter, E.J.; Bricelj, V.M.; Mitchell, J.G.; Kuenstner, S.H.; Colflesh, D.; Dewey, M. Recurrent and persistent brown tide blooms perturb coastal marine ecosystem. Estuaries 1987, 10, 284–290. [Google Scholar] [CrossRef]
- Butler, M.J., IV; Hunt, J.H.; Herrnkind, W.F.; Childress, M.J.; Bertelsen, R.; Sharp, W.; Matthews, T.; Field, J.M.; Marshall, H.G. Cascading disturbances in Florida Bay, USA: Cyanobacteria blooms, sponge mortality, and implications for juvenile spiny lobsters Panulirus argus. Mar. Ecol. Prog. Ser. 1995, 129, 119–125. [Google Scholar] [CrossRef]
- Frieder, C.A.; Kessouri, F.; Ho, M.; Sutula, M.; Bianchi, D.; McWilliams, J.C.; Deutsch, C.; Howard, E. Effects of urban eutrophication on pelagic habitat capacity in the Southern California Bight. ESS Open Arch. 2023. Available online: https://d197for5662m48.cloudfront.net/documents/publicationstatus/130200/preprint_pdf/cca98c7aa83c14dc943466ce32711731.pdf (accessed on 15 March 2023).
- Inomura, K.; Deutsch, C.; Jahn, O.; Dutkiewicz, S.; Follows, M.J. Global patterns in marine organic matter stoichiometry driven by phytoplankton ecophysiology. Nat. Geosci. 2022, 15, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, P.K.; Foster, S.D.; King, E.; Risbey, J.; O’Kane, T.J.; Monselesan, D.; Hobday, A.J.; Hartog, J.R.; Thompson, P.A. Global patterns of change and variation in sea surface temperature and chlorophyll a. Sci. Rep. 2018, 8, 14624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivak, A.C.; Sanderman, J.; Bowen, J.L.; Canuel, E.A.; Hopkinson, C.S. Global-change controls on soil-carbon accumulation and loss in coastal vegetated ecosystems. Nat. Geosci. 2019, 12, 685–692. [Google Scholar] [CrossRef]
- Gobler, C.J. Climate change and harmful algal blooms: Insights and perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef] [PubMed]
- Burd, A.B.; Jackson, G.A. An analysis of water column distributions in Florida Bay. Estuaries 2002, 25, 570–585. [Google Scholar] [CrossRef]
- Briceño, H.O.; Boyer, J.N. Climatic controls on phytoplankton biomass in a sub-tropical estuary, Florida Bay, USA. Estuaries Coasts 2010, 33, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Nelson, N.G.; Munoz-Carpena, R.; Phlips, E.J. A novel quantile method reveals spatiotemporal shifts in phytoplankton biomass descriptors between bloom and non-bloom conditions in a subtropical estuary. Mar. Ecol. Prog. Ser. 2017, 567, 57–78. [Google Scholar] [CrossRef]
- Galbraith, E.; Convertino, M. The Eco-Evo Mandala: Simplifying Bacterioplankton Complexity into Ecohealth Signatures. Entropy 2021, 23, 1471. [Google Scholar] [CrossRef]
- Malone, T.C.; Newton, A. The globalization of cultural eutrophication in the coastal ocean: Causes and consequences. Front. Mar. Sci. 2020, 7, 670. [Google Scholar] [CrossRef]
- Galbraith, E.; Frade, P.; Convertino, M. Metabolic shifts of oceans: Summoning bacterial interactions. Ecol. Ind. 2022, 138, 108871. [Google Scholar] [CrossRef]
- Wang, X.F. Complex networks: Topology, dynamics and synchronization. Int. J. Bifurc. Chaos 2002, 12, 885–916. [Google Scholar] [CrossRef]
- Ilany, A.; Booms, A.S.; Holekamp, K.E. Topological effects of network structure on long-term social network dynamics in a wild mammal. Ecol. Lett. 2015, 18, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Convertino, M. Inferring ecosystem networks as information flows. Sci. Rep. 2021, 11, 1–22. [Google Scholar] [CrossRef]
- Reijneveld, J.C.; Ponten, S.C.; Berendse, H.W.; Stam, C.J. The application of graph theoretical analysis to complex networks in the brain. Clin. Neurophysiol. 2007, 118, 2317–2331. [Google Scholar] [CrossRef] [PubMed]
- Donges, J.F.; Zou, Y.; Marwan, N.; Kurths, J. Complex networks in climate dynamics. Eur. Phys. J. Spec. Top. 2009, 174, 157–179. [Google Scholar] [CrossRef] [Green Version]
- Coscia, M.; Giannotti, F.; Pedreschi, D. A classification for community discovery methods in complex networks. Stat. Anal. Data Min. ASA Data Sci. J. 2011, 4, 512–546. [Google Scholar] [CrossRef] [Green Version]
- Feldhoff, J.H.; Lange, S.; Volkholz, J.; Donges, J.F.; Kurths, J.; Gerstengarbe, F.W. Complex networks for climate model evaluation with application to statistical versus dynamical modeling of South American climate. Clim. Dyn. 2015, 44, 1567–1581. [Google Scholar] [CrossRef]
- Li, J.; Convertino, M. Optimal microbiome networks: Macroecology and criticality. Entropy 2019, 21, 506. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Donner, R.V.; Marwan, N.; Donges, J.F.; Kurths, J. Complex network approaches to nonlinear time series analysis. Phys. Rep. 2019, 787, 1–97. [Google Scholar] [CrossRef]
- Zhang, J.; Small, M. Complex network from pseudoperiodic time series: Topology versus dynamics. Phys. Rev. Lett. 2006, 96, 238701. [Google Scholar] [CrossRef] [Green Version]
- Gfeller, D.; De Los Rios, P.; Caflisch, A.; Rao, F. Complex network analysis of free-energy landscapes. Proc. Natl. Acad. Sci. USA 2007, 104, 1817–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Ban, Y. Complex network topology of transportation systems. Transp. Rev. 2013, 33, 658–685. [Google Scholar] [CrossRef]
- Zhou, C.; Zemanová, L.; Zamora, G.; Hilgetag, C.C.; Kurths, J. Hierarchical organization unveiled by functional connectivity in complex brain networks. Phys. Rev. Lett. 2006, 97, 238103. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Zemanová, L.; Zamora-Lopez, G.; Hilgetag, C.C.; Kurths, J. Structure–function relationship in complex brain networks expressed by hierarchical synchronization. N. J. Phys. 2007, 9, 178. [Google Scholar] [CrossRef]
- Albert, R.; Barabási, A.L. Statistical mechanics of complex networks. Rev. Modern Phys. 2002, 74, 47. [Google Scholar] [CrossRef] [Green Version]
- Boccaletti, S.; Latora, V.; Moreno, Y.; Chavez, M.; Hwang, D.U. Complex networks: Structure and dynamics. Phys. Rep. 2006, 424, 175–308. [Google Scholar] [CrossRef]
- Costa, L.d.F.; Rodrigues, F.A.; Travieso, G.; Villas Boas, P.R. Characterization of complex networks: A survey of measurements. Adv. Phys. 2007, 56, 167–242. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E. The structure and function of complex networks. SIAM Rev. 2003, 45, 167–256. [Google Scholar] [CrossRef] [Green Version]
- Andrade Jr, J.S.; Herrmann, H.J.; Andrade, R.F.; Da Silva, L.R. Apollonian networks: Simultaneously scale-free, small world, Euclidean, space filling, and with matching graphs. Phys. Rev. Lett. 2005, 94, 018702. [Google Scholar] [CrossRef] [Green Version]
- Dorogovtsev, S.N.; Goltsev, A.V.; Mendes, J.F.F. Pseudofractal scale-free web. Phys. Rev. E 2002, 65, 066122. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Raghavan, P.; Rajagopalan, S.; Sivakumar, D.; Tomkins, A.; Upfal, E. Stochastic models for the web graph. In Proceedings of the 41st Annual Symposium on Foundations of Computer Science, Redondo Beach, CA, USA, 12–14 November 2000; pp. 57–65. [Google Scholar]
- Hidalgo, J.; Grilli, J.; Suweis, S.; Munoz, M.A.; Banavar, J.R.; Maritan, A. Information-based fitness and the emergence of criticality in living systems. Proc. Natl. Acad. Sci. USA 2014, 111, 10095–10100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Carrillo, E.; López-Corona, O.; Toledo-Roy, J.C.; Lovett, J.C.; de León-González, F.; Osorio-Olvera, L.; Equihua, J.; Robredo, E.; Frank, A.; Dirzo, R.; et al. Assessing sustainability in North America’s ecosystems using criticality and information theory. PLoS ONE 2018, 13, e0200382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Convertino, M.; Valverde Jr, L.J. Toward a pluralistic conception of resilience. Ecol. Indic. 2019, 107, 105510. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Convertino, M. Temperature increase drives critical slowing down of fish ecosystems. PLoS ONE 2021, 16, e0246222. [Google Scholar] [CrossRef] [PubMed]
- Convertino, M.; Reddy, A.; Liu, Y.; Munoz-Zanzi, C. Eco-epidemiological scaling of leptospirosis: Vulnerability mapping and early warning forecasts. Sci. Total Environ. 2021, 799, 149102. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Convertino, M. Algal Bloom Ties: Systemic Biogeochemical Stress and Chlorophyll-a Shift Forecasting. Preprints 2023. submitted. [Google Scholar]
- Boyer, J.N.; Kelble, C.R.; Ortner, P.B.; Rudnick, D.T. Phytoplankton bloom status: Chlorophyll a biomass as an indicator of water quality condition in the southern estuaries of Florida, USA. Ecol. Ind. 2009, 9, S56–S67. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Lynch, T.C. Blooms of the picoplanktonic cyanobacterium Synechococcus in Florida Bay, a subtropical inner-shelf lagoon. Limnol. Oceanogr. 1999, 44, 1166–1175. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.N.; Briceño, H.O. South Florida Coastal Water Quality Monitoring Network. FY2006 Cumulative Report South Florida Water Management District, Southeast Environmental Research Center, Florida International University. 2007. Available online: http://serc.fiu.edu/wqmnetwork/ (accessed on 15 January 2023).
- Cloern, J.E.; Jassby, A.D. Patterns and scales of phytoplankton variability in estuarine–coastal ecosystems. Estuaries Coasts 2010, 33, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, T. Measuring information transfer. Phys. Rev. Lett. 2000, 85, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Convertino, M.; Wang, H. Envirome Disorganization and Ecological Riskscapes: The Algal Bloom Epitome. In Risk Assessment for Environmental Health; CRC Press: Boca Raton, FL, USA, 2022; pp. 327–346. [Google Scholar]
- Duan, Z.; Xu, H.; Huang, Y.; Feng, J.; Wang, Y. Multivariate time series forecasting with transfer entropy graph. Tsinghua Sci. Technol. 2022, 28, 141–149. [Google Scholar] [CrossRef]
- Sanders, R. The Pareto principle: Its use and abuse. J. Serv. Mark. 1987, 1, 37–40. [Google Scholar] [CrossRef]
- Anderson, S.C.; Cooper, A.B.; Dulvy, N.K. Ecological prophets: Quantifying metapopulation portfolio effects. Methods Ecol. Evol. 2013, 4, 971–981. [Google Scholar] [CrossRef]
- Martinello, M.; Hidalgo, J.; Maritan, A.; Di Santo, S.; Plenz, D.; Muñoz, M.A. Neutral theory and scale-free neural dynamics. Phys. Rev. X 2017, 7, 041071. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Patterson, D.; Levin, S.A.; Wang, J. Non-equilibrium early-warning signals for critical transitions in ecological systems. Proc. Natl. Acad. Sci. USA 2023, 120, e2218663120. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Galbraith, E.; Convertino, M. Algal Bloom Ties: Spreading Network Inference and Extreme Eco-Environmental Feedback. Entropy 2023, 25, 636. https://doi.org/10.3390/e25040636
Wang H, Galbraith E, Convertino M. Algal Bloom Ties: Spreading Network Inference and Extreme Eco-Environmental Feedback. Entropy. 2023; 25(4):636. https://doi.org/10.3390/e25040636
Chicago/Turabian StyleWang, Haojiong, Elroy Galbraith, and Matteo Convertino. 2023. "Algal Bloom Ties: Spreading Network Inference and Extreme Eco-Environmental Feedback" Entropy 25, no. 4: 636. https://doi.org/10.3390/e25040636
APA StyleWang, H., Galbraith, E., & Convertino, M. (2023). Algal Bloom Ties: Spreading Network Inference and Extreme Eco-Environmental Feedback. Entropy, 25(4), 636. https://doi.org/10.3390/e25040636