

Gear Shifting in Biological Energy Transduction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Variation of the Force Ratio May Induce Catabolic Gear Shifting
2.2. Gear Shifting Simulations
2.3. Simulation of ATP Synthesis Flux through a Dual Pathway
2.4. Discontinuous Optimal Gear Shifting
2.5. Reproducibility and Accessibility of the Data
3. Results
3.1. Mosaic Non-Equilibrium Thermodynamics and How the Variation of the Force Ratio Induces Gear Shifting of Catabolism
3.2. Phenomenological Non-Equilibrium Thermodynamics: Gear Shifting Affects the Phenomenological Stoichiometry as Well as the Degree of Coupling
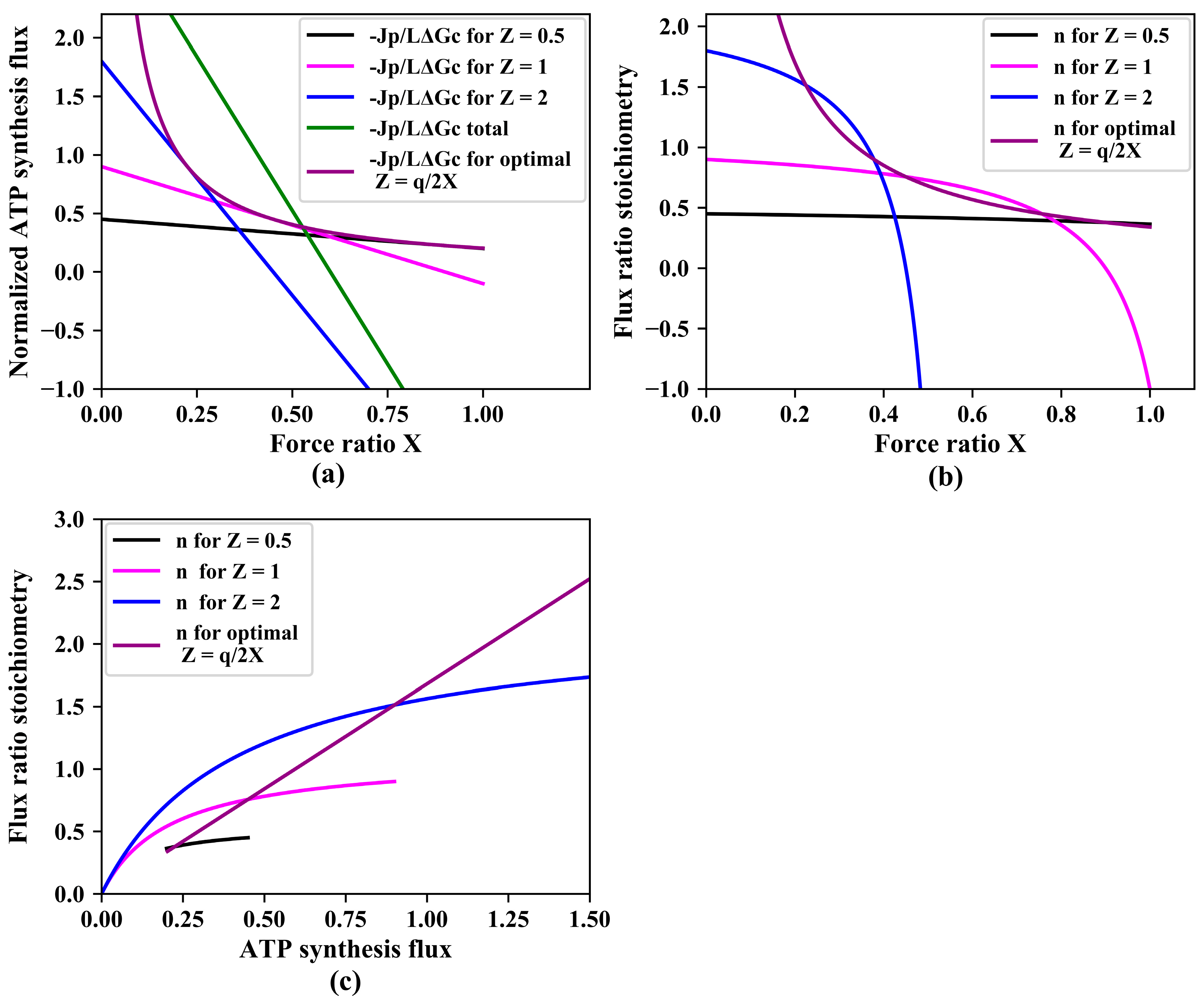
3.3. Variations in the Phenomenological Stoichiometry Could Improve ATP Synthesis and Growth
3.4. Gear Shifting and Varied Relative Pathway Capacities Could Improve ATP Synthesis and Growth
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pitzer, K.S. Thermodynamics, 3rd ed.; McGraw-Hil: New York, NY, USA, 1995. [Google Scholar]
- Hill, T.L. An Introduction to Statistical Thermodynamics; Dover Publications Inc.: New York, NY, USA, 1986. [Google Scholar]
- de Groot, S.R.; Mazur, P. Non-Equilibrium Thermodynamics; Dover Publications Inc.: New York, NY, USA, 1984. [Google Scholar]
- Glansdorff, P.; Nicolis, G.; Prigogine, I. The Thermodynamic Stability Theory of Non-Equilibrium States. Proc. Natl. Acad. Sci. USA 1974, 71, 197–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keizer, J. Statistical Thermodynamics of Nonequilibrium Processes; Springer: New York, NY, USA, 1987. [Google Scholar]
- Mikulecky, D.C. Network Thermodynamics and Complexity: A Transition to Relational Systems Theory. Comput. Chem. 2001, 25, 369–391. [Google Scholar] [CrossRef] [PubMed]
- Glansdorff, P.; Prigogine, I. Thermodynamic Theory of Structure, Stability and Fluctuations; Wiley: London, UK, 1971. [Google Scholar]
- Katchalsky, A.; Curran, P.F. Nonequilibrium Thermodynamics in Biophysics; Harvard University Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Onsager, L. Reciprocal Relations in Irreversible Processes. I. Phys. Rev. 1931, 37, 405–426. [Google Scholar] [CrossRef] [Green Version]
- Westerhoff, H.V.; Hellingwerf, K.J.; Arents, J.C.; Scholte, B.J.; Van Dam, K. Mosaic Nonequilibrium Thermodynamics Describes Biological Energy Transduction. Proc. Natl. Acad. Sci. USA 1981, 78, 3554–3558. [Google Scholar] [CrossRef] [Green Version]
- Feinman, R.D.; Fine, E.J. Nonequilibrium Thermodynamics and Energy Efficiency in Weight Loss Diets. Theor. Biol. Med. Model. 2007, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Mondeel, T.D.G.A.; Astrologo, S.; Zhang, Y.; Westerhoff, H.V. NET Works after All? Engineering Robustness through Diversity. IFAC-Pap. 2018, 51, 128–137. [Google Scholar] [CrossRef]
- Hernández-Lemus, E. Nonequilibrium Thermodynamics of Cell Signaling. J. Thermodyn. 2012, 1, 432143. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Barak, R.; Liarzi, O.; Shainskaya, A.; Eisenbach, M. In Vivo Acetylation of CheY, a Response Regulator in Chemotaxis of Escherichia Coli. J. Mol. Biol. 2008, 376, 1260–1271. [Google Scholar] [CrossRef]
- Popov, E.M. Protein Folding as a Nonlinear Nonequilibrium Thermodynamic Process. Biochem. Mol. Biol. Int. 1999, 47, 443–453. [Google Scholar] [CrossRef]
- Kemp, M.; Go, Y.M.; Jones, D.P. Nonequilibrium Thermodynamics of Thiol/Disulfide Redox Systems: A Perspective on Redox Systems Biology. Free Radic. Biol. Med. 2008, 44, 921–937. [Google Scholar] [CrossRef] [Green Version]
- Grande-Garcia, I. The Evolution of Brain and Mind: A Non-Equilibrium Thermodynamics Approach. Ludus Vitalis 2007, XV, 103–125. [Google Scholar]
- Buxton, R.B. The Thermodynamics of Thinking: Connections between Neural Activity, Energy Metabolism and Blood Flow. Trans. R. Soc. Lond. B Biol. Sci. Philos. 2021, 376, 20190624. [Google Scholar] [CrossRef] [PubMed]
- Loge, R.E.; Suo, Z. Nonequilibrium Thermodynamics of Ferroelectric Domain Evolution. Acta Mater. 1996, 44, 3429–3438. [Google Scholar] [CrossRef]
- Dewey, T.G.; Donne, M.D. Non-Equilibrium Thermodynamics of Molecular Evolution. J. Theor. Biol. 1998, 193, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Mondeel, T.D.G.A.; Rehman, S.; Zhang, Y.; Verma, M.; Durre, P.; Barberis, M.; Westerhoff, H.V. Maps for When the Living Gets Tough: Maneuvering through a Hostile Energy Landscape. IFAC Pap. 2016, 49, 364–370. [Google Scholar] [CrossRef]
- Abudukelimu, A.; Mondeel, T.D.G.A.; Barberis, M.; Westerhoff, H.V. Learning to read and write in evolution: From static pseudoenzymes and pseudosignalers to dynamic gear shifters. Biochem Soc Trans. 2017 45, 635–652. [CrossRef]
- Otten, M.F.; Stork, D.M.; Reijnders, W.N.M.; Westerhoff, H.V.; Van Spanning, R.J.M. Regulation of Expression of Terminal Oxidases in Paracoccus Denitrificans. Eur. J. Biochem. 2001, 268, 2486–2497. [Google Scholar] [CrossRef]
- Bekker, M.; De Vries, S.; Ter Beek, A.; Hellingwerf, K.J.; Teixeira De Mattos, M.J. Respiration of Escherichia Coli Can Be Fully Uncoupled via the Nonelectrogenic Terminal Cytochrome Bd-II Oxidase. J. Bacteriol. 2009, 191, 5510–5517. [Google Scholar] [CrossRef] [Green Version]
- Quehenberger, J.; Shen, L.; Albers, S.V.; Siebers, B.; Spadiut, O. Sulfolobus—A Potential Key Organism in Future Biotechnology. Front. Microbiol. 2017, 8, 2474. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kouril, T.; Snoep, J.L.; Siebers, B.; Barberis, M.; Westerhoff, H.V. The Peculiar Glycolytic Pathway in Hyperthermophylic Archaea: Understanding Its Whims by Experimentation in Silico. Int. J. Mol. Sci. 2017, 18, 876. [Google Scholar] [CrossRef] [Green Version]
- Westerhoff, H.V.; Van Dam, K. Thermodynamics and Control of Biological Free-Energy Transduction; Elsevier: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Kedem, O.; Caplan, S.R. Degree of Coupling and Its Relation to Efficiency of Energy of Conversion. Trans. Faraday Soc. 1965, 21, 1897–1911. [Google Scholar] [CrossRef]
- Stucki, J.W. The Optimal Efficiency and the Economic Degrees of Coupling of Oxidative Phosphorylation. Eur. J. Biochem. 1980, 283, 269–283. [Google Scholar] [CrossRef]
- Rottenberg, H. The Thermodynamic Description of Enzyme-Catalyzed Reactions: The Linear Relation between the Reaction Rate and the Affinity. Biophys. J. 1973, 13, 503–511. [Google Scholar] [CrossRef] [Green Version]
- van der Meer, R.; Westerhoff, H.V.; Van Dam, K. Linear Relation between Rate and Thermodynamic Force in Enzyme-Catalyzed Reactions. Biochim. Biophys. Acta Bioenerg. 1980, 591, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Chemiosmotic Coupling in Oxidative and Photosynthetic Phosphorylation. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 1507–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerhoff, H.V.; Hellingwerf, K.; Van Dam, K. Thermodynamic Efficiency of Microbial Growth Is Low but Optimal for Maximal Growth Rate. Proc. Natl. Acad. Sci. USA 1983, 80, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.S.; Ibarra, R.U.; Palsson, B.O. In Silico Predictions of Escherichia Coli Metabolic Capabilities Are Consistent with Experimental Data. Nat. Biotechnol. 2001, 19, 125–130. [Google Scholar] [CrossRef]
- Arents, J.C.; van Dekken, H.; Hellingwerf, K.J.; Westerhoff, H.V. Linear Relations between Proton Current and pH Gradient in Bacteriorhodopsin Liposomes. Biochemistry 1981, 20, 5114–5123. [Google Scholar] [CrossRef] [PubMed]
- Herrgård, M.J.; Swainston, N.; Dobson, P.; Dunn, W.B.; Arga, K.Y.; Arvas, M. A Consensus Yeast Metabolic Network Reconstruction Obtained from a Community Approach to Systems Biology. Nat. Biotechnol. 2008, 26, 1155–1160. [Google Scholar] [CrossRef]
- Rohwer, J.M. Kinetic Modelling of Plant Metabolic Pathways. J. Exp. Bot. 2012, 63, 2275–2292. [Google Scholar] [CrossRef] [Green Version]
- Penkler, G.; Du Toit, F.; Adams, W.; Rautenbach, M.; Palm, D.C.; Van Niekerk, D.D.; Snoep, J.L. Construction and Validation of a Detailed Kinetic Model of Glycolysis in Plasmodium Falciparum. FEBS J. 2015, 282, 1481–1511. [Google Scholar] [CrossRef]
- Edwards, J.S.; Palsson, B.O. Metabolic Flux Balance Analysis and the in Silico Analysis of Escherichia Coli K-12 Gene Deletions. BMC Bioinform. 2000, 1, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Westerhoff, H.V. Competitive, Multi-Objective, and Compartmented Flux Balance Analysis for Addressing Tissue-Specific Inborn Errors of Metabolism. J. Inherit. Metab. Dis. 2023, 1–13. [Google Scholar] [CrossRef]
- Keizer, J.; Foxt, R.F. Qualms Regarding the Range of Validity of the Glansdorff-Prigogine Criterion for Stability of Non-Equilibrium States. Proc. Nat. Acad. Sci. USA 1974, 71, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortassa, S.; Aon, M.A.; Westerhoff, H.V. Linear Nonequilibrium Thermodynamics Describes the Dynamics of an Autocatalytic System. Biophys. J. 1991, 60, 794–803. [Google Scholar] [CrossRef] [Green Version]
- Goldbeter, A. Biochemical Oscillations and Cellular Rhythms; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Van Hoek, P.; Van Dijken, J.P.; Pronk, J.T. Effect of Specific Growth Rate on Fermentative Capacity of Baker’s Yeast. Appl. Environ. Microbiol. 1998, 64, 4226–4233. [Google Scholar] [CrossRef] [Green Version]
- Damiani, C.; Colombo, R.; Gaglio, D.; Mastroianni, F.; Pescini, D.; Westerhoff, H.V.; Mauri, G.; Vanoni, M.; Alberghina, L. A Metabolic Core Model Elucidates How Enhanced Utilization of Glucose and Glutamine, with Enhanced Glutamine- Dependent Lactate Production, Promotes Cancer Cell Growth: The WarburQ Effect. PLoS Comput. Biol. 2017, 13, e1005758. [Google Scholar] [CrossRef] [Green Version]
- Stouthamer, A.H. Yield Studies in Microorganisms; Meadowfield Press Limited: Durham, UK, 1976. [Google Scholar]
- Grigaitis, P.; Teusink, B. An Excess of Glycolytic Enzymes under Glucose-Limited Conditions May Enable Saccharomyces Cerevisiae to Adapt to Nutrient Availability. FEBS Lett. 2022, 596, 3203–3210. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Westerhoff, H.V. Gear Shifting in Biological Energy Transduction. Entropy 2023, 25, 993. https://doi.org/10.3390/e25070993
Zhang Y, Westerhoff HV. Gear Shifting in Biological Energy Transduction. Entropy. 2023; 25(7):993. https://doi.org/10.3390/e25070993
Chicago/Turabian StyleZhang, Yanfei, and Hans V. Westerhoff. 2023. "Gear Shifting in Biological Energy Transduction" Entropy 25, no. 7: 993. https://doi.org/10.3390/e25070993
APA StyleZhang, Y., & Westerhoff, H. V. (2023). Gear Shifting in Biological Energy Transduction. Entropy, 25(7), 993. https://doi.org/10.3390/e25070993