1. Introduction
The sudden or recurrent emergence of epidemics has been an everlasting threat to humanity. Highly infectious and fatal diseases such as pestilence, typhus, cholera, and leprosy were among the main causes of death in medieval times in Europe and, until the 20th century, a major scourge of humanity [
1]. This permanent challenge has naturally driven interest in protective measures and predictive models.
The systematic mathematical study of epidemic spreading began only a century ago with the seminal work of Kermack and McKendrick [
2]. They were the first to introduce what we call currently a “compartment model”. In their so-called SIR model, the individuals are categorized into the compartments susceptible (S), infected (I), and recovered (immune) (R), characterizing the states of their health. While standard SIR-type models are able to capture the main features of a certain class of infectious diseases such as mumps, measles, and rubella, they fail to describe persistent oscillatory behaviors and spontaneous outbursts, which are observed in many epidemics.
A large amount of work still is devoted to compartmental models [
3,
4], where an impressive field has emerged [
5,
6], and the interest was again considerably enhanced by the context of the COVID-19 pandemic [
7]. In addition to purely macroscopic models, the study of epidemic dynamics in complex networks has attracted considerable attention [
8,
9,
10]. In these works, the importance of the graph topology for spreading phenomena has been highlighted. In particular, Pastor-Satorras and Vespignani showed that, for a wide range of scale-free networks, no critical threshold for epidemic spreading exists [
9]. The topological features crucial for epidemic spreading include the small world property (short average network distances (the network “distance” of two nodes is the number of edges of the shortest path connecting them) and a high clustering coefficient, measuring the existence of redundant paths between pairs of nodes [
11,
12].
Further interesting directions are represented by combinations of network science and stochastic compartmental models [
13,
14,
15,
16,
17]. Such models include Markovian and non-Markovian approaches [
18,
19,
20,
21,
22], where non-Markovianity is introduced by non-exponentially distributed sojourn times in the compartments [
23,
24]. In these works, explicit formulae for the endemic equilibrium in terms of mean compartmental sojourn times and the basic reproduction number are derived and numerically validated in random walk simulations. A further non-Markovian model appeared recently [
25], where non-Markovianity comes into play by introducing an “age of infection”, allowing individuals to recover when their infection period exceeds a certain threshold, generalizing the initial idea of Kermack and McKendrick.
Other works emphasize the importance of the spatial heterogeneity effects of infection patterns in epidemic spreading phenomena [
26]. The role of local clusters in generating periodic epidemic outbursts has been highlighted in the references [
27,
28]. A cluster model to explain periodic behavior was introduced a long time ago [
29]. The role of the complex interplay of retardation (delayed compartmental transitions) and fluctuations for oscillatory behavior has been investigated in one of our recent works [
22].
The aim of the present paper is to study the spreading of a certain class of vector-transmitted diseases in a population of individuals (random walkers) moving on complex graphs aiming to mimic human mobility patterns in complex environments such as cities, streets, and transportation networks. Essential elements in our model are the accounting of the mortality of infected individuals (random walkers) and an indirect transmission pathway via vectors (nodes of the network). The random walkers are mimicking individuals that are navigating on the network. The nodes of the network are assumed to mimic the vectors (for instance, the mosquitoes in the case of Malaria). The reason why we chose the nodes of the network to mimic the vectors is based on the observation that, in real-world situations, the vectors live in stationary well-defined areas such as swamps and others. The nodes are immortal, since we assume that the vectors (mosquitoes) are not falling ill during their infection time span.
The present paper is organized as follows. In
Section 2, we establish a stochastic mean-field model for the evolution of the compartmental populations. The special case of zero mortality is considered in
Section 3, where we obtain an explicit formula for the endemic equilibrium (stationary constant compartmental populations for infinite time). In this way, we identify a crucial parameter controlling the stability of the healthy state having the interpretation of the basic reproduction number
(
Section 4), where the healthy state is stable for
and unstable for
. A detailed proof of the stability of the endemic state for
is provided in
Appendix A.2. In
Section 5, we analyze the stability of the healthy state with mortality, derive the basic reproduction number
, and prove that
, i.e., mortality reduces the basic reproduction number. In
Section 6, we test the robustness of our mean-field model under “complex real-world conditions” by implementing its assumptions in multiple random random walker simulations on Barabási–Albert (BA)-, Erdös–Rényi (ER)-, and Watts–Strogatz (WS)-type graphs (see [
14,
15,
30] and
Appendix A.4). These graph types have different complexity and connectivity features with an impact on the spreading.
2. Compartmental Model with Mortality
The goal of this section is to develop a mean-field model for a certain class of diseases with indirect infection transmission via vectors, which includes Dengue, Malaria (transmission by mosquitoes), pestilence (transmission by fleas), and others [
8,
31]. To that end, we consider a population of
Z random walkers navigating independently on a connected (ergodic) graph. Each walker performs independent steps from one to another connected node on the network (specified subsequently). We assume that walkers and nodes are in one of the compartments, S (susceptible) and I (infected). In addition, walkers can be in compartment D (dead), whereas nodes never die.
Let
(
) be the number of walkers (nodes) in compartments S and I and
the non-decreasing number of walkers (in compartment D) that died from the disease up to time
t. We consider
walkers (
Z independent of time) and a constant number of nodes
. We assume at instant
the first spontaneous occurrence of the disease of a few infected walkers
or nodes
(and no dead walkers
). We introduce the compartmental fractions
,
,
for the walkers (normalized with respect to
Z) with
and
,
with
. To limit the complexity of our model, we do not consider the demographic evolution, i.e., there are no natural birth and death processes. We denote with
the infection rates (rates of transitions S → I) of walkers and nodes, respectively. We assume the following simple bi-linear forms:
with constant rate parameters
(independent of time).
indicates the infection rate of walkers, where its dependence on
is telling us that susceptible walkers can be infected only by (visiting) infected nodes.
stands for the infection rate of nodes depending on
indicating that susceptible nodes can only be infected by (visits of) infected walkers. There are no direct transmissions among walkers and among nodes. Infections of walkers (nodes) take place with specific transmission probabilities in the contact of a node and a walker, which are captured by (yet not identical to) the transmission rate constants
.
The infection time spans
without mortality (waiting times in compartment I) of walkers and nodes are assumed to be independent random variables drawn from specific distributions specified hereafter. As the only admitted death process, we assume that infected walkers may die within the time span of their infection. To capture this kind of mortality caused by the disease, we introduce a further independent random variable
, which indicates the life span of an infected walker. Both the infection and life spans
are counted from the time instant of the infection. A walker survives the disease if
and dies from it for
. With these assumptions, we first give a stochastic formulation of the evolution equations:
where
indicates the (non-negative) mortality rate of walkers. We indicate with
the average over the contained (set of independent) random variables
outlined hereafter and in
Appendix A.1.
stands for the Heaviside function (
A2), and
for Dirac’s
-distribution. An epidemic always starts from “natural” initial conditions
,
(globally healthy state), where, at
, the first infections occur spontaneously and can be “generated” by adding the source terms
to the infection rates of walkers and nodes, respectively. Equivalently, we introduce initial conditions
(
) with
consisting typically of a few infected walkers and/or nodes in a large susceptible population without dead walkers
.
The interpretation of the system (
2) is as follows. The instantaneous infection rate
governs the transitions S → I of walkers (due to the visits of infected nodes). The term
describes the rate of walkers recovering at time
t and infected at
, i.e., their infection time span has elapsed and they survived as
(indicated by
). Then,
captures the rate of walkers infected at
dying at time
t during the infection time span (indicated by
for
).
Remark 1. The infection time span of a walker (sojourn time in compartment I) is , i.e., if (where the walker survives the disease), and is if the walker dies within the infectious time span (). is the walker’s infection time span without mortality (retrieved for ). The probability of the persistence of a walker’s infection at time t, given the infection starts at time 0, is (see (7)). Note that only if , i.e., when the walker is in compartment I. As a crucial element of our model, we will analyze the statistics of the walker’s infection time span . The initially infected walkers and nodes are as well subjected to the transition pathways, i.e., walkers either recover (alive) with rate or they die with rate , and nodes always recover with rate . For , these terms are evanescent; thus, the initial conditions do not affect large time limits (endemic state for zero mortality). The importance of these terms can be seen by setting (no infections). Without these terms, the initially infected walkers and nodes would stay infected forever, inconsistent with our assumptions.
The rate equations for the nodes can be interpreted in the same way as the interplay of instantaneous infections and delayed recovery without mortality. We emphasize that the evolution equations of the nodes and walkers are non-linearly coupled by the implicit dependencies of the infection rates (
1). In order to derive an explicit representation of the system (
2), we need to take a closer look at the averaging procedures and the involved distributions related to the independent random variables
drawn from specific probability density functions (PDFs), which we define by
with their respective PDFs (kernels)
, which are normalized
. Then, recall the averaging rule for the (suitable) functions
of the random variable
T, which we use throughout the paper:
see also
Appendix A.1. An important case is
. Then, by applying (
4), we introduce the persistence probabilities of the walker’s (node’s) infection (without mortality):
and the probability of the walker’s survival up to time
t (given
):
The persistence probabilities fulfill the initial condition
corresponding to the normalization of the waiting time PDFs
and are vanishing at infinity
. To evaluate the averages in (
2), we will use the following quantities:
In these averages, we make use of the independence of the waiting times
, and of the causality of
and the kernels
(i.e.,
for
). Of utmost importance are the “defective” PDFs (DPDFs)
of death and recovery. “Defective” means that
are not proper PDFs since they are not normalized to one, but rather to
, respectively. Consult [
32] for a recent account of defective distributions and related stochastic processes. They have the following interpretation.
is the probability of transition I → D within
of an infected walker (infected at
).
is the probability of transition I → S within
of a walker infected at
. Therefore,
is non-negative (as are
,
) and is a proper well-normalized PDF of all exits of walkers from compartment I (i.e., I → S + I → D). Without mortality (
), this PDF retrieves
.
The quantities
introduced in (
7) have the following interpretation.
is the probability that a walker infected at instant 0 is at time
t in compartment S (i.e., recovered prior or up to time
t).
is the probability that a walker infected at instant 0 is at time
t in compartment D (i.e., died prior and up to time
t). The infinite time limits are important:
has the interpretation of the overall probability that an infected walker survives the infection, and
is the overall probability for an infected walker to die from the disease. We refer to
also as “overall mortality”. It must not be confused with the infinite time limit of the dead walker’s fraction
, which is different from
, as we will see in detail subsequently. A small value
may cause a high value of
, for instance, for short infectious periods where walkers may be repeatedly infected.
In most cases, not all infected walkers die from their disease (in an infinite observation time); hence, (as is defective). represents the limit of a fatal disease and a disease without mortality. (as is defective) is the complementary probability with .
With these remarks, the system (
2) reads
The PDF (
8) for which a walker leaves compartment I (either by recovery or by death) allows rewriting the second equation of (
9) as
Worthy of closer consideration is the mortality rate of the infected walkers (representing the total mortality; entry rate into the D compartment):
where, clearly,
. Integrating this relation yields the fraction
of dead walkers:
An interesting quantity is the cumulative recovery rate of walkers (integrated entry rates of walkers into the S compartment; see the first equation in (
2)):
The quantity
records all recovery events of walkers up to time
t, where individual walkers may suffer repeated infections and recoveries. We observe that (see (
7))
Relation (
12) records all death events of walkers up to time
t. Since each walker may die only once, it follows indeed that
. Contrarily, the quantity
is not restricted to this interval as walkers may be repeatedly infected and recovered, but due to mortality, eventually only a finite number of times (
; see (
18)). Mortality renders the chain of infection and recovery events transient (due to the defective feature of
). To shed more light on the behavior of
, consider for a moment zero mortality (
) and
: we then have
(shown in
Section 3); thus,
coming along with an infinite chain of recurrent infection and recovery events (as
turns into the proper non-defective PDF
).
Using (
7), we can rewrite (
2) in equivalent integral form:
and with (redundant) Equation (
12) for the fraction of dead walkers. (
15) is a self-consistent system since the infection rates are implicit functions of the unknown susceptible and infected population fractions, i.e.,
,
(see (
1)). Explore now the infinite time limit of (
15), where it is convenient to consider the Laplace-transformed equations. We introduce the Laplace transform (LT) (denoted with a hat) of a function
as
where
denotes the (suitably chosen) Laplace variable. In order to retrieve infinite time limits, we use the asymptotic feature:
Observing that the LT of
is
and
, where
(see (
7)), we arrive at
where
. In the same way, one obtains
Since
is non-zero, the asymptotic values
depend on the initial condition
and the infection (rate) history. This is no longer true for zero mortality (
) and considered in
Section 3. We define the overall probability
that a walker dies from (
survives) the disease:
consistent with our previous interpretation of
, and the ratio:
The quantities (
19) and (
20) depend only on the stochastic features of
and
. They are independent of the infection rates and initial conditions and, therefore, of the topological properties of the network. In addition, they also do not depend on the stochastic features of the node’s infection time span
.
2.1. Markovian (Memoryless) Case
Generally, the system (
9) contains the history of the process (memory), which makes the process non-Markovian. The only exception is when all waiting times are exponentially distributed, namely
,
, where
and
. Then, (
9) takes with (
15) the memoryless form:
Putting the left-hand sides to zero yields the stationary state:
Let us check whether this result is consistent with (
17). To this end, we integrate the second equation in (
21) knowing that
, leading to
thus,
. Plugging this relation into (
22) and accounting for
, we recover indeed the representation of the expression (
17).
For zero mortality
, one can straightforwardly obtain the constant endemic equilibrium values
by setting the left-hand side of (
21) to zero, leading to the subsequent Equation (
32) derived in
Section 3 for general waiting time distributions with finite means.
2.2. A Few More Words on Waiting Time Distributions
In our simulations, we assumed that the time spans
are independent random variables drawn from specific Gamma distributions. The advantage of using Gamma distributions is that they may realize a large variety of shapes; see
Figure 1 for a few examples. To generate Gamma-distributed random numbers, we employed the Python 3.10.8 random number generator (library
numpy.random). Recall the Gamma distribution:
where
is the so-called “shape parameter” and
the rate parameter (often, the term “scale parameter” is used,
) and
stands for the Gamma function. It is worthy of mention that the Gamma distribution for
(also referred to as the Erlang distribution) is the PDF of the sum of independent and identically exponentially distributed random variables. We also will subsequently use the LT of the Gamma PDF:
The Gamma PDF has finite mean
, and for
, the Gamma PDF is weakly singular at
and
recovers exponential PDFs. For
, the Gamma PDF is completely monotonic (CM) (see Appendix, (
A9), for a definition). For the range
, the Gamma PDF has a maximum at
and becomes narrower the larger
(while keeping its mean
fixed); specifically, we can generate sharp waiting times using the feature:
We also will subsequently use the persistence probability of the Gamma distribution (see the right frame of
Figure 1):
where
indicates the upper incomplete Gamma function with
. (
27) necessarily fulfills the initial condition
and is vanishing at infinity
.
3. Endemic Equilibrium for Zero Mortality
Here, we consider the large time asymptotics of the compartment populations without mortality (
), where the self-consistent system (
15) reads
The endemic state emerging in the large time asymptotics does not depend on the initial conditions
as
. For what follows, it is convenient to consider the LTs of (
28), which read
where
are the LTs of the persistence distributions and
, reflecting constant populations of walkers and nodes. In order to determine the endemic equilibrium, we assume that the mean infection time spans for the nodes and walkers exist:
thus, the admissible range of the Laplace variable is
(if chosen real). Now, using (
16), we obtain the endemic equilibrium from
, where the initial conditions are wiped out at infinity as
. Assuming that the infection rates are constant in the endemic equilibrium, we have
, (
) and arrive at
thus,
One can see that the globally healthy state
is also a solution of this equation. Besides that, only solutions
correspond to an endemic equilibrium. One obtains
for the endemic equilibrium, which is independent of the initial conditions
. It depends only on the phenomenological rate constants
and the mean infection time spans
. We point out that the endemic equilibrium (
33) has exchange symmetry
between walkers and nodes, reflecting this feature in the system (
2) of evolution equations without mortality. The endemic values
are within
, i.e., existing only if
. We further mention the useful relation
following straightforwardly from (
32), connecting
directly with the endemic state. We interpret
as the basic reproduction number (average number of new infections at
—nodes or walkers—due to one infected node or walker at
). That this is really the appropriate interpretation can be seen by the following somewhat rough consideration of the infection rates at
. Assume we have initially one single infected node
and no infected walkers
. The expected number of walkers infected by this first infected node during its infectious period
is
The number of nodes getting infected by these
infected walkers during their infectious time
is
Hence,
is the average number of infected nodes caused by the first infected node (with zero initially infected walkers). Due to the exchange symmetry (nodes ↔ walkers) of the infection rates, this argumentation remains true when we start with one infected walker and no infected nodes.
We infer that the globally healthy state is unstable for
, where the endemic equilibrium (
33) is a unique stable fixed point and attractor for all initial conditions
. We will confirm this in the next section by a linear stability analysis of the globally healthy state. The stability of the endemic state is demonstrated in the next section together with
Appendix A.2.
The limit (while are kept constant) is remarkable, where all walkers become infected , but not all nodes and vice versa.
We plot the endemic state in
Figure 2 versus
, where positive values for
occur only for
, which correspond to endemic states. Next, we perform a linear stability analysis of the endemic and healthy state, where we will indeed identify
as a crucial control parameter.
4. Stability Analysis of Endemic and Healthy State without Mortality
Here, we are interested in the condition of spreading for zero mortality, or equivalently, in the condition for which the globally healthy state (endemic state) is unstable (stable). To check the stability of the endemic fixed point
,
, we set
where
are “small” constant amplitudes. This ansatz accounts for the constant populations of nodes and walkers. Then, we have for the infection rates up to linear orders in the amplitudes:
Plugging these relations in our evolution Equation (
2) without mortality, omitting two redundant equations leads to the system:
where we have used
and the cases of
-distributed
are contained for
. We point out that, in the ansatz (
35), we relax causality, i.e., we admit
for
; thus,
The solvability of this matrix equation requires the determinant to vanish, leading to a transcendental characteristic equation for
:
Generally, a fixed point is unstable if solutions with the positive real part of
exist. Consider this first for the globally healthy state
for which Equation (
38) reads
where we notice that
are the LTs of the persistence probabilities of the infection time spans. Consider this equation for
, and take into account (
30); we arrive at
We observe that
for
. On the other hand, we have for
that
, and hence,
One can, hence, infer from the complete monotony of
and, therefore, of
(see
Appendix A.2, Equation (
A9), for a precise definition) that
(
); thus,
has one single positive zero only if
, i.e., for
, which, therefore, is the condition of the instability of the healthy state (spreading of the disease). Conversely, for
, the healthy state turns into a stable fixed point where there is no spreading of the disease. In particular, the healthy state is always unstable (
) if at least one of the mean infection time spans
. This is true for fat-tailed kernels scaling as
(
) for
. We consider such a distribution briefly in the subsequent section. We plot function
versus
for different
for Gamma-distributed
in
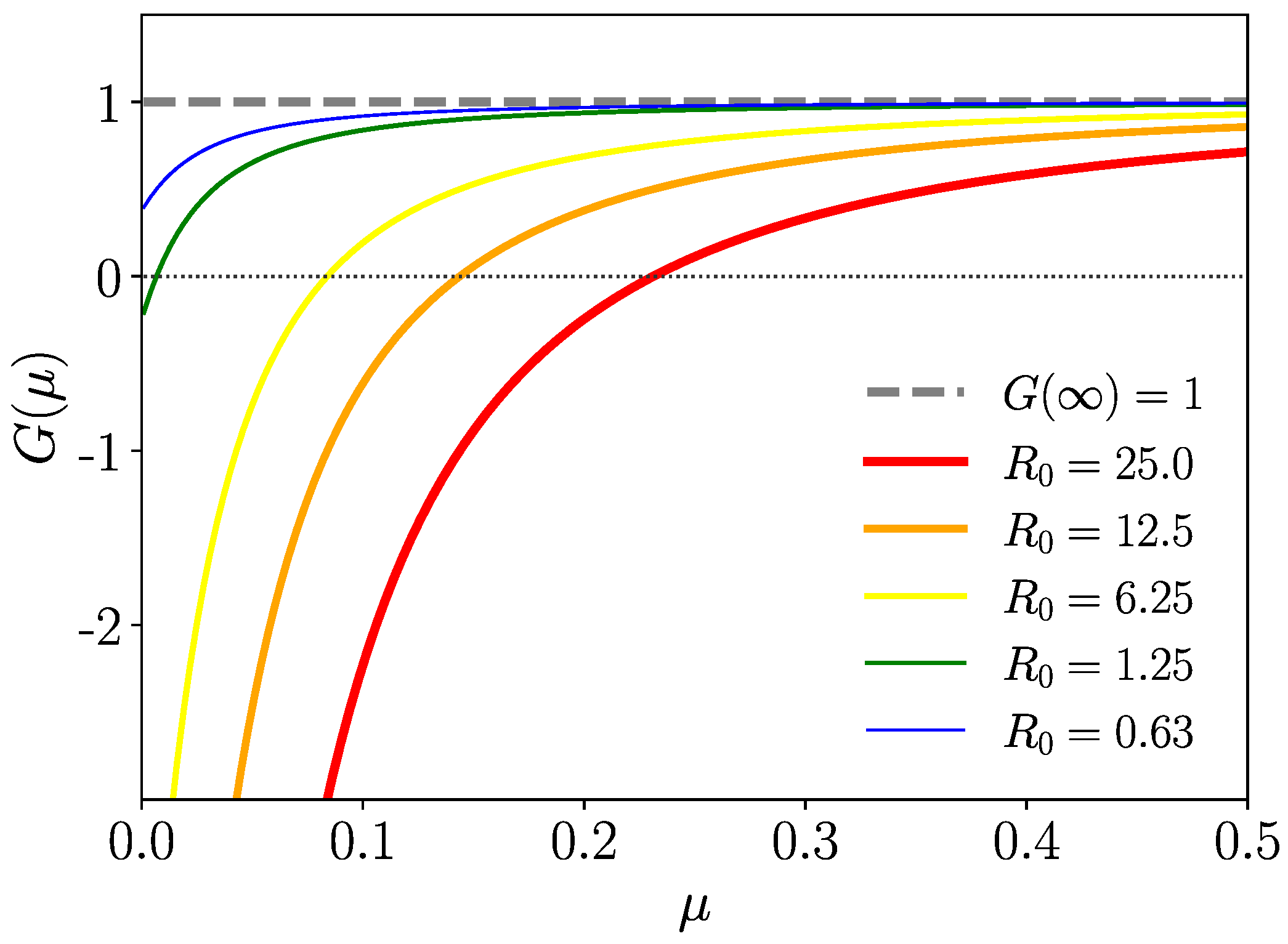
Figure 3.
Now, we consider the stability of the endemic state with
, where, from (
38), this function reads
with
On the other hand,
(as
), and from the monotony of
, it follows that there is no positive solution of
for
. We plot
in
Figure 4 for different values of
and Gamma-distributed
. In
Appendix A.2, we complete the analytical proof that
for
.
5. Stability Analysis of the Healthy State with Mortality
An important question is how mortality modifies the instability of the healthy state and the basic reproduction number. To shed light on this question, we perform a linear stability analysis of the healthy state
, where we set
with
and
. Plugging this ansatz for
into the three independent equations of (
2), say the first, third, and fourth one, and performing the averages (relaxing causality as previously), we arrive at
Putting the determinant of the matrix to zero yields the condition:
where the LTs
of the DPDFs
defined in (
7) come into play. We are interested in under which conditions there is a positive solution (instability of the healthy state) of (
46). Since
and
with
, we see that
is a solution of (
46). Recall from (
7) that
is the (properly normalized) PDF (
8). Condition (
46) then reads
where
is the LT of the persistence probability
of the walker’s infection, i.e., the probability that
(see Remark 1). For zero mortality, we have
, (
and
), retrieving the condition (
39). The mean sojourn time in compartment I with mortality yields
where we arrive at
Relation (
48) shows that
(equality only for zero mortality). On the other hand, we have
, so there is a positive solution of
only if
where
is the basic reproduction number modified by mortality with
(equality only for zero mortality). To visualize the effect of mortality on the instability of the healthy state, we plot
for a few values of
in
Figure 5. Increasing mortality turns an unstable healthy state into a stable one.
In the random walk simulations, we deal with Gamma-distributed
, where the persistence probabilities are then normalized incomplete Gamma functions (
27). To explore the effect of mortality for such cases, we determine
by numerical integration of (
48) as a function of the mortality rate parameter
and plot the result in
Figure 6, where one can see that
is monotonously decreasing with mortality rate
. We also include a case of a fat-tailed (Mittag–Leffler)-distributed
, which we discuss hereafter. The parameters in
Figure 6 are such that the zero mortality case occurs with
as the upper bound for the Gamma-distributed cases. The essential feature is that
decays monotonically with increasing mortality rate parameter
approaching zero for
. Diseases with high mortality stabilize the healthy state even for
.
Consider briefly the case where
is exponentially distributed (i.e.,
in the Gamma distribution of
) with
. Then, we have
; thus,
The zero mortality case is recovered for
with
. For Gamma-distributed
, this yields
where
,
are the parameters of the respective Gamma distributions of the infection times of nodes and walkers. The Markovian case where all waiting times are exponentially distributed is covered for
and yields
containing the zero mortality case for
.
Fat-Tailed-Distributed
Finally, an interesting case emerges if
follows a fat-tailed distribution, i.e.,
for
t large (
) and
,
. Let us take a look at how mortality is affecting this situation. Fat-tailed
distributions describe diseases where the infectious periods are very long and the healthy state without mortality is extremely unstable (
). Infected walkers can infect many nodes during their long infection time spans. An important case of this class is constituted by the Mittag–Leffler (ML) distribution
, where
indicates the Mittag–Leffler function; see [
33,
34] and the references therein for representations and connections with fractional calculus. The ML function recovers the exponential for
(
). Assuming exponential mortality
, one obtains with (
51)
containing the LT of the ML persistence probability distribution
. The essential feature here is that
is weakly singular at
with a monotonously decreasing
scaling law, where the healthy state becomes stable for mortality parameters larger as
. We depict this case in
Figure 6 for
(violet curve).



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












