Polymers for DNA Delivery

{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
Gene delivery systems
Viral delivery systems
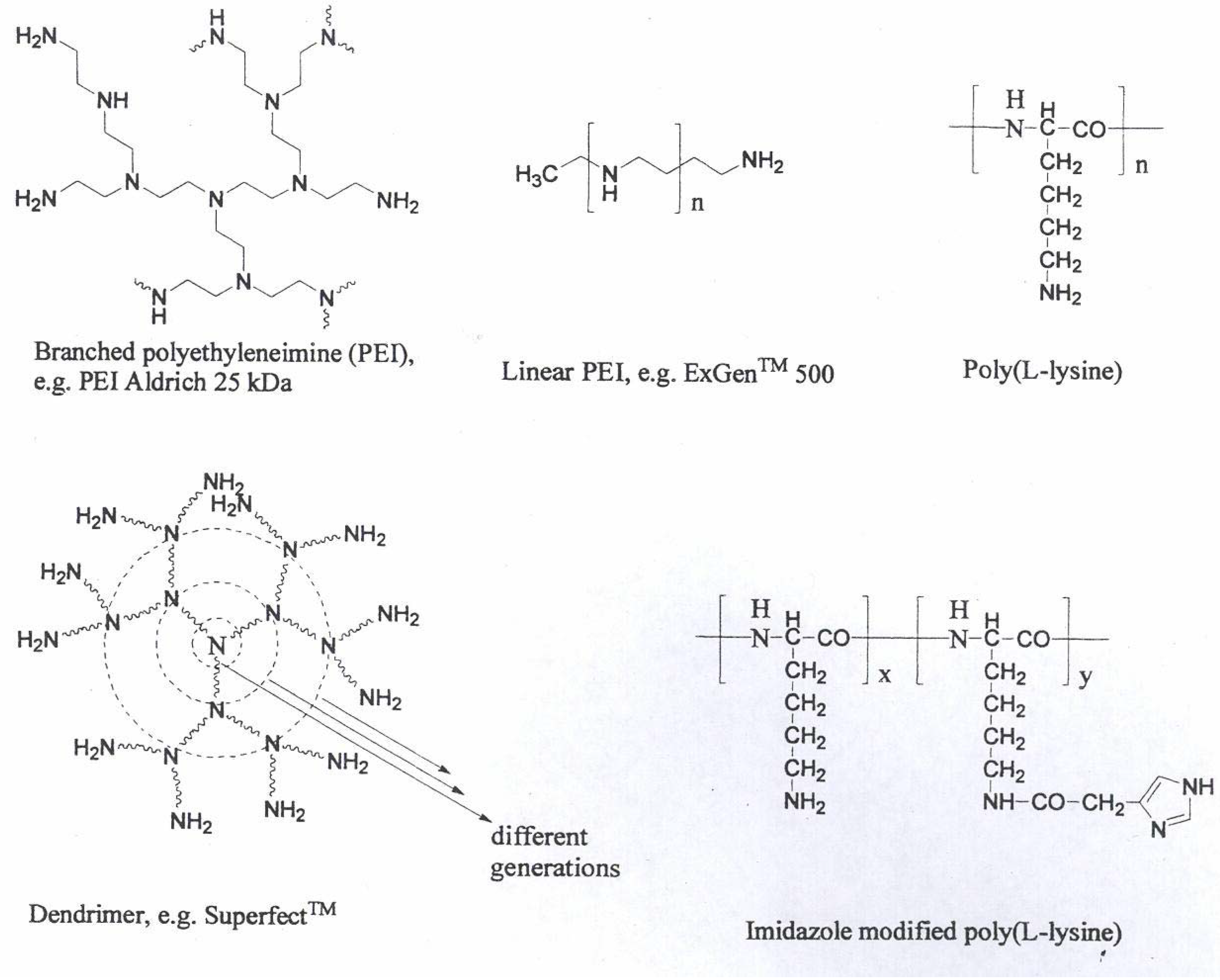
Cationic polymers
PEI

PLL
Dendrimers
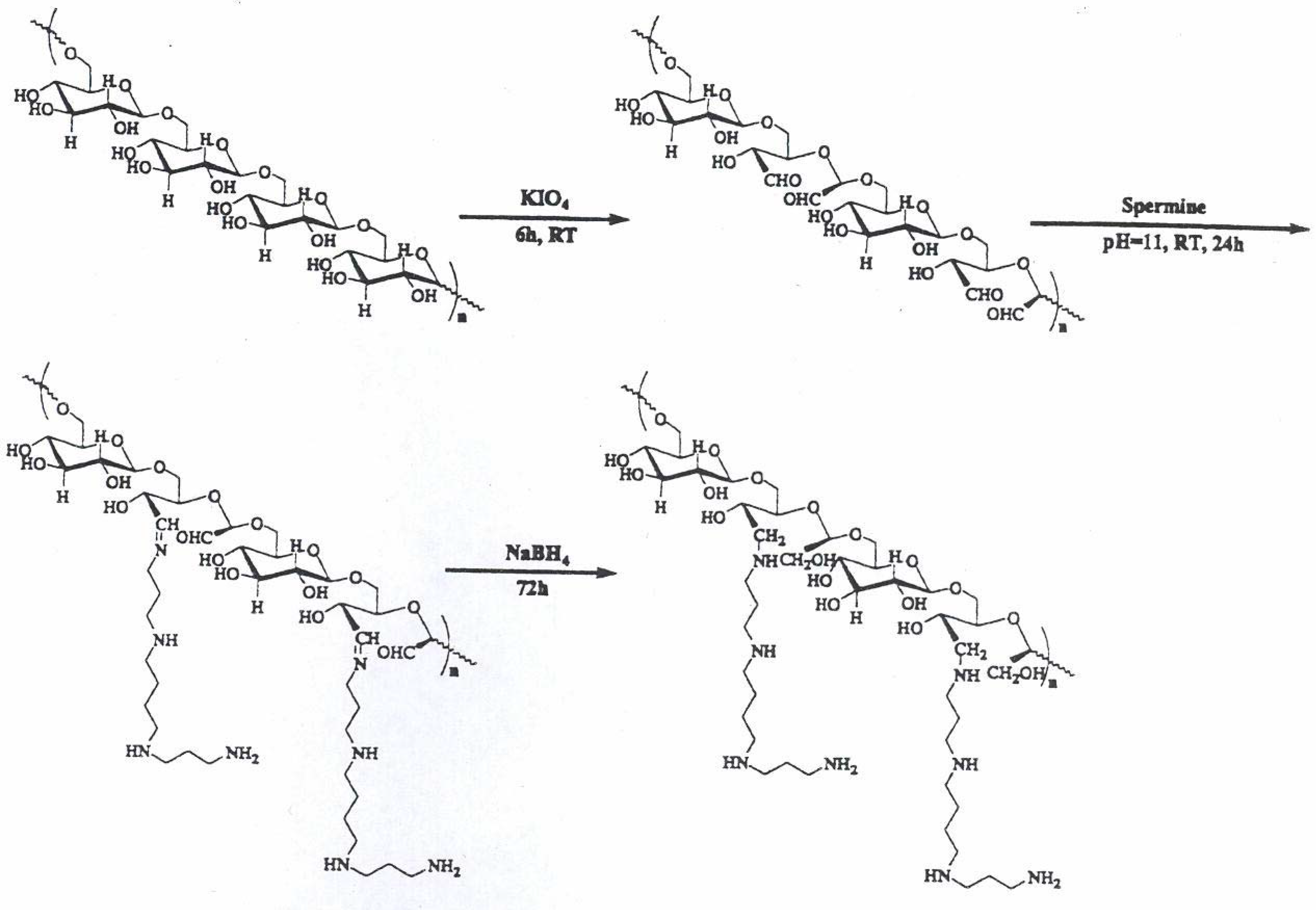
Polysaccharide-oligoamine based conjugates
Synthesis

In vitro transfection
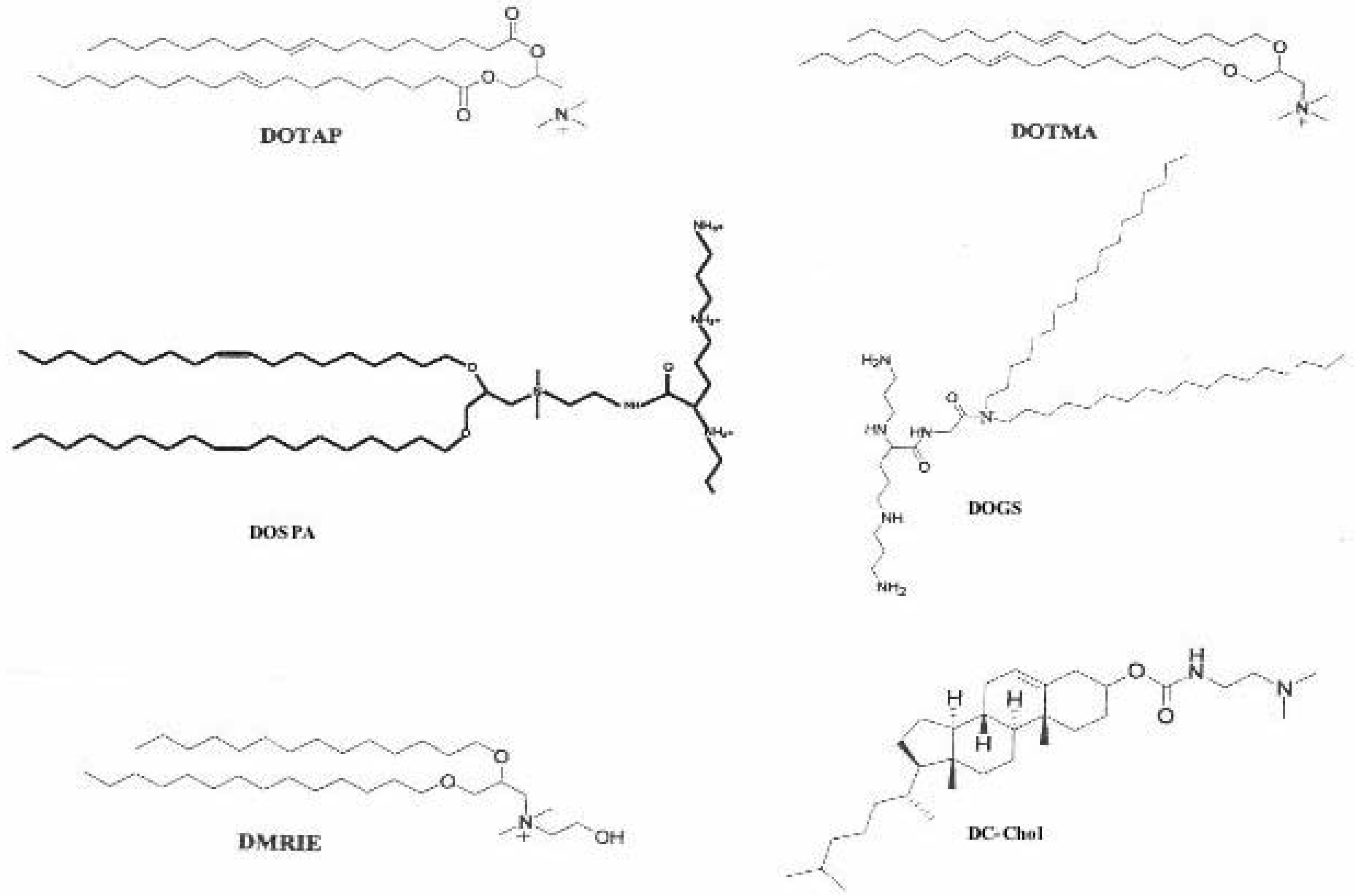
Cationic lipids
Structure size and morphology of cationic liposomes

Helper-lipids
Transfection of cells in culture
Uptake of cationic complexes
Escape from the endosome
Through the cytoplasm
Stability and mobility of naked DNA in the cytoplasm
Effects of DNA-complexation with cationic carriers
Dissociation of DNA from the complex
Into the nucleus
- (1)
- During mitosis the nuclear membrane disassembles and, thus, even large molecules, such as plasmids are able to gain access [154].
- (2)
- During interphases, the only way to enter the nucleus is through the NPC. Small molecules (< ~50 kDa, ~10 nm [155] or ions are able to diffuse passively through the NPC. The size and the steric properties of plasmid DNA [156,157] makes its enter via passive diffusion difficult [158]. Therefore, it was found that the passive entry of plasmid DNA becomes less efficient with increasing size [159], and no more than 0.1% of the plasmid copies microinjected into the cytoplasm reached the nucleus [144]. Larger molecules, such as proteins or RNA, require an “identification tag” that is recognized by receptors and, thus enables translocation into and out of the nucleus. The upper size limit for this form of entry is ~26 nm (~8 million Da) [160]. This size limit varies not only between species, but also within the same cell line depending on the confluency or energy status of the cells [160]. The diameter of the substrate seems to be the most important property for passage across the NPC, as no limitation for the length of a substrate was found. These “identification tag” are termed nuclear localization sequences (NLS) if they mediate transport into the nucleus and nuclear export signals (NES) if they enable exit from the nucleus. An NLS is in fact a short amino acid sequence that enables the active transport of proteins or viral DNA into the nucleus [161]. NLS do not conform to a specific consensus sequence, very likely because they interact with different receptors. Several studies have demonstrated that the incorporation of NLS resulted in enhanced nuclear uptake and transgene expression [162,163,164,165,166,167,168,169].
From in-vitro to in-vivo
Hurdles at the systemic level
Biodistribution and gene expression after i.v. application
Steric stabilization of complexes
Local application
Toxicity
Conclusions
References
- http://www.geocities.com/socialism_2000/pages/genetic.html.
- http://www.ualberta.ca/~csps/JPPS1(2)/biotech.htm.
- Stribley, J. M.; Rehman, K. S.; Niu, H.; Christman, G. M. Gene therapy and reproductive medicine. Fertil. Steril. 2002, 77, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Nabel, G. J. Development of optimized vectors for gene therapy. Proc. Natl. Acad. Sci. USA. 1999, 96, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Hullett, D. A. Gene therapy in transplantation. J. Heart Lung Transplant. 1996, 15, 857–862. [Google Scholar] [PubMed]
- Rosenberg, S. A.; Aebersold, P.; Cornetta, K.; Kasid, A.; Morgan, R. A.; Moen, R.; et al. Gene transfer into humans—immunotherapy of patients with advanced melanoma, using tumor-infiltrating lymphocytes modified by retroviral gene transduction. N. Engl. J. Med. 1990, 323, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Wivel, N. A.; Wilson, J. M. Methods of gene delivery. Hematol. Oncol. Clin.North. Am. 1998, 12, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Crystal, R. G. Transfer of genes to humans: early lessons and obstacles to success. Science 1995, 270, 404–410. [Google Scholar] [PubMed]
- Kim, V. N.; Mitrophanous, K.; Kingsman, S. M.; Kingsman, A. J. Minimal requirement for a lentivirus vector based on human immunodeficiency virus type 1. J. Virol. 1998, 72, 811–816. [Google Scholar] [PubMed]
- Poeschla, E.; Corbeau, P.; Wong-Staal, F. Development of HIV vectors for anti-HIV gene therapy. Proc. Natl. Acad. Sci. USA. 1996, 93, 11395–11399. [Google Scholar] [PubMed]
- Jiao, S.; Williams, P.; Berg, R. K.; Hodgeman, B. A.; Liu, L. J.; Repetto, G.; Wolff, J. A. Direct gene transfer into nonhuman primate myofibers in vivo. Hum. Gene Ther. 1992, 3, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J. A.; Williams, P.; Acsadi, G.; Jiao, S.; Jani, A.; Chong, W. Conditions affecting direct gene transfer into rodent muscle in vivo. Biotechniques 1991, 11, 474–485. [Google Scholar] [PubMed]
- Hickman, M. A.; Malone, R. W.; Lehmann-Bruinsma, K.; Sih, T. R.; Knoell, D.; Szoka, F. C.; Walzem, R.; Carlson, D. M.; Powell, J. S. Gene expression following direct injection of DNA into liver. Hum. Gene Ther. 1994, 5, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Ardehali, A.; Fyfe, A.; Laks, H.; Drinkwater, D. C.; Qiao, J. H.; Lusis, A. J. Direct gene transfer into donor hearts at the time of harvest. J. Thorac. Cardiovasc. Surg. 1995, 109, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Vile, R. G.; Hart, I. R. Use of tissue-specific expression of the herpes simplex virus thymidine kinase gene to inhibit growth of established murine melanomas following direct intratumoral injection of DNA. Cancer Res. 1993, 53, 3860–3864. [Google Scholar] [PubMed]
- Culver, K. W. Gene therapy: a handbook for physicians; Mary Ann Liebert: New York, 1994. [Google Scholar]
- Nishi, T.; Yoshizato, K.; Yamashiro, S.; Takeshima, H.; Sato, K.; Hamada, K.; Kitamura, I.; Yoshimura, T.; Saya, H.; Kuratsu, J.; Ushio, Y. High-efficiency in vivo gene transfer using intraarterial plasmid DNA injection following in vivo electroporation. Cancer Res. 1996, 56, 1050–1055. [Google Scholar] [PubMed]
- Aihara, H.; Miyazaki, J. Gene transfer into muscle by electroporation in vivo. Nat. Biotechnol. 1998, 16, 867–870. [Google Scholar] [CrossRef]
- Ulmer, J. B.; Donnelly, J. J.; Parker, S. E.; Rhodes, G. H.; Felgner, P. L.; Dwarki, V. J.; et al. Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 1993, 259, 1745–1749. [Google Scholar] [PubMed]
- Ledley, F. D. Nonviral gene therapy: the promise of genes as pharmaceutical products. Hum. Gene Ther. 1995, 6, 1129–1144. [Google Scholar] [CrossRef] [PubMed]
- Co, D. O.; Borowski, A. H.; Leung, J. D.; van der Kaa, J.; Hengst, S.; Platenburg, G. J.; et al. Generation of transgenic mice and germline transmission of a mammalian artificial chromosome introduced into embryos by pronuclear microinjection. Chromosome Res. 2000, 8, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Harries, D.; May, S.; Gelbart, W. M.; Ben-Shaul, A. Structure, stability, and thermodynamics of lamellar DNA-lipid complexes. Biophys. J. 1998, 75, 159–173. [Google Scholar] [PubMed]
- Zuidam, N. J.; Barenholz, Y. Electrostatic and structural properties of complexes involving plasmid DNA and cationic lipids commonly used for gene delivery. Biochim. Biophys. Acta. 1998, 1368, 115–128. [Google Scholar] [PubMed] [Green Version]
- Benns, J. M.; Choi, J. S.; Mahato, R. I.; Park, J. S.; Kim, S. W. pH-sensitive cationic polymer gene delivery vehicle: N-Ac-poly(L-histidine)-graft-poly(L-lysine) comb shaped polymer. Bioconjug. Chem. 2000, 11, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Midoux, P.; Monsigny, M. Efficient gene transfer by histidylated polylysine/pDNA complexes. Bioconjug. Chem. 1999, 10, 406–411. [Google Scholar] [PubMed]
- Tang, M. X.; Redemann, C. T.; Szoka, F. C. Jr. In vitro gene delivery by degraded polyamidoamine dendrimers. Bioconjug. Chem. 1996, 7, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Bielinska, A.; Kukowska-Latallo, J. F.; Johnson, J.; Tomalia, D. A.; Baker, J. R., Jr. Regulation of in vitro gene expression using antisense oligonucleotides or antisense expression plasmids transfected using starburst PAMAM dendrimers. Nucleic Acids Res. 1996, 24, 2176–2182. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Bieber, T.; Li, Y.; Elsasser, H. P.; Kissel, T. A novel non-viral vector for DNA delivery based on low molecular weight, branched polyethylenimine: effect of molecular weight on transfection efficiency and cytotoxicity. Pharm. Res. 1999, 16, 1273–1279. [Google Scholar] [PubMed]
- Marschall, P.; Malik, N.; Larin, Z. Transfer of YACs up to 2.3 Mb intact into human cells with polyethylenimine. Gene Ther. 1999, 6, 1634–1637. [Google Scholar] [PubMed]
- Campeau, P.; Chapdelaine, P.; Seigneurin-Venin, S.; Massie, B.; Tremblay, J. P. Transfection of large plasmids in primary human myoblasts. Gene Ther. 2001, 8, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, M. A.; Guo, W.; Lee, R. J. Efficient gene transfer using reversibly cross-linked low molecular weight polyethylenimine. Bioconjug. Chem. 2001, 12, 989–994. [Google Scholar] [PubMed]
- Abdallah, B.; Hassan, A.; Benoist, C.; Goula, D.; Behr, J. P.; Demeneix, B. A. A powerful non-viral vector for in vivo gene transfer into the adult mammalian brain: polyethylenimine. Hum. Gene Ther. 1996, 7, 1947–1954. [Google Scholar]
- De Smedt, S. C.; Demeester, J.; Hennink, W. E. Cationic polymer based gene delivery systems. Pharm. Res. 2000, 17, 113–126. [Google Scholar] [PubMed]
- Guang Liu, W.; De Yao, K. Chitosan and its derivatives—a promising non-viral vector for gene transfection. J. Control. Release 2002, 83, 1–11. [Google Scholar] [PubMed]
- Thanou, M.; Florea, B. I.; Geldof, M.; Junginger, H. E.; Borchard, G. Quaternized chitosan oligomers as novel gene delivery vectors in epithelial cell lines. Biomaterials 2002, 23, 153–159. [Google Scholar] [PubMed]
- van de Wetering, P.; Schuurmans-Nieuwenbroek, N. M.; Hennink, W. E.; Storm, G. Comparative transfection studies of human ovarian carcinoma cells in vitro, ex vivo and in vivo with poly(2-(dimethylamino)ethyl methacrylate)-based polyplexes. J. Gene Med. 1999, 1, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Boussif, O.; Lezoualc'h, F.; Zanta, M. A.; Mergny, M. D.; Scherman, D.; Demeneix, B.; Behr, J. P. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: polyethylenimine. Proc. Natl. Acad. Sci. USA. 1995, 92, 7297–7301. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Moro, E.; Pettenazzo, A.; Behr, J. P.; Zacchello, F.; Scarpa, M. ExGen 500 is an efficient vector for gene delivery to lung epithelial cells in vitro and in vivo. Gene Ther. 1997, 4, 1100–1106. [Google Scholar] [PubMed]
- Coll, J. L.; Chollet, P.; Brambilla, E.; Desplanques, D.; Behr, J. P.; Favrot, M. In vivo delivery to tumors of DNA complexed with linear polyethylenimine. Hum. Gene Ther. 1999, 10, 1659–1666. [Google Scholar] [PubMed]
- Wightman, L.; Kircheis, R.; Rossler, V.; Carotta, S.; Ruzicka, R.; Kursa, M.; Wagner, E. Different behavior of branched and linear polyethylenimine for gene delivery in vitro and in vivo. J. Gene Med. 2001, 3, 362–372. [Google Scholar] [PubMed]
- Brunner, S.; Furtbauer, E.; Sauer, T.; Kursa, M.; Wagner, E. Overcoming the nuclear barrier: cell cycle independent non-viral gene transfer with linear polyethylenimine or electroporation. Mol. Ther. 2002, 5, 80–86. [Google Scholar] [PubMed]
- Wolfert, M. A.; Dash, P. R.; Nazarova, O.; Oupický, D.; Seymour, L. W.; Smart, S.; Strohalm, J.; Ulbrich, K. Polyelectrolyte vectors for gene delivery: influence of cationic polymer on biophysical properties of complexes formed with DNA. Bioconjug. Chem. 1999, 10, 993–1004. [Google Scholar] [PubMed]
- Merdan, T.; Kunath, K.; Fischer, D.; Kopeek, J.; Kissel, T. Intracellular processing of poly(ethylene imine)/ribozyme complexes can be observed in living cells by using confocal laser scanning microscopy and inhibitor experiments. Pharm. Res. 2002, 19, 140–146. [Google Scholar]
- Pouton, C. W.; Lucas, P.; Thomas, B. J.; Uduehi, A. N.; Milroy, D. A.; Moss, S. H. Polycation-DNA complexes for gene delivery: a comparison of the biopharmaceutical properties of cationic polypeptides and cationic lipids. J. Control. Release 1998, 53, 289–299. [Google Scholar] [PubMed]
- Wagner, E.; Plank, C.; Zatloukal, K.; Cotten, M.; Birnstiel, M. L. Influenza virus hemagglutinin HA-2 N-terminal fusogenic peptides augment gene transfer by transferrin-polylysine-DNA complexes: toward a synthetic virus-like gene-transfer vehicle. Proc. Natl. Acad. Sci. USA. 1992, 89, 7934–7938. [Google Scholar] [CrossRef] [PubMed]
- Fajac, I.; Allo, J. C.; Souil, E.; Merten, M.; Pichon, C.; Figarella, C.; Monsigny, M.; Briand, P.; Midoux, P. Histidylated polylysine as a synthetic vector for gene transfer into immortalized cystic fibrosis airway surface and airway gland serous cells. J. Gene Med. 2000, 2, 368–378. [Google Scholar] [PubMed]
- Merdan, T.; Kopecek, J.; Kissel, T. Prospects for cationic polymers in gene and oligonucleotide therapy against cancer. Adv. Drug Deliv. Rev. 2002, 54, 715–758. [Google Scholar] [PubMed]
- Azzam, T.; Eliyahu, H.; Shapira, L.; Linial, M.; Barenholz, Y.; Domb, A. J. Polysaccharide-oligoamine based conjugates for gene delivery. J. Med. Chem. 2002, 45, 1817–1824. [Google Scholar] [PubMed]
- Azzam, T.; Raskin, A.; Makovitzki, A.; Brem, H.; Vierling, P.; Lineal, M.; Domb, A. J. Cationic Polysaccharides for Gene Delivery. Macormolecules 2002, 35, 9947–9953. [Google Scholar]
- Azzam, T.; Eliyahu, H.; Makovitzki, A.; Domb, A. J. Dextran–spermine conjugate: an efficiect vector for gene delivery. Macromol. Symp. 2003, 195, 247–261. [Google Scholar] [CrossRef]
- Hosseinkhani, H.; Azzam, T.; Tabata, Y.; Domb, A. J. Dextran-spermine polycation: an efficient nonviral vector for in vitro and in vivo gene transfection. Gene Ther. 2004, 11, 194–203. [Google Scholar] [PubMed]
- Siiman, O.; Wilkinson, J.; Burshteyn, A.; Roth, P.; Ledis, S. Fluorescent neoglycoproteins: antibody-aminodextran-phycobiliprotein conjugates. Bioconjugate Chem. 1999, 10, 1090–1106. [Google Scholar]
- Lasic, D. D.; Templeton, N. S. Liposomes in gene therapy. Adv. Drug Deliv. Rev. 1996, 20, 221–266. [Google Scholar]
- Behr, J. P. DNA strongly binds to micelles and vesicles containing lipopolyamines or lipointercalants. Tetrahedron Lett. 1986, 27, 5861–5864. [Google Scholar]
- Felgner, P. L.; Gadek, T. R.; Holm, M.; Roman, R.; Chan, H. W.; Wenz, M.; Northrop, J. P.; Ringold, G. M.; Danielsen, M. Lipofection: a highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl. Acad. Sci. USA. 1987, 84, 7413–7417. [Google Scholar] [CrossRef]
- Felgner, J. H.; Kumar, R.; Sridhar, C. N.; Wheeler, C. J.; Tsai, Y. J.; Border, R.; Ramsey, P.; Martin, M.; Felgner, P. L. Enhanced gene delivery and mechanism studies with a novel series of cationic lipid formulations. J. Biol. Chem. 1994, 269, 2550–2561. [Google Scholar] [PubMed]
- Lee, E. R.; Marshall, J.; Siegel, C. S.; Jiang, C.; Yew, N. S.; Nichols, M. R.; Nietupski, J. B.; Ziegler, R. J.; Lane, M. B.; Wang, K. X.; Wan, N. C.; Scheule, R. K.; Harris, D. J.; Smith, A. E.; Cheng, S. H. Detailed analysis of structures and formulations of cationic lipids for efficient gene transfer to the lung. Hum. Gene Ther. 1996, 7, 1701–1717. [Google Scholar]
- Wang, J.; Guo, X.; Xu, Y.; Barron , L.; Szoka, F. C. Jr. Synthesis and characterization of long chain alkyl acyl carnitine esters. Potentially biodegradable cationic lipids for use in gene delivery. J. Med. Chem. 1998, 41, 2207–2215. [Google Scholar]
- Solodin, I.; Brown, C. S.; Bruno, M. S.; Chow, C. Y.; Jang, E. H.; Debs, R. J.; Heath, T. D. A novel series of amphiphilic imidazolinium compounds for in vitro and in vivo gene delivery. Biochemistry 1995, 34, 13537–13544. [Google Scholar] [PubMed]
- Wheeler, C. J.; Felgner, P. L.; Tsai, Y.; Marshall, J.; Shukhu, L.; Doh, S. G.; Hartikka, J.; Nietupski, J. B.; Manthrope, M.; Nichols, M.; Plewe, M.; Liang, X.; Norman, J.; Smith, A.; Cheng, S. H. A novel cationic lipid reagent greatly enhances plasmid DNA delivery and expression in mouse lung. Proc. Natl. Acad. Sci. USA. 1996, 93, 11454–11459. [Google Scholar] [PubMed]
- Paukku, T.; Lauraeus, S.; Huhtaniemi, I.; Kinnunen, P. K. Novel cationic liposomes for DNA-transfection with high efficiency and low toxicity. Chem. Phys. Lipids 1997, 87, 23–29. [Google Scholar] [PubMed]
- Gorman, C. M.; Aikawa, M.; Fox, B.; Fox, E.; Lapuz, C.; Michaud, B.; Nguyen, H.; Roche, E.; Sawa, T.; Wiener-Kronish, J. P. Efficient in vivo delivery of DNA to pulmonary cells using the novel lipid EDMPC. Gene Ther. 1997, 4, 983–992. [Google Scholar] [PubMed]
- Leventis, R.; Silvius, J. R. Interactions of mammalian cells with lipid dispersions containing novel metabolizable cationic amphiphiles. Biochim. Biophys. Acta. 1990, 1023, 124–132. [Google Scholar] [PubMed]
- Stamatatos, L.; Leventis, R.; Zuckermann, M. J.; Silvius, J. R. Interactions of cationic lipid vesicles with negatively charged phospholipid vesicles and biological membranes. Biochemistry 1988, 27, 3917–3925. [Google Scholar] [PubMed]
- Lenssen, K.; Jantscheff, P.; von Kiedrowski, G.; Massing, U. Combinatorial synthesis of new cationic lipids and high-throughput screening of their transfection properties. Chembiochem. 2002, 3, 852–858. [Google Scholar] [PubMed]
- Felgner, P. L.; Tsai, Y. J.; Sukhu, L.; Wheeler, C. J.; Manthorpe, M.; Marshall, J.; Cheng, S. H. Improved cationic lipid formulations for in vivo gene therapy. Ann. NY Acad. Sci. 1995, 772, 126–139. [Google Scholar] [PubMed]
- Scheule, R. K.; George, J. A.; Bagley, R. G.; Marshall, J.; Kaplan, J. M.; Akita, G. Y.; Wang, K. X.; Lee, E. R.; Harris, D. J.; Jiang, C.; Yew, N. S.; Smith, A. E.; Cheng, S. H. Basis of pulmonary toxicity associated with cationic lipid-mediated gene transfer to the mammalian lung. Hum. Gene Ther. 1997, 8, 689–707. [Google Scholar]
- Simberg, D.; Weisman, S.; Talmon, Y.; Faerman, A.; Shoshani, T.; Barenholz, Y. The role of organ vascularization and lipoplex-serum initial contact in intravenous murine lipofection. J Biol. Chem. 2003, 278, 39858–39865. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mounkes, L. C.; Liggitt, H. D.; Brown, C. S.; Solodin, I.; Heath, T. D.; Debs, R. J. Factors influencing the efficiency of cationic liposome-mediated intravenous gene delivery. Nat. Biotech. 1997, 15, 167–173. [Google Scholar]
- Kerner, M.; Meyuhas, O.; Hirsch-Lerner, D.; Rosen, L. J.; Min, Z.; Barenholz, Y. Interplay in lipoplexes between type of pDNA promoter and lipid composition determines transfection efficiency of human growth hormone in NIH3T3 cells in culture. Biochim. Biophys. Acta. 2001, 1532, 128–136. [Google Scholar] [PubMed]
- Zuidam, N. J.; Hirsch-Lerner, D.; Margulies, S.; Barenholz, Y. Lamellarity of cationic liposomes and mode of preparation of lipoplexes affect transfection efficiency. Biochim. Biophys. Acta. 1999, 1419, 207–220. [Google Scholar] [PubMed]
- Israelachvili, J. N.; Marcelja, S.; Horn, R. G. Physical principles of membrane organization. Q. Rev. Biophys. 1980, 13, 121–200. [Google Scholar] [PubMed]
- Radler, J. O.; Koltover, I.; Salditt, T.; Safinya, C. R. Structure of DNA–cationic liposome complexes: DNA intercalation in multilamellar membranes in distinct interhelical packing regimes. Science 1997, 275, 810–814. [Google Scholar] [PubMed] [Green Version]
- Koltover, I.; Salditt, T.; Radler, J. O.; Safinya, C. R. An inverted hexagonal phase of cationic liposome–DNA complexes related to DNA release and delivery. Science 1998, 281, 78–81. [Google Scholar] [PubMed]
- Simberg, D.; Danino, D.; Talmon, Y.; Minsky, A.; Ferrari, M. E.; Wheeler, C. J.; Barenholz, Y. Phase behavior, DNA ordering, and size instability of cationic lipoplexes. Relevance to optimal transfection activity. J Biol. Chem. 2001, 276, 47453–47459. [Google Scholar] [PubMed]
- Behr, J. P.; Demeneix, B.; Leoffler, J. P.; Perez-Mutul, J. Efficient gene transfer into mammalian primary endocrine cells with lipolyamine-coated-DNA. Proc. Natl. Acad. Sci. USA. 1989, 86, 6982–6986. [Google Scholar] [PubMed]
- Ferrari, M. E.; Nguyen, C. M.; Zelphati, O.; Tsai, Y.; Felgner, P. L. Analytical methods for the characterization of cationic lipid–nucleic acid complexes. Hum. Gene Ther. 1998, 9, 341–351. [Google Scholar] [PubMed]
- Pedroso de Lima, M. C.; Simoes, S.; Pires, P.; Faneca, H.; Duzgunes, N. Cationic lipid-DNA complexes in gene delivery: from biophysics to biological applications. Adv. Drug Deliv. Rev. 2001, 47, 277–294. [Google Scholar] [PubMed]
- Hui, S. W.; Langner, M.; Zhao, Y. L.; Hurley, E.; Chan, K. The role of helper lipids in cationic liposome-mediated gene transfer. Biophys. J. 1996, 71, 590–599. [Google Scholar] [PubMed]
- Mok, K. W. C.; Cullis, P. R. Structural and fusogenic properties of cationic liposomes in the presence of plasmid DNA. Biophys. J. 1997, 73, 2534–2545. [Google Scholar] [PubMed]
- Simões, S.; Slepushkin, V.; Gaspar, R.; Pedroso de Lima, M. C.; Düzgüne, N. Gene delivery by negatively charged ternary complexes of DNA, cationic liposomes and transferrin or fusigenic peptides. Gene Ther. 1998, 5, 955–964. [Google Scholar] [PubMed]
- Harvie, P.; Wong, F. M. P.; Bally, M. B. Characterization of lipid DNA interactions. I. Destabilization of bound lipids and DNA dissociation. Biophys. J. 1998, 75, 1040–1051. [Google Scholar]
- Simões, S.; Slepushkin, V.; Gaspar, R.; Pedroso de Lima, M.C.; Düzgüne, N. Mechanisms of gene transfer mediated by lipoplexes associated with targeting ligands and pH-sensitive peptides. Gene Ther. 1999, 6, 1798–1807. [Google Scholar] [PubMed]
- Hong, K.; Zheng, W.; Baker, A.; Papahadjopoulos, D. Stabilization of cationic liposome–plasmid DNA complexes by polyamines and poly(ethylene glycol)–phospholipid conjugates for efficient in vivo gene delivery. FEBS Lett. 1997, 400, 233–237. [Google Scholar] [PubMed] [Green Version]
- Song, Y. K.; Liu, D. Free liposomes enhance the transfection activity of DNA/lipid complexes in vivo by intravenous administration. Biochim. Biophys. Acta 1998, 1372, 141–150. [Google Scholar] [PubMed]
- Sternberg, B.; Hong, K.; Zheng, W.; Papahadjopoulos, D. Ultrastructural characterization of cationic liposome–DNA complexes showing enhanced stability in serum and high transfection activity in vivo. Biochim. Biophys. Acta 1998, 1375, 23–35. [Google Scholar] [PubMed]
- Smith, J. G.; Wedeking, T.; Vernachio, J. H.; Way, H.; Niven, R. W. Characterization and in vivo testing of a heterogeneous cationic lipid–DNA formulation. Pharm. Res. 1998, 15, 1356–1363. [Google Scholar] [PubMed]
- Crook, K.; Stevenson, B. J.; Dubochet, M.; Porteous, D. J. Inclusion of cholesterol in DOTAP transfection complexes increases the delivery of DNA to cells in vitro in the presence of serum. Gene Ther. 1998, 5, 137–143. [Google Scholar] [PubMed]
- Singh, A. K.; Kasinath, B. S.; Lewis, E. J. Interaction of polycations with cell-surface negative charges of epithelial cells. Biochim. Biophys. Acta 1992, 1120, 337–342. [Google Scholar] [PubMed]
- Godbey, W. T.; Wu, K. K.; Mikos, A. G. Tracking the intracellular path of poly(ethylenimine)/DNA complexes for gene delivery. Proc. Natl. Acad. Sci. U S A 1999, 96, 5177–5181. [Google Scholar] [PubMed]
- Mislick, K. A.; Baldeschwieler, J. D. Evidence for the role of proteoglycans in cation-mediated gene transfer. Proc. Natl. Acad. Sci. USA 1996, 93, 12349–12354. [Google Scholar] [PubMed]
- Lecocq, M.; Wattiaux-De Coninck, S.; Laurent, N.; Wattiaux, R.; Jadot, M. Uptake and intracellular fate of polyethylenimine in vivo. Biochem. Biophys. Res. Commun. 2000, 278, 414–418. [Google Scholar] [PubMed]
- Remy-Kristensen, A.; Clamme, J. P.; Vuilleumier, C.; Kuhry, J. G.; Mely, Y. Role of endocytosis in the transfection of L929 fibroblasts by polyethylenimine/DNA complexes. Biochim. Biophys. Acta 2001, 1514, 21–32. [Google Scholar] [PubMed]
- Zabner, J.; Fasbender, A. J.; Moninger, T.; Poellinger, K. A.; Welsh, M. J. Cellular and molecular barriers to gene transfer by a cationic lipid. J. Biol. Chem. 1995, 270, 18997–19007. [Google Scholar] [PubMed] [Green Version]
- Mounkes, L. C.; Zhong, W.; Cipres-Palacin, G. Heath TD, Debs RJ. Proteoglycans mediate cationic liposome-DNA complex-based gene delivery in vitro and in vivo. J. Biol. Chem. 1998, 273, 26164–26170. [Google Scholar] [PubMed]
- Bally, M. B.; Harvie, P.; Wong, F. M. P.; Kong, S.; Wasan, E. K.; Reimer, D. L. Biological barriers to cellular delivery of lipid-based DNA carriers. Adv. Drug Deliv. Rev. 1999, 38, 291–315. [Google Scholar] [PubMed]
- Cheng, P. W. Receptor ligand-facilitated gene transfer: Enhancement of liposome-mediated gene transfer and expression by transferrin. Hum. Gene Ther. 1996, 7, 275–282. [Google Scholar] [PubMed]
- Girão, T.; Simões, S.; Pires, P.; Nir, S.; Pedroso de Lima, M. C. Kinetic analysis of the initial steps involved in lipoplex–cell interactions: effect of various factors that influence transfection activity. Biochim. Biophys. Acta 2001, 1510, 136–151. [Google Scholar] [PubMed]
- Matsui, H.; Johnson, L. G.; Randell, S. H.; Boucher, R. C. Loss of binding and entry of liposome-DNA complexes decreases transfection efficiency in differentiated airway epithelial cells. J. Biol. Chem. 1997, 272, 1117–1126. [Google Scholar] [PubMed]
- Reimer, D. L.; Kong, S. , Bally, M. B. Analysis of cationic liposome-mediated interactions of plasmid DNA with murine and human melanoma cells in vitro. J. Biol. Chem. 1997, 272, 19480–19487. [Google Scholar] [PubMed]
- Friend, D. S.; Papahadjopoulos, D.; Debs, R. J. Endocytosis and intracellular processing accompanying transfection mediated by cationic liposomes. Biochim. Biophys. Acta 1996, 1278, 41–50. [Google Scholar] [PubMed]
- Stegmann, T.; Legendre, J. Y. Gene transfer mediated by cationic lipids: lack of a correlation between lipid mixing and transfection. Biochim. Biophys. Acta 1997, 1325, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Zuhorn, I. S.; Hoekstra, D. On the mechanism of cationic amphiphile-mediated transfection. To fuse or not to fuse: is that the question? J. Membr. Biol. 2002, 189, 167–179. [Google Scholar] [PubMed]
- Zelphati, O.; Szoka, F. C. Jr. Intracellular distribution and mechanism of delivery of oligonucleotides mediated by cationic lipids. Pharm. Res. 1996, 13, 1367–1372. [Google Scholar] [PubMed]
- Blau, S.; Jubeh, T. T.; Haupt, S. M.; Rubinstein, A. Drug targeting by surface cationization. Crit. Rev. Ther. Drug Carrier Syst. 2000, 17, 425–465. [Google Scholar] [PubMed]
- Frankel, A. D.; Pabo, C. O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 1988, 55, 1189–1193. [Google Scholar] [PubMed]
- Torchilin, V. P.; Rammohan, R.; Weissig, V.; Levchenko, T. S. TAT peptide on the surface of liposomes affords their efficient intracellular delivery even at low temperature and in the presence of metabolic inhibitors. Proc. Natl. Acad. Sci. USA 2001, 98, 8786–8791. [Google Scholar] [PubMed]
- Pierschbacher, M. D.; Ruoslahti, E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature 1984, 309, 30–33. [Google Scholar] [PubMed]
- Lu, X.; Deadman, J. J.; Williams, J. A.; Kakkar, V. V.; Rahman, S. Synthetic RGD peptides derived from the adhesive domains of snake-venom proteins: evaluation as inhibitors of platelet aggregation. Biochem. J. 1993, 296, 21–24. [Google Scholar] [PubMed]
- Erbacher, P.; Remy, J.S.; Behr, J.P. Gene transfer with synthetic virus-like particles via the integrin-mediated endocytosis pathway. Gene Ther. 1999, 6, 138–145. [Google Scholar] [PubMed]
- Hart, S.L.; Harbottle, R.P.; Cooper, R.; Miller, A.; Williamson, R.; Coutelle, C. Gene delivery and expression mediated by an integrin-binding peptide. Gene Ther. 1995, 2, 552–554. [Google Scholar] [PubMed]
- Colin, M.; Maurice, M.; Trugnan, G.; Kornprobst, M.; Harbottle, R. P.; Knight, A.; Cooper, R. G.; Miller, A. D.; Capeau, J.; Coutelle, C.; Brahimi-Horn, M. C. Cell delivery, intracellular trafficking and expression of an integrin-mediated gene transfer vector in tracheal epithelial cells. Gene Ther. 2000, 7, 139–152. [Google Scholar] [PubMed]
- Muller, K.; Nahde, T.; Fahr, A.; Muller, R.; Brusselbach, S. Highly efficient transduction of endothelial cells by targeted artificial virus-like particles. Cancer Gene Ther. 2001, 8, 107–117. [Google Scholar]
- Uike, H.; Sakakibara, R.; Iwanaga, K.; Ide, M.; Ishiguro, M. Efficiency of targeted gene delivery of ligand-poly--lysine hybrids with different crosslinks. Biosci. Biotechnol. Biochem. 1998, 62, 1247–1248. [Google Scholar] [PubMed]
- Wightman, L.; Patzelt, E.; Wagner, E.; Kircheis, R. Development of transferrin-polycation/DNA based vectors for gene delivery to melanoma cells. J. Drug Target. 1999, 7, 293–303. [Google Scholar] [PubMed]
- Kircheis, R.; Kichler, A.; Wallner, G.; Kursa, M.; Ogris, M.; Felzmann, T.; Buchberger, M.; Wagner, E. Coupling of cell-binding ligands to polyethylenimine for targeted gene delivery. Gene Ther. 1997, 4, 409–418. [Google Scholar] [PubMed]
- Mahato, R. I.; Takemura, S.; Akamatsu, K.; Nishikawa, M.; Takakura, Y.; Hashida, M. Physicochemical and disposition characteristics of antisense oligonucleotides complexed with glycosylated poly(L-lysine). Biochem. Pharmacol. 1997, 53, 887–895. [Google Scholar] [PubMed]
- Chowdhury, N. R.; Wu, C. H.; Wu, G. Y.; Yerneni, P. C.; Bommineni, V. R.; Chowdhury, J. R. Fate of DNA targeted to the liver by asialoglycoprotein receptor-mediated endocytosis in vivo. Prolonged persistence in cytoplasmic vesicles after partial hepatectomy. J. Biol. Chem. 1993, 268, 11265–11271. [Google Scholar] [PubMed]
- Stankovics, J.; Crane, A. M.; Andrews, E.; Wu, C. H.; Wu, G. Y.; Ledley, F. D. Overexpression of human methylmalonyl CoA mutase in mice after in vivo gene transfer with asialoglycoprotein/polylysine/DNA complexes. Hum. Gene Ther. 1994, 5, 1095–1104. [Google Scholar] [PubMed]
- Hashida, M.; Takemura, S.; Nishikawa, M.; Takakura, Y. Targeted delivery of plasmid DNA complexed with galactosylated poly(L-lysine). J. Controll. Release 1998, 53, 301–310. [Google Scholar]
- Kawakami, S.; Wong, J.; Sato, A.; Hattori, Y.; Yamashita, F.; Hashida, M. Biodistribution characteristics of mannosylated, fucosylated, and galactosylated liposomes in mice. Biochim. Biophys. Acta 2000, 1524, 258–265. [Google Scholar] [PubMed]
- Nishikawa, M.; Yamauchi, M.; Morimoto, K.; Ishida, E.; Takakura, Y.; Hashida, M. Hepatocyte-targeted in vivo gene expression by intravenous injection of plasmid DNA complexed with synthetic multi-functional gene delivery system. Gene Ther. 2000, 7, 548–555. [Google Scholar] [PubMed]
- Li, S.; Tan, Y.; Viroonchatapan, E.; Pitt, B. R.; Huang, L. Targeted gene delivery to pulmonary endothelium by anti-PECAM antibody. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, 504–511. [Google Scholar]
- Suh, W.; Chung, J. K.; Park, S. H.; Kim, S. W. Anti-JL1 antibody-conjugated poly(L-lysine) for targeted gene delivery to leukemia T cells. J. Controll. Release 2001, 72, 171–178. [Google Scholar]
- Yano, L.; Shimura, M.; Taniguchi, M.; Hayashi, Y.; Suzuki, T.; Hatake, K.; Takaku, F.; Ishizaka, Y. Improved gene transfer to neuroblastoma cells by a monoclonal antibody targeting RET, a receptor tyrosine kinase. Hum. Gene Ther. 2000, 11, 995–1004. [Google Scholar] [PubMed]
- Kim, H.; Muller, W. J. The role of the epidermal growth factor receptor family in mammary tumorigenesis and metastasis. Exp. Cell Res. 1999, 253, 78–87. [Google Scholar] [PubMed]
- Blessing, T.; Kursa, M.; Holzhauser, R.; Kircheis, R.; Wagner, E. Different strategies for formation of pegylated EGF-conjugated PEI/DNA complexes for targeted gene delivery. Bioconjug. Chem. 2001, 12, 529–537. [Google Scholar] [PubMed]
- Putnam, D.; Gentry, C. A.; Pack, D. W.; Langer, R. Polymer-based gene delivery with low cytotoxicity by a unique balance of side-chain termini. Proc. Natl. Acad. Sci. USA 2001, 98, 1200–1205. [Google Scholar] [PubMed] [Green Version]
- Akinc, A; Langer, R. Measuring the pH environment of DNA delivered using nonviral vectors: implications for lysosomal trafficking. Biotechnol. Bioeng. 2002, 78, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, N. D.; Szoka, F. C. Jr.; Verkman, A. S. Chloride accumulation and swelling in endosomes enhances DNA transfer by polyamine-DNA polyplexes. J. Biol. Chem. 2003, 278, 44826–44831. [Google Scholar] [PubMed]
- Kichler, A.; Leborgne, C.; Coeytaux, E.; Danos, O. Polyethylenimine-mediated gene delivery: a mechanistic study. J. Gene Med. 2001, 3, 135–144. [Google Scholar] [PubMed]
- Hafez, I. M.; Maurer, N.; Cullis, P. R. On the mechanism whereby cationic lipids promote intracellular delivery of polynucleic acids. Gene Ther. 2001, 8, 1188–1196. [Google Scholar] [PubMed]
- Wattiaux, R.; Jadot, M.; Dubois, F.; Misquith, S.; Wattiaux-De Coninck, S. Uptake of exogenous DNA by rat liver: effect of cationic lipids. Biochem. Biophys. Res. Commun. 1995, 213, 81–87. [Google Scholar] [PubMed]
- Xu, Y.; Szoka, F. C. Jr. Mechanism of DNA release from cationic liposome/DNA complexes used in cell transfection. Biochemistry 1996, 35, 5616–5623. [Google Scholar] [PubMed]
- El Ouahabi, A.; Thiry, M.; Pector, V.; Fuks, R.; Ruysschaert, J. M.; Vandenbranden, M. The role of endosome destabilizing activity in the gene transfer process mediated by cationic lipids. FEBS Lett. 1997, 414, 187–192. [Google Scholar] [PubMed]
- Noguchi, A.; Furuno, T.; Kawaura, C.; Nakanishi, M. Membrane fusion plays an important role in gene transfection mediated by cationic liposomes. FEBS Lett. 1998, 433, 169–173. [Google Scholar] [PubMed]
- Zhang, Z. Y.; Smith, B. D. High-generation polycationic dendrimers are unusually effective at disrupting anionic vesicles: membrane bending model. Bioconjug. Chem. 2000, 11, 805–814. [Google Scholar] [PubMed]
- Klemm, A. R.; Young, D.; Lloyd, J. B. Effects of polyethyleneimine on endocytosis and lysosome stability. Biochem. Pharmacol. 1998, 56, 41–46. [Google Scholar] [PubMed]
- Lechardeur, D.; Sohn, K. J.; Haardt, M.; Joshi, P. B.; Monck, M.; Graham, R. W.; Beatty, B.; Squire, J.; O'Brodovich, H.; Lukacs, G. L. Metabolic instability of plasmid DNA in the cytosol: a potential barrier to gene transfer. Gene Ther. 1999, 6, 482–497. [Google Scholar] [PubMed]
- Page, R. L.; Butler, S. P.; Subramanian, A.; Gwazdauskas, F. C.; Johnson, J. L.; Velander, W. H. Transgenesis in mice by cytoplasmic injection of polylysine/DNA mixtures. Transgenic Res. 1995, 4, 353–360. [Google Scholar] [PubMed]
- Lukacs, G. L.; Haggie, P.; Seksek, O.; Lechardeur, D.; Freedman, N.; Verkman, A. S. Size-dependent DNA mobility in cytoplasm and nucleus. J. Biol. Chem. 2000, 275, 1625–1629. [Google Scholar] [PubMed]
- Dowty, M. E.; Williams, P.; Zhang, G.; Hagstrom, J. E.; Wolff, J. A. Plasmid DNA entry into postmitotic nuclei of primary rat myotubes. Proc. Natl. Acad. Sci. USA 1995, 92, 4572–4576. [Google Scholar] [PubMed]
- Godbey, W. T.; Wu, K. K.; Hirasaki, G. J.; Mikos, A. G. Improved packing of poly(ethylenimine)/DNA complexes increases transfection efficiency. Gene Ther. 1999, 6, 1380–1388. [Google Scholar] [PubMed]
- Pollard, H.; Remy, J. S.; Loussouarn, G.; Demolombe, S.; Behr, J. P.; Escande, D. Polyethylenimine but not cationic lipids promotes transgene delivery to the nucleus in mammalian cells. J. Biol. Chem. 1998, 273, 7507–7511. [Google Scholar] [PubMed]
- Hill, I. R.; Garnett, M. C.; Bignotti, F.; Davis, S.S. Determination of protection from serum nuclease activity by DNA-polyelectrolyte complexes using an electrophoretic method. Anal. Biochem. 2001, 291, 62–68. [Google Scholar] [PubMed]
- Godbey, W. T.; Barry, M. A.; Saggau, P.; Wu, K. K.; Mikos, A. G. Poly(ethylenimine)-mediated transfection: a new paradigm for gene delivery. J. Biomed. Mater. Res. 2000, 51, 321–328. [Google Scholar] [PubMed]
- Tang, J. X.; Janmey, P. A. The polyelectrolyte nature of F-actin and the mechanism of actin bundle formation. J. Biol. Chem. 1996, 271, 8556–8563. [Google Scholar] [PubMed]
- Wong, G. C.; Tang, J. X.; Lin, A.; Li, Y.; Janmey, P. A.; Safinya, C. R. Hierarchical self-assembly of F-actin and cationic lipid complexes: stacked three-layer tubule networks. Science 2000, 288, 2035–2039. [Google Scholar] [PubMed] [Green Version]
- Kabanov, A. V. Taking polycation gene delivery systems from in vitro to in vivo. Pharm. Sci. Technol. Today 1999, 2, 365–372. [Google Scholar] [PubMed]
- Wattiaux, R.; Laurent, N.; Wattiaux-De Coninck, S.; Jadot, M. Endosomes, lysosomes: their implication in gene transfer. Adv. Drug Deliv. Rev. 2000, 41, 201–208. [Google Scholar] [PubMed]
- Luo, D.; Saltzman, W. M. Synthetic DNA delivery systems. Nat. Biotechnol. 2000, 18, 33–37. [Google Scholar] [PubMed]
- Vasu, S. K.; Forbes, D. J. Nuclear pores and nuclear assembly. Curr. Opin. Cell Biol. 2001, 13, 363–375. [Google Scholar] [PubMed]
- Matsuoka, Y.; Takagi, M.; Ban, T.; Miyazaki, M.; Yamamoto, T.; Kondo, Y.; Yoneda, Y. Identification and characterization of nuclear pore subcomplexes in mitotic extract of human somatic cells. Biochem. Biophys. Res. Commun. 1999, 254, 417–423. [Google Scholar]
- Brunner, S.; Sauer, T.; Carotta, S.; Cotten, M.; Saltik, M.; Wagner, E. Cell cycle dependence of gene transfer by lipoplex, polyplex and recombinant adenovirus. Gene Ther. 2000, 7, 401–407. [Google Scholar] [PubMed]
- Talcott, B.; Moore, M. S. Getting across the nuclear pore complex. Trends Cell Biol. 1999, 9, 312–318. [Google Scholar] [PubMed]
- Paine, P. L.; Moore, L. C.; Horowitz, S. B. Nuclear envelope permeability. Nature 1975, 254, 109–114. [Google Scholar] [PubMed]
- Peters, R. Fluorescence microphotolysis to measure nucleocytoplasmic transport and intracellular mobility. Biochim. Biophys. Acta 1986, 864, 305–359. [Google Scholar] [PubMed]
- Salman, H.; Zbaida, D.; Rabin, Y.; Chatenay, D.; Elbaum, M. Kinetics and mechanism of DNA uptake into the cell nucleus. Proc. Natl. Acad. Sci. USA 2001, 98, 7247–7252. [Google Scholar] [PubMed]
- Ludtke, J. J.; Zhang, G.; Sebestyen, M. G.; Wolff, J. A. A nuclear localization signal can enhance both the nuclear transport and expression of 1 kb DNA. J. Cell Sci. 1999, 112, 2033–2041. [Google Scholar] [PubMed]
- Feldherr, C. M.; Akin, D. Signal-mediated nuclear transport in proliferating and growth-arrested BALB/c 3T3 cells. J. Cell Biol. 1991, 115, 933–939. [Google Scholar] [PubMed]
- Cokol, M.; Nair, R.; Rost, B. Finding nuclear localization signals. EMBO Rep. 2000, 1, 411–415. [Google Scholar] [PubMed]
- Collas, P.; Husebye, H.; Alestrom, P. The nuclear localization sequence of the SV40 T antigen promotes transgene uptake and expression in zebrafish embryo nuclei. Transgenic Res. 1996, 5, 451–458. [Google Scholar] [PubMed]
- Collas, P.; Alestrom, P. Nuclear localization signals enhance germline transmission of a transgene in zebrafish. Transgenic Res. 1998, 7, 303–309. [Google Scholar] [PubMed]
- Liang, M. R.; Alestrom, P.; Collas, P. Glowing zebrafish: integration, transmission, and expression of a single luciferase transgene promoted by non-covalent DNA-nuclear transport peptide complexes. Mol. Reprod. Dev. 2000, 55, 8–13. [Google Scholar] [PubMed]
- Neves, C.; Escriou, V.; Byk, G.; Scherman, D.; Wils, P. Intracellular fate and nuclear targeting of plasmid DNA. Cell Biol. Toxicol. 1999, 15, 193–202. [Google Scholar] [PubMed]
- Branden, L. J.; Mohamed, A. J.; Smith, C. I. A peptide nucleic acid-nuclear localization signal fusion that mediates nuclear transport of DNA. Nat. Biotechnol. 1999, 17, 784–787. [Google Scholar] [PubMed]
- Zanta, M. A.; Belguise-Valladier, P.; Behr, J. P. Gene delivery: a single nuclear localization signal peptide is sufficient to carry DNA to the cell nucleus. Proc. Natl. Acad. Sci. USA 1999, 96, 91–96. [Google Scholar] [PubMed]
- Ciolina, C.; Byk, G.; Blanche, F.; Thuillier, V.; Scherman, D.; Wils, P. Coupling of nuclear localization signals to plasmid DNA and specific interaction of the conjugates with importin alpha. Bioconjug. Chem. 1999, 10, 49–55. [Google Scholar] [PubMed]
- Sebestyen, M. G.; Ludtke, J. J.; Bassik, M. C.; Zhang, G.; Budker, V.; Lukhtanov, E. A.; Hagstrom, J. E.; Wolff, J. A. DNA vector chemistry: the covalent attachment of signal peptides to plasmid DNA. Nat. Biotechnol. 1998, 16, 80–85. [Google Scholar] [PubMed]
- Egilmez, N. K.; Iwanuma, Y.; Bankert, R. B. Evaluation and optimization of different cationic liposome formulations for in vivo gene transfer. Biochem. Biophys. Res. Commun. 1996, 169–173. [Google Scholar]
- Simberg, D.; Weisman, S.; Talmon, Y. DOTAP, a typical cationic lipid: From A to Z. Crit. Rev. Ther. Drug Carr. Syst. 2004; in press. [Google Scholar]
- Ogris, M.; Brunner, S.; Schuller, S.; Kircheis, R.; Wagner, E. PEGylated DNA/transferrin-PEI complexes: reduced interaction with blood components, extended circulation in blood and potential for systemic gene delivery. Gene Ther. 1999, 6, 595–605. [Google Scholar]
- Oupický, D.; Koák; Dash, P. R.; Seymour, L. W.; Ulbrich, K. Effect of albumin and polyanion on the structure of DNA complexes with polycation containing hydrophilic non-ionic block. Bioconjug. Chem. 1999, 10, 764–772. [Google Scholar] [PubMed]
- Plank, C.; Mechtler, K.; Szoka, F. C. Jr.; Wagner, E. Activation of the complement system by synthetic DNA complexes: a potential barrier for intravenous gene delivery. Hum. Gene Ther. 1996, 7, 1437–1446. [Google Scholar] [PubMed]
- Dash, P. R.; Read, M. L.; Barrett, L. B.; Wolfert, M. A.; Seymour, L. W. Factors affecting blood clearance and in vivo distribution of polyelectrolyte complexes for gene delivery. Gene Ther. 1999, 6, 643–650. [Google Scholar] [PubMed]
- Verbaan, F. J.; Oussoren, C.; van Dam, I. M.; Takakura, Y.; Hashida, M.; Crommelin, D. J.; Hennink, W. E.; Storm, G. The fate of poly(2-dimethyl amino ethyl)methacrylate-based polyplexes after intravenous administration. Int. J. Pharm. 2001, 214, 99–101. [Google Scholar] [PubMed]
- Templeton, N. S.; Lasic, D. D.; Frederik, P. M.; Strey, H. H.; Roberts, D. D.; Pavlakis, G. N. Improved DNA:liposome complexes for increased systemic delivery and gene expression. Nat. Biotech. 1997, 15, 647–652. [Google Scholar]
- Yang, J.P.; Huang, L. Overcoming the inhibitory effect of serum on lipofection by increasing the charge ratio of cationic liposome and DNA. Gene Ther. 1997, 4, 950–960. [Google Scholar] [PubMed]
- Eliyahu, H.; Servel, N.; Domb, A. J.; Barenholz, Y. Lipoplex-induced hemagglutination: potential involvement in intravenous gene delivery. Gene Ther. 2002, 9, 850–858. [Google Scholar] [PubMed]
- Kunath, K.; vHarpe, A.; Petersen, H.; Fischer, D.; Voigt, K.; Kissel, T.; Bickel, U. The structure of PEG-modified poly(ethylene imines) influences biodistribution and pharmacokinetics of their complexes with NF-kappaB decoy in mice. Pharm. Res. 2002, 19, 810–817. [Google Scholar] [PubMed]
- Chollet, P.; Favrot, M. C.; Hurbin, A.; Coll, J. L. Side-effects of a systemic injection of linear polyethylenimine-DNA complexes. J. Gene Med. 2002, 4, 84–91. [Google Scholar] [PubMed]
- Goula, D.; Benoist, C.; Mantero, S.; Merlo, G.; Levi, G.; Demeneix, B. A. Polyethylenimine-based intravenous delivery of transgenes to mouse lung. Gene Ther. 1998, 5, 1291–1295. [Google Scholar] [PubMed]
- Zou, S. M.; Erbacher, P.; Remy, J. S.; Behr, J. P. Systemic linear polyethylenimine (L-PEI)-mediated gene delivery in the mouse. J. Gene Med. 2000, 2, 128–134. [Google Scholar] [PubMed]
- Goula, D.; Becker, N.; Lemkine, G. F.; Normandie, P.; Rodrigues, J.; Mantero, S.; Levi, G.; Demeneix, B. A. Rapid crossing of the pulmonary endothelial barrier by polyethylenimine/DNA complexes. Gene Ther. 2000, 7, 499–504. [Google Scholar] [PubMed]
- Sokol, P. P.; Longenecker, K. L.; Kachel, D. L.; Martin, W. J., 2nd. Mechanism of putrescine transport in human pulmonary artery endothelial cells. J. Pharmacol. Exp. Ther. 1993, 265, 60–66. [Google Scholar] [PubMed]
- Aziz, S. M.; Olson, J. W.; Gillespie, M. N. Multiple polyamine transport pathways in cultured pulmonary artery smooth muscle cells: regulation by hypoxia. Am. J. Respir. Cell Mol. Biol. 1994, 10, 160–166. [Google Scholar] [PubMed]
- Nguyen, H. K.; Lemieux, P.; Vinogradov, S. V.; Gebhart, C. L.; Guerin, N.; Paradis, G.; Bronich, T. K.; Alakhov, V. Y.; Kabanov, A. V. Evaluation of polyether-polyethyleneimine graft copolymers as gene transfer agents. Gene Ther. 2000, 7, 126–138. [Google Scholar] [PubMed]
- Toncheva, V.; Wolfert, M. A.; Dash, P. R.; Oupický, D.; Ulbrich, K.; Seymour, L. W.; Schacht, E. H. Novel vectors for gene delivery formed by self-assembly of DNA with poly(L-lysine) grafted with hydrophilic polymers. Biochim. Biophys. Acta 1998, 1380, 354–368. [Google Scholar] [PubMed]
- Meyer, O.; Kirpotin, D.; Hong, K.; Sternberg, B.; Park, J. W.; Woodle, M. C.; Papahadjopoulos, D. Cationic liposomes coated with polyethylene glycol as carriers for oligonucleotides. J. Biol. Chem. 1998, 273, 15621–15627. [Google Scholar] [CrossRef]
- Kircheis, R.; Wightman, L.; Schreiber, A.; Robitza, B.; Rossler, V.; Kursa, M.; Wagner, E. Polyethylenimine/DNA complexes shielded by transferrin target gene expression to tumors after systemic application. Gene Ther. 2001, 8, 28–40. [Google Scholar] [PubMed]
- Kopeek, J.; Kopeková, P.; Minko, T.; Lu, Z. HPMA copolymer–anticancer drug conjugates: design, activity, and mechanism of action. Eur. J. Pharm. Biopharm. 2000, 50, 61–81. [Google Scholar] [PubMed]
- Oupický, D.; Howard, K. A.; Konák, C.; Dash, P. R.; Ulbrich, K.; Seymour, L. W. Steric stabilization of poly--Lysine/DNA complexes by the covalent attachment of semitelechelic poly[N-(2-hydroxypropyl)methacrylamide]. Bioconjug. Chem. 2000, 11, 492–501. [Google Scholar] [PubMed]
- Oupický, D.; Parker, A. L.; Seymour, L. W. Laterally stabilized complexes of DNA with linear reducible polycations: strategy for triggered intracellular activation of DNA delivery vectors. J. Am. Chem. Soc. 2002, 124, 8–9. [Google Scholar]
- Oupický, D.; Carlisle, R. C.; Seymour, L. W. Triggered intracellular activation of disulfide crosslinked polyelectrolyte gene delivery complexes with extended systemic circulation in vivo. Gene Ther. 2001, 8, 713–724. [Google Scholar] [PubMed] [Green Version]
- Aoki, K.; Furuhata, S.; Hatanaka, K.; Maeda, M.; Remy, J. S.; Behr, J. P.; Terada, M.; Yoshida, T. Polyethylenimine-mediated gene transfer into pancreatic tumor dissemination in the murine peritoneal cavity. Gene Ther. 2001, 8, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kircheis, R.; Schuller, S.; Brunner, S.; Ogris, M.; Heider, K. H.; Zauner, W.; Wagner, E. Polycation-based DNA complexes for tumor-targeted gene delivery in vivo. J. Gene Med. 1999, 1, 111–120. [Google Scholar]
- Wells, J. M.; Li, L. H.; Sen, A.; Jahreis, G. P.; Hui, S. W. Electroporation-enhanced gene delivery in mammary tumors. Gene Ther. 2000, 7, 541–547. [Google Scholar] [PubMed]
- Boletta, A.; Benigni, A.; Lutz, J.; Remuzzi, G.; Soria, M. R.; Monaco, L. Non-viral gene delivery to the rat kidney with polyethylenimine. Hum. Gene Ther. 1997, 8, 1243–1251. [Google Scholar] [PubMed]
- Gautam, A.; Densmore, C. L.; Golunski, E.; Xu, B.; Waldrep, J. C. Transgene expression in mouse airway epithelium by aerosol gene therapy with PEI-DNA complexes. Mol. Ther. 2001, 3, 551–556. [Google Scholar] [PubMed]
- Goula, D.; Remy, J. S.; Erbacher, P.; Wasowicz, M.; Levi, G.; Abdallah, B.; Demeneix, B. A. Size, diffusibility and transfection performance of linear PEI/DNA complexes in the mouse central nervous system. Gene Ther. 1998, 5, 712–717. [Google Scholar] [PubMed]
- Lemkine, G. F.; Goula, D.; Becker, N.; Paleari, L.; Levi, G.; Demeneix, B. A. Optimisation of polyethylenimine-based gene delivery to mouse brain. J. Drug Target. 1999, 7, 305–312. [Google Scholar] [PubMed]
- Wang, S.; Ma, N.; Gao, S. J.; Yu, H.; Leong, K. W. Transgene expression in the brain stem effected by intramuscular injection of polyethylenimine/DNA complexes. Mol. Ther. 2001, 3, 658–664. [Google Scholar] [PubMed]
- Sawa, Y.; Kaneda, Y.; Bai, H. Z.; Suzuki, K.; Fujimoto, J.; Morishita, R.; Matsuda, H. Efficient transfer of oligonucleotides and plasmid DNA into the whole heart through the coronary artery. Gene Ther. 1998, 5, 1472–1480. [Google Scholar] [PubMed]
- Affleck, D. G.; Yu, L.; Bull, D. A.; Bailey, S. H.; Kim, S. W. Augmentation of myocardial transfection using TerplexDNA: a novel gene delivery system. Gene Ther. 2001, 8, 349–353. [Google Scholar] [PubMed]
- Meuli, M.; Liu, Y.; Liggitt, D.; Kashani-Sabet, M.; Knauer, S.; Meuli-Simmen, C.; Harrison, M. R.; Adzick, N. S.; Heath, T. D.; Debs, R. J. Efficient gene expression in skin wound sites following local plasmid injection. J. Invest. Dermatol. 2001, 116, 131–135. [Google Scholar] [PubMed] [Green Version]
- Baranov, A.; Glazkov, P.; Kiselev, A.; Ostapenko, O.; Mikhailov, V.; Ivaschenko, T.; Sabetsky, V.; Baranov, V. Local and distant transfection of mdx muscle fibers with dystrophin and LacZ genes delivered in vivo by synthetic microspheres. Gene Ther. 1999, 6, 1406–1414. [Google Scholar] [PubMed]
- Ma, X.; Glover, C.; Miller, H.; Goldstein, J.; O'Brien, E. Focal arterial transgene expression after local gene delivery. Can. J. Cardiol. 2001, 17, 873–883. [Google Scholar] [PubMed]
- Armeanu, S.; Pelisek, J.; Krausz, E.; Fuchs, A.; Groth, D.; Curth, R.; Keil, O.; Quilici, J.; Rolland, P. H.; Reszka, R.; Nikol, S. Optimization of non-viral gene transfer of vascular smooth muscle cells in vitro and in vivo. Mol. Ther. 2000, 1, 366–375. [Google Scholar] [PubMed]
- Freeman, D. J.; Niven, R. W. The influence of sodium glycocholate and other additives on the in vivo transfection of plasmid DNA in the lungs. Pharm. Res. 1996, 13, 202–209. [Google Scholar] [PubMed]
- Stribling, R.; Brunette, E.; Liggitt, D.; Gaensler, K.; Debs, R. Aerosol gene delivery in vivo. Proc. Natl. Acad. Sci. USA 1992, 89, 11277–11281. [Google Scholar] [PubMed]
- Guillaume, C.; Delepine, P.; Droal, C.; Montier, T.; Tymen, G.; Claude, F. Aerosolization of cationic lipid-DNA complexes: lipoplex characterization and optimization of aerosol delivery conditions. Biochem. Biophys. Res. Commun. 2001, 286, 464–471. [Google Scholar] [PubMed]
- Densmore, C. L.; Kleinerman, E. S.; Gautam, A.; Jia, S. F.; Xu, B.; Worth, L. L.; Waldrep, J. C.; Fung, Y. K.; T'Ang, A.; Knight, V. Growth suppression of established human osteosarcoma lung metastases in mice by aerosol gene therapy with PEI-p53 complexes. Cancer Gene Ther. 2001, 8, 619–627. [Google Scholar] [PubMed]
- Scheule, R. K. The role of CpG motifs in immunostimulation and gene therapy. Adv. Drug Deliv. Rev. 2000, 44, 119–134. [Google Scholar] [PubMed]
- Bramson, J. L.; Bodner, C. A.; Graham, R. W. Activation of host antitumoral responses by cationic lipid/DNA complexes. Cancer Gene Ther. 2000, 7, 353–359. [Google Scholar] [PubMed]
- Whitmore, M.; Li, S.; Huang, L. LPD lipopolyplex initiates a potent cytokine response and inhibits tumor growth. Gene Ther. 1999, 6, 1867–1875. [Google Scholar] [PubMed] [Green Version]
- Gautam, A.; Densmore, C. L.; Waldrep, J. C. Pulmonary cytokine responses associated with PEI-DNA aerosol gene therapy. Gene Ther. 2001, 8, 254–257. [Google Scholar] [PubMed]
- Freimark, B. D.; Blezinger, H. P.; Florack, V. J.; Nordstrom, J. L.; Long, S. D.; Deshpande, D. S.; Nochumson, S.; Petrak, K. L. Cationic lipids enhance cytokine and cell influx levels in the lung following administration of plasmid: cationic lipid complexes. J. Immunol. 1998, 160, 4580–4586. [Google Scholar] [PubMed]
- Tousignant, J. D.; Gates, A. L.; Ingram, L. A.; Johnson, C. L.; Nietupski, J. B.; Cheng, S. H.; Eastman, S. J.; Scheule, R. K. Comprehensive analysis of the acute toxicities induced by systemic administration of cationic lipid:plasmid DNA complexes in mice. Hum. Gene Ther. 2000, 11, 2493–2513. [Google Scholar] [PubMed]
© 2005 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Eliyahu, H.; Barenholz, Y.; Domb, A.J. Polymers for DNA Delivery. Molecules 2005, 10, 34-64. https://doi.org/10.3390/10010034
Eliyahu H, Barenholz Y, Domb AJ. Polymers for DNA Delivery. Molecules. 2005; 10(1):34-64. https://doi.org/10.3390/10010034
Chicago/Turabian StyleEliyahu, H., Y. Barenholz, and A. J. Domb. 2005. "Polymers for DNA Delivery" Molecules 10, no. 1: 34-64. https://doi.org/10.3390/10010034
APA StyleEliyahu, H., Barenholz, Y., & Domb, A. J. (2005). Polymers for DNA Delivery. Molecules, 10(1), 34-64. https://doi.org/10.3390/10010034