Introduction
The antibiotic era started in the 1950s, and from then onwards the use of plant antimicrobials declined [
1]; although it was not the case as far as the traditional healing systems that heavily rely on the medicines from the natural sources, especially plants, are concerned. The emergence and spread of microbial resistance is growing each day, thereby necessitating the development of new antimicrobials of natural or synthetic origin [
2]. As far as the natural sources are concerned, apart from the microbial sources, plants appear to be valuable antimicrobial resources. Plants can produce a large number of secondary metabolites that may exceed a hundred thousand molecules [
3]; all of these don’t have antimicrobial potential, but some of them can produce significant activity against the human pathogens. This activity is not necessarily of the same magnitude as that of the prevailing antimicrobials, but still provides some scope for optimism [
4]. Consequently, the search for such antimicrobials has intensified during recent times. This can be observed from the fact that whereas the number of articles published on the antimicrobial activities of medicinal plants during 1966 - 1994 was 115, the number of articles on the same subject appearing during 1995 - 2004, is 307 [
5]; a more than two-fold increase in just one decade, showing the growth of interest in the search for antimicrobials of natural origin.
Dodonaea viscosa (L.) Jaeq. commonly known as Ghawraskay (Pushto), is a viscid shrub that grows to a height of 2 m. It is used in folk medicine as a remedy for fever, rheumatism and gout. The crude extract has inhibitory effects against
Staphylococcus aureus, Streptococcus pyogenes, and Corynebacterium dephtherieae, but no activity against
Escherichia coli and
Pseudomonas aeruginosa, thereby suggesting potential against notable Gram positive organisms [
6]. Coxsackie virus B3 and influenza A virus were inhibited, demonstrating the antiviral potential, while the yeast
Candida albicans, conidiophore
Aspergillus fumigatus and dermatophyte
Trichophyton rubrum were not inhibited [
6]. More recently the antifungal activity of
Dodonaea viscosa against 40
C. albicans isolates was reported [
7]. It has also shown anti-inflammatory effects in experimental animals [
8].
Phytochemical investigations of
Dodonaea viscosa have resulted in the isolation of flavonoids [
9,
10,
13], saponins [
11], and diterpenes [
12]. Notable among these compounds are pinocembrin, santin, penduletin, alizarin, 5-hydroxy-3,6,7,4’-tetramethoxy flavone, 5,7,4’-trihydorxy-3,6-dimethoxy flavone, isorhamnetin-3-rhamnosylgalactoside, 5,7-dihydroxy-3’-(hydroxymethylbutyl)-3,6,4’-tri-methoxy flavones [
9], 5,6,4’-trihydroxy-3,7-dimethoxy flavone [
13], viscosol [
10], hautriwaic acid [
12,
13], dehydrohautriwaic acid, methyl dodonates [
12],
ent-15,16-epoxy-3β,8α-dihydroxy-9αH-labda-13(16),14-diene[
13], dodonoside A and dodonoside B [
11]. This plant has not been thoroughly studied for antimicrobial activities, so in this study we have subjected the ethanolic extract and subsequent fractions of
Dodonaea viscosa to assays to establish their antibacterial potential.
Results and Discussion
Geitie
et al. [
6] reported the absence of activity against Gram negative organisms, but interestingly in our study (
Table 1) we have observed promising antimicrobial activity against both Gram positive and negative organisms.
Table 1.
Antibacterial activity of Crude and Aqueous Extracts of Dodonaea viscosa.
Table 1.
Antibacterial activity of Crude and Aqueous Extracts of Dodonaea viscosa.
| | Average Zone of Inhibition (in mm) |
| Bacteria | Crude extract | Aqueous extract | Positive Control | Negative Control |
| Staphylococcus aureus(G+ve) | 12.0 ± 0.3 | 0.0 | 41.0 ± 1.1† | 0.0 |
| Micrococcus luteus (G+ve) | 12.9 ± 0.2 | 0.0 | 43.0 ± 0.5* | 0.0 |
| Bacillus subtilis (G+ve) | 13.3 ± 0.2 | 0.0 | 40.0 ± 0.4† | 0.0 |
| Bacillus cereus(G+ve) | 0.0 | 0.0 | 32.4 ± 0.4† | 0.0 |
| Escherichia coli (G-ve) | 11.0 ± 0.2 | 0.0 | 30.0 ± 0.5* | 0.0 |
| Pseudomonas aeruginosa (Gve) | 12.4 ± 0.4 | 0.0 | 32.0 ± 0.5† | 0.0 |
| Salmonella typhi(G-ve) | 0.0 | 0.0 | 33.0 ± 0.3* | 0.0 |
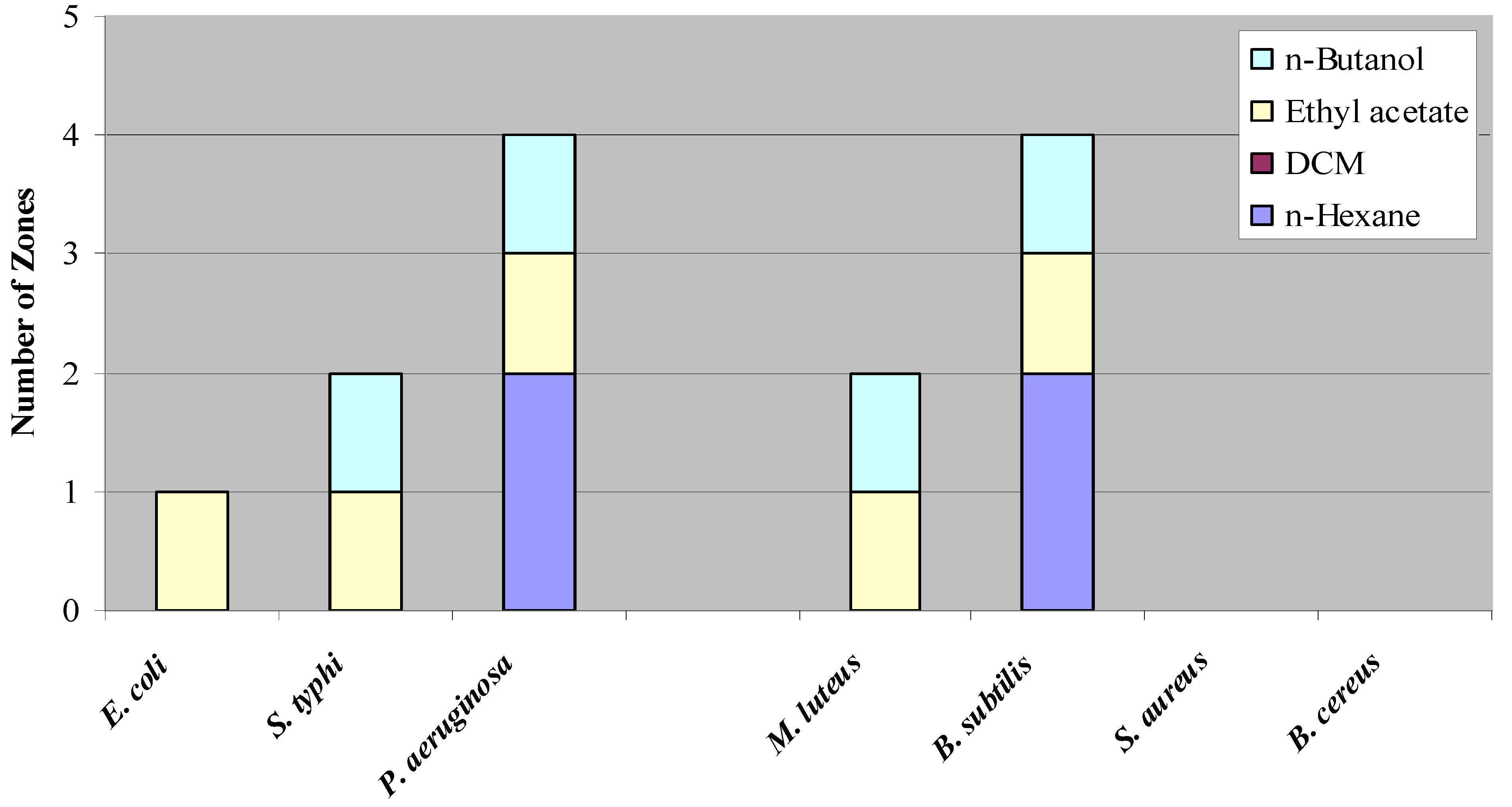
Figure 1.
Number of Inhibition Zones Against Test Organisms.
Figure 1.
Number of Inhibition Zones Against Test Organisms.
Figure 1 shows the number of zones observed against the various test organisms. It is clear from the figure that Gram negative organisms are more sensitive than the Gram positive organisms. The degree of sensitivity and location of the zones with respect to related
Rf value are listed in
Table (2). This inhibitory potential towards the Gram negative organisms was later confirmed when the crude extracts were subjected to contact bioautography. The presence of multiple inhibition zones against these organisms as well as the
S. typhi indicates the presence of entities capable of suppressing their growth.
Figure 1 shows the number of zones observed against the various test organisms. It is clear from the figure that Gram negative organisms are more sensitive than the Gram positive organisms. The degree of sensitivity and location of the zones with respect to related
Rf value are listed in
Table (2). This inhibitory potential towards the Gram negative organisms was later confirmed when the crude extracts were subjected to contact bioautography. The presence of multiple inhibition zones against these organisms as well as the
S. typhi indicates the presence of entities capable of suppressing their growth.
Table 2.
Location and Prominence of Zones of Inhibition at Different Rf values of Dodonaea viscosa fractions against Test Bacteria.
Table 2.
Location and Prominence of Zones of Inhibition at Different Rf values of Dodonaea viscosa fractions against Test Bacteria.
| | Zone/(s) of Inhibition at Rf value of Fraction |
|---|
| Bacteria | n-Hexane | DCM | Ethyl acetate | n-Butanol |
|---|
| Staphylococcus aureus | -* | - | - | - |
| Micrococcus luteus | - | - | 0.06 | 0.03 |
| (++) | (++) |
| Bacillus subtilis | 0.14 | - | 0.03 | 0.06 |
| 0.42 | | |
| (++) | (++) | (++) |
| Bacillus cereus | - | - | - | - |
| Escherichia coli | - | - | 0.03 | - |
| (+) |
| Salmonella typhi | - | - | 0.06 | 0.04 |
| (+) | (+) |
| Pseudomonas aeruginosa | 0.14 | - | 0.06 | 0.04 |
| 0.54 | | |
| (++) | (++) | (++) |
The results shows good activity against
B. subtilis, with prominent inhibition zones for the
n-hexane (two zones of inhibition), ethyl acetate (one zone of inhibition) and
n-butanol (one zone of inhibition) extracts.
M. luteus has one zone of inhibition each for the ethyl acetate and
n-butanol fractions. Interestingly,
S. aureus, that gave a 12.0 ± 0.3 mm zone of inhibition (
Table 1) failed to produce any zone of inhibition in the contact bioautography, which is most probably due to the synergestic action between the compounds present in the crude extract that got fractionated and later separated through the TLC, a reduced level or some chemical alteration of the active compound, thereby, resulting in the loss of anti-staphylococcal action.
The results given in
Table 2 shows that ethyl acetate fraction was active against five out of seven tested organisms, followed by the
n-butanol fraction that inhibits four organisms and the
n-hexane fraction that is inhibitory to two organisms. An interesting observation with respect to the
n-hexane fraction is the presence of multiple zones of inhibition, i.e., two zones each against
B. subtilis (at
Rf =0.14 and 0.42) and
P. aeruginosa (at
Rf =0.14 and 0.54). The antibacterial spectrum against Gram positive and Gram negative organisms, thereby, suggests presence of antibacterial entities that are capable of targeting either the bacterial wall or intracellular targets like bacterial ribosomes, etc.
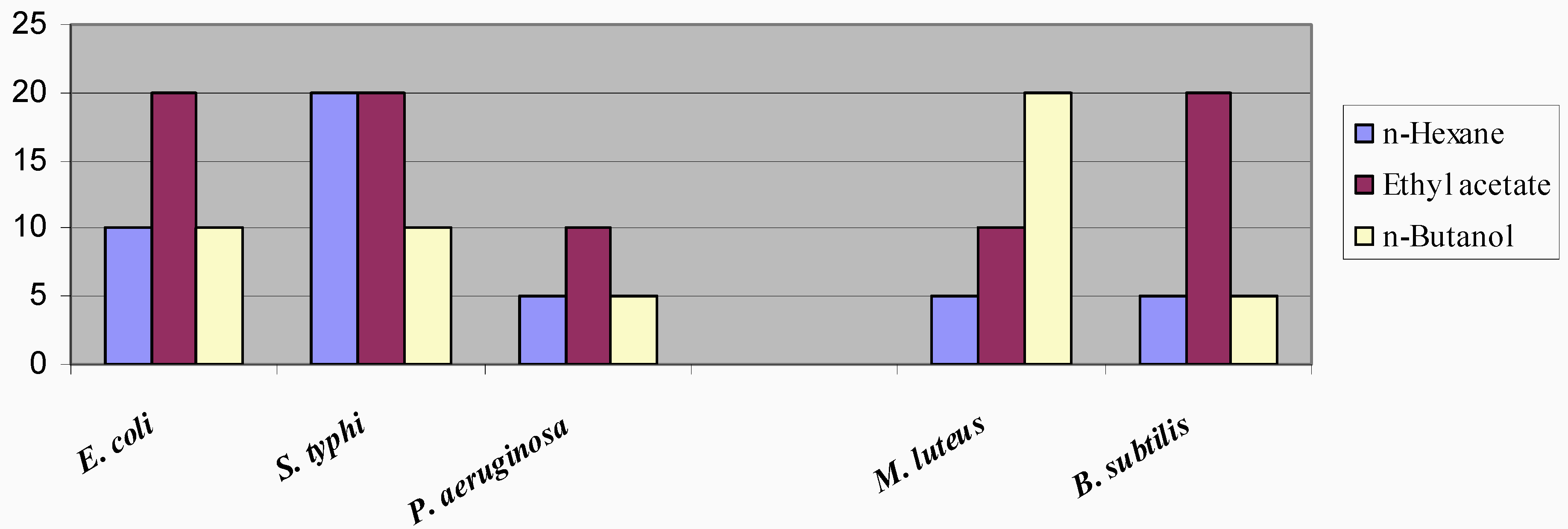
Minimum inhibitory concentrations (MICs) of the crude fractions (
Figure 2) were found to be within the range of 5 – 20.0 mg/mL. The high levels of the MIC's of the fractions are attributable to the facts that the active components are present in low concentrations, or there are some antagonistic components present that serve as growth promoters for the bacteria, thereby, necessitating the presence of high amount of the fraction to inhibit the growth.
Figure 2.
Minimum Inhibitory Concentrations (mg/ml) of Different Fractions of Dodonaea viscosa against Test Organisms.
Figure 2.
Minimum Inhibitory Concentrations (mg/ml) of Different Fractions of Dodonaea viscosa against Test Organisms.
The minimum bactericidal concentration (MBC) data (
Table 3) shows an extension of the MIC data. It depicts the concentrations of the fractions that completely eliminated (99.9% reduction) the viable bacteria under test. On the basis of the data it can be concluded that
n-hexane fraction has a higher bactericidal potential as compared to the ethyl acetate and
n-butanol fractions.
Table 3.
Minimum Bactericidal Concentrations (mg/mL) of Different Fractions of Dodonaea viscosa.
Table 3.
Minimum Bactericidal Concentrations (mg/mL) of Different Fractions of Dodonaea viscosa.
| Bacteria | Positive Control | n-hexane | ethyl acetate | n-butanol |
|---|
| Gram Positive |
| Micrococcus luteus | Chloramphenicol | 10 | 20 | >20 |
| Bacillus subtilis | Ciprofloxacin | 10 | 20 | 10 |
| Gram Negative |
| Escherichia coli | Chloramphenicol | 20 | >20 | 10 |
| Salmonella typhi | Chloramphenicol | >20 | >20 | 20 |
| Pseudomonas aeruginosa | Ciprofloxacin | 10 | 20 | 10 |
| | | | |
P. aeruginosa is an environmental inhabitant that is also an opportunistic pathogen. Its virulence is due to the production of exotoxin A, endotoxins, several proteolytic enzymes, alginates, pili, and hemolysins. It has been found to be involved in respiratory tract, urinary tract, bloodstream, and central nervous system infections of nosocomial origin [
16]. With growing resistance against the prevailing antibacterial agents, observations are that this pathogen is becoming resistant against gentamicin, ciprofloxacin [
17], tetracycline, chloramphenicol, and norfloxacin [
18].
E. coli is involved in causing severe infections of the urinary tract (of both community and nosocomial origin), sepsis, meningitis, and
E. coli-associated diarrheal diseases (Enteropathogenic
E. coli, Enterotoxigenic
E. coli, Enteroheamorrhagic
E. coli, Enteroinvasive
E. coli, and Enteroaggregative
E. coli) [
19].
E. coli resistance against fluoroquinolones, penicillins, cephalosporins, aminoglycosides [
20] and sulfamethoxazole are reported [
21].
S. typhi the causative agent for the enteric fevers, enterocolitis, and septicemia [
22] is also becoming resistant against the antibacterial like ampicillin, tetracycline and chloramphenicol [
23].
It is now an accepted fact that there is an increased resistance against the prevailing antimicrobials and our armory against these microorganisms is running out at a very quick pace. In order to withstand this challenge we need to develop new antimicrobials, which in itself is a great challenge and requires a huge investment. Turning back towards the molecules from natural sources like plants can be a good option. If these molecules are not as potent as the conventional antimicrobials they can be used as adjuncts, enhancing their therapeutic effectiveness. The isolation of active principles can be facilitated by bioassay guided fractionation. The present study indicates the presence of molecules that could have some therapeutic value if isolated and studied further.
Experimental
Plant material
Dodonaea viscosa (L.) Jaeq. belongs to the Sapindaceae family. The aerial parts of the plant were collected from Kohat, NWFP (Pakistan) in September 2007. A specimen was matched with the reference voucher number 592, preserved in the Herbarium of Pakistan, Department of Plant Sciences, Quaid-i-Azam University, Islamabad, Pakistan
Fractionation
The crude ethanolic extract was sequentially partitioned with n-hexane (3 x 1,000 mL), dichloromethane (3 x 1,000 mL), ethyl acetate (3 x 1,000 mL), and n-butanol (3 x 1,000 mL). A rotary evaporator was used to concentrate the portions that yielded a gummy n-hexane (75 g), syrupy dichloromethane (70 g) and ethyl acetate (92 g), gummy n-butanol (81 g) and an aqueous fraction (126 g), respectively.
Preliminary screening for the presence of antimicrobial activity
The crude extract was dissolved in 10% (v/v) dimethyl sulfoxide (DMSO, Fluka, Spain) in normal saline (0.9% w/v) to make the concentration equal to 3.2 mg/mL. The bacterial strains were cultured on nutrient agar (Merck, Germany) in 9 cm diameter plates, a single isolated colony of each test bacteria was picked and transferred to Nutrient Broth (Oxoid, UK) and incubated for a period of 18 hours except for the Salmonella typhi (4 hours incubation) and Micrococcus luteus (48 hours incubation) in a shaker. After incubation period the turbidity of the solutions were adjusted to 0.5 McFarland turbidity standard, using sterile nutrient broth. Hundred microliter of the inoculum was evenly spread over the entire area of 9 cm diameter Petri plates containing Muller Hinton Agar [Oxoid, UK] media, with the help of a sterile glass spreader and allowed to dry. With the help of a sterile cork borer, wells measuring 7 mm in diameter were made. Seventy microliter of crude extract solution was transferred aseptically into the wells. 10% (v/v) DMSO in normal saline and standard antibiotics served as negative and positive controls respectively. The plates were placed in upright position in refrigerator for a period of two hours in order to allow the materials to diffuse around the well area. The zones of inhibition were measured after an incubation of 24 hours at 37oC, in case of all the organisms, except for Micrococcus luteus, which was incubated for a period of 48 hours, in an upright position. All the tests were run in triplicate.
Minimum inhibitory concentration
The fractions that showed antibacterial potential were further assessed for the minimum inhibitory concentration (MIC), which is the minimal concentration of plant extract, or fraction thereof that inhibits the bacterial growth [
14]. The stock solutions of the fractions were prepared in pure DMSO in strength of 160 g/L and stored at +4 °C in a refrigerator till their use. Hundred microliter of stock solution containing approximately 16mg of each fraction was transferred aseptically to the micro-titer plate and sequentially half diluted in sterile solution of DMSO in normal saline (9% v/v) such that the last well contained ≈32μg/ml. The bacterial strains were prepared using Muller Hinton Broth (Oxoid, UK) and hundred microliter of the adjusted inoculum (turbidity adjusted equivalent to 0.5 MacFarland) was transferred aseptically to the 96-well micro-titer plate (TPP, Switzerland) in all of the wells containing the test dilutions, negative control (DMSO in normal saline) and standard antibiotics (either Ciprofloxacin or Chloramphenicol) containing wells that served as positive controls. The micro-titer plates were incubated at 37
oC for 24 hours for all the organisms except for the
M. luteus that required an incubation period of 48 hours. The tests were run in triplicate.
Minimum bactericidal concentration
It is defined as the concentration of the antimicrobial that results in a 99.9% reduction in CFU/ml compared with the organism concentration in the original inoculum [
15]. The micro-titer plates were prepared in the same manner as discussed above for MIC studies. After 24 hours incubation or 48 hours incubation in case of
M. luteus, from each of the visibly clear wells containing plant test materials, negative control and positive control, hundred microliter samples were taken and spread evenly on the entire surface of nutrient agar plates with the help of sterile glass spreader. The plates were allowed to dry and then kept in inverted position in incubator at 37
oC for 24 hours for all of the test organisms except the
M. luteus that was incubated for 48 hours at 37
oC. After the incubation period, the colonies were counted and compared with negative control. The concentration of the plant extract that completely inhibited the growth of the test organism, thereby, resulting in no growth or a 99.9% reduction in CFU/ml was taken as the MBC. The tests were run in triplicate.


{kind=link}
{kind=link}

