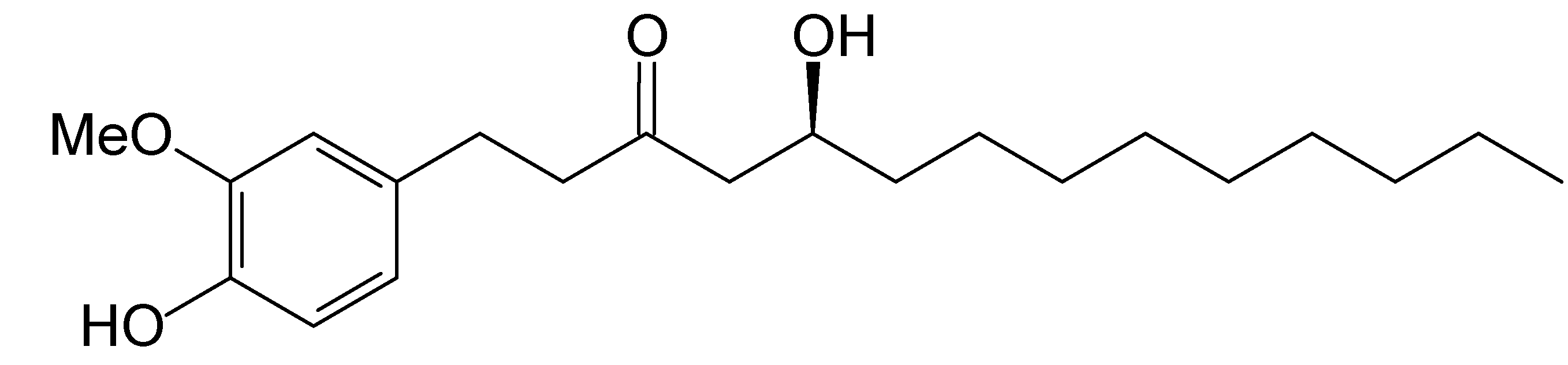
Effect of [10]-Gingerol on [Ca2+]i and Cell Death in Human Colorectal Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction

Results and Discussion
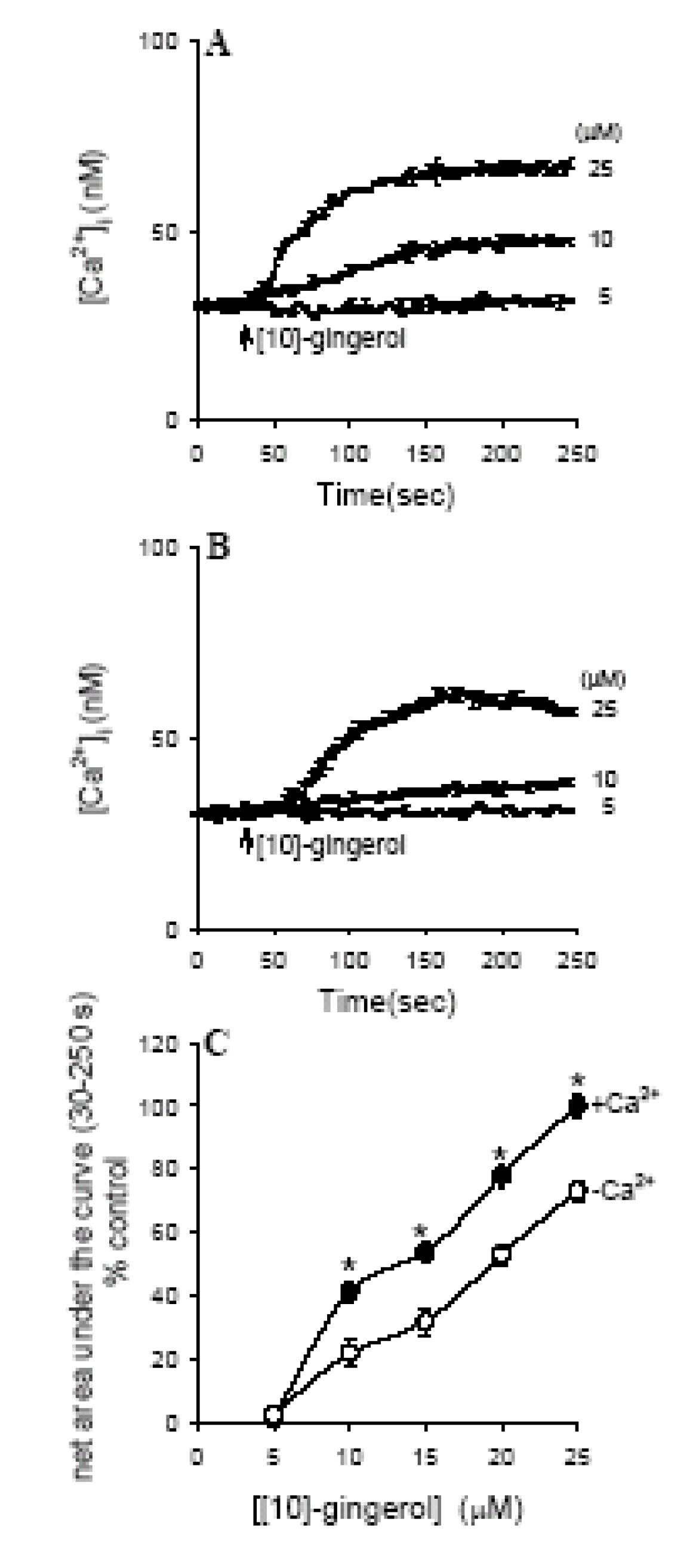
Effect of [10]-gingerol on [Ca2+]i

Effect of removing extracellular Ca2+ on [10]-gingerol-induced [Ca2+]i signals
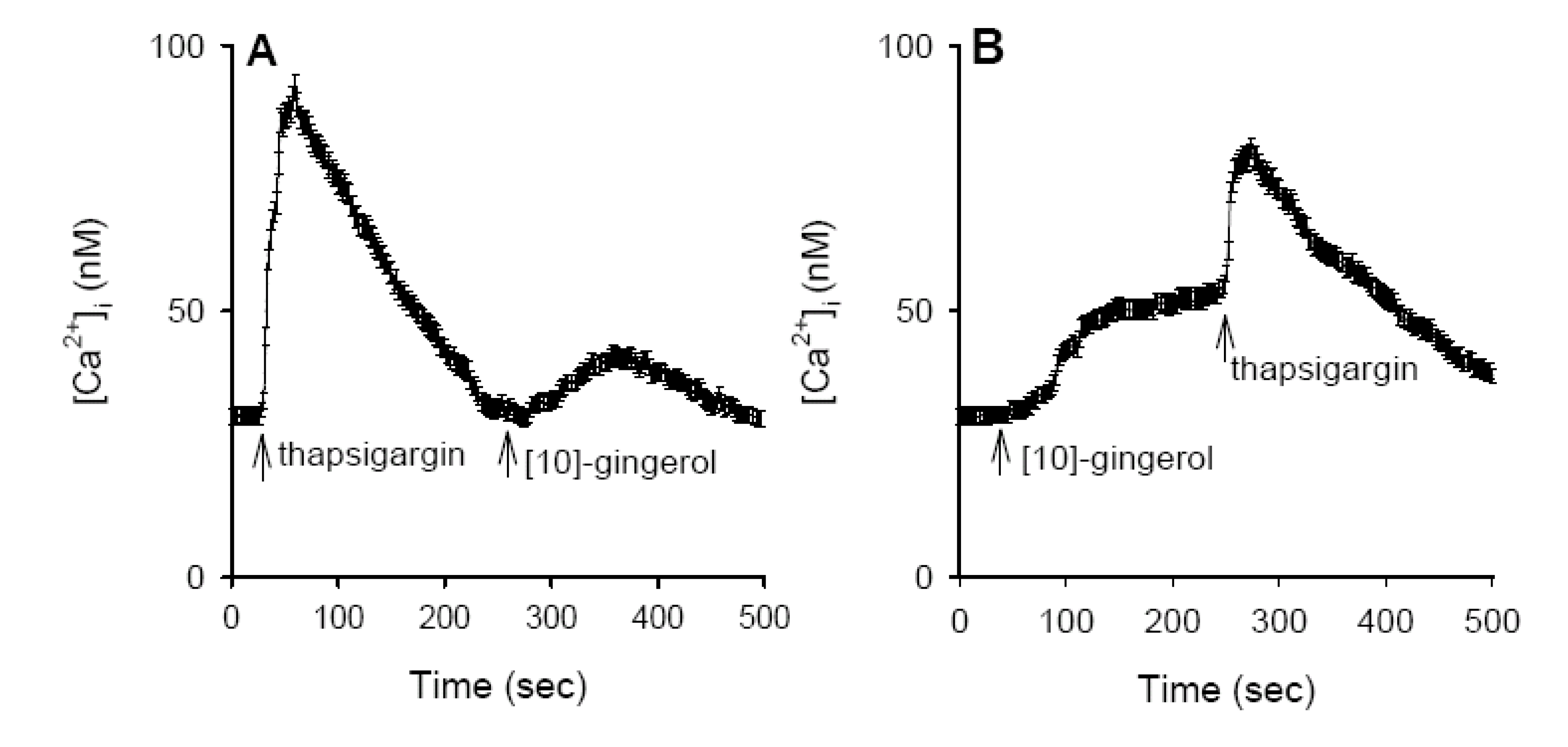
Internal Ca2+ stores for [10]-gingerol-induced [Ca2+]i Rises

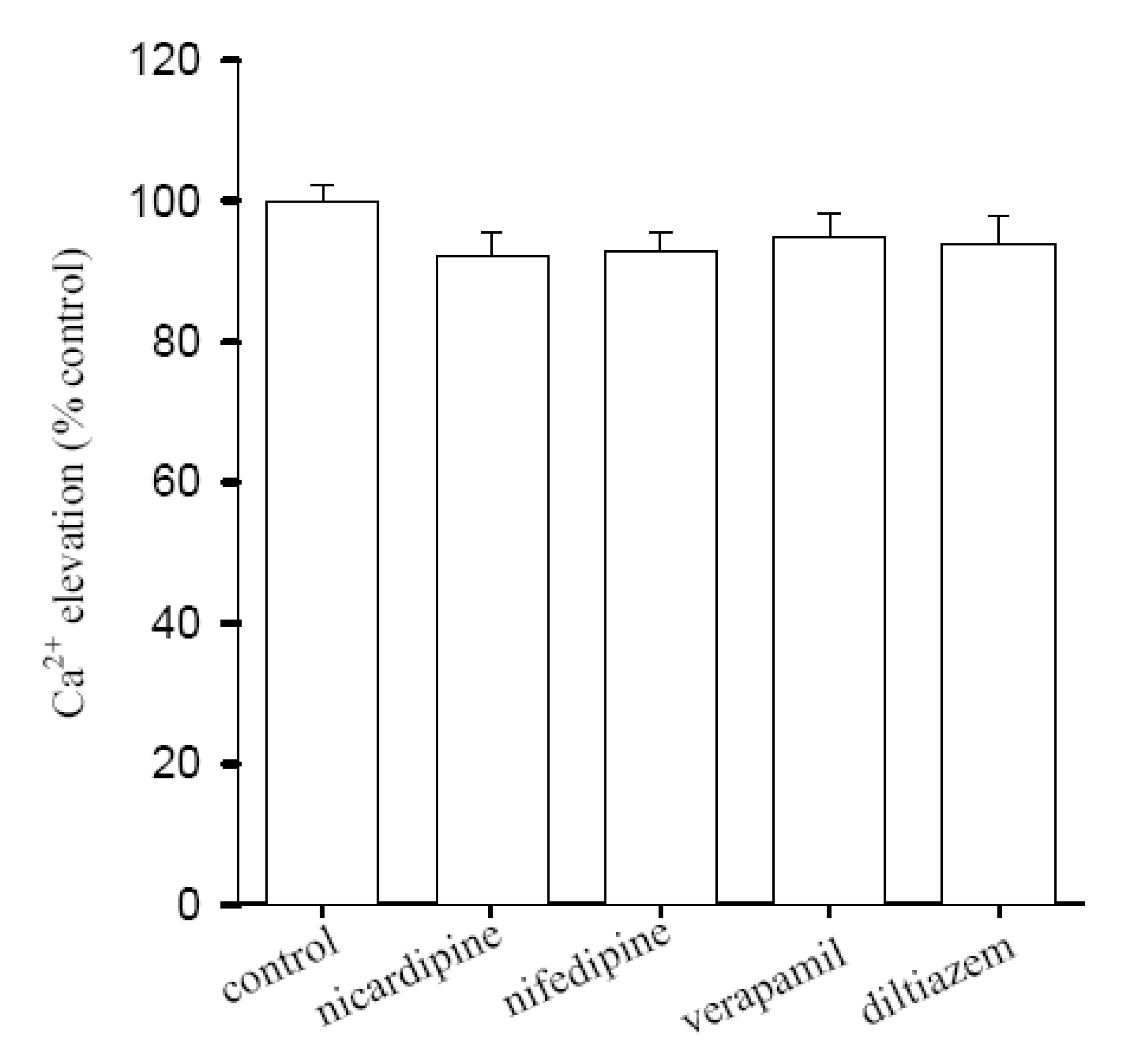
Lack of inhibitory effect of L-type Ca2+ entry blockers on Ca2+ release by [10]-gingerol

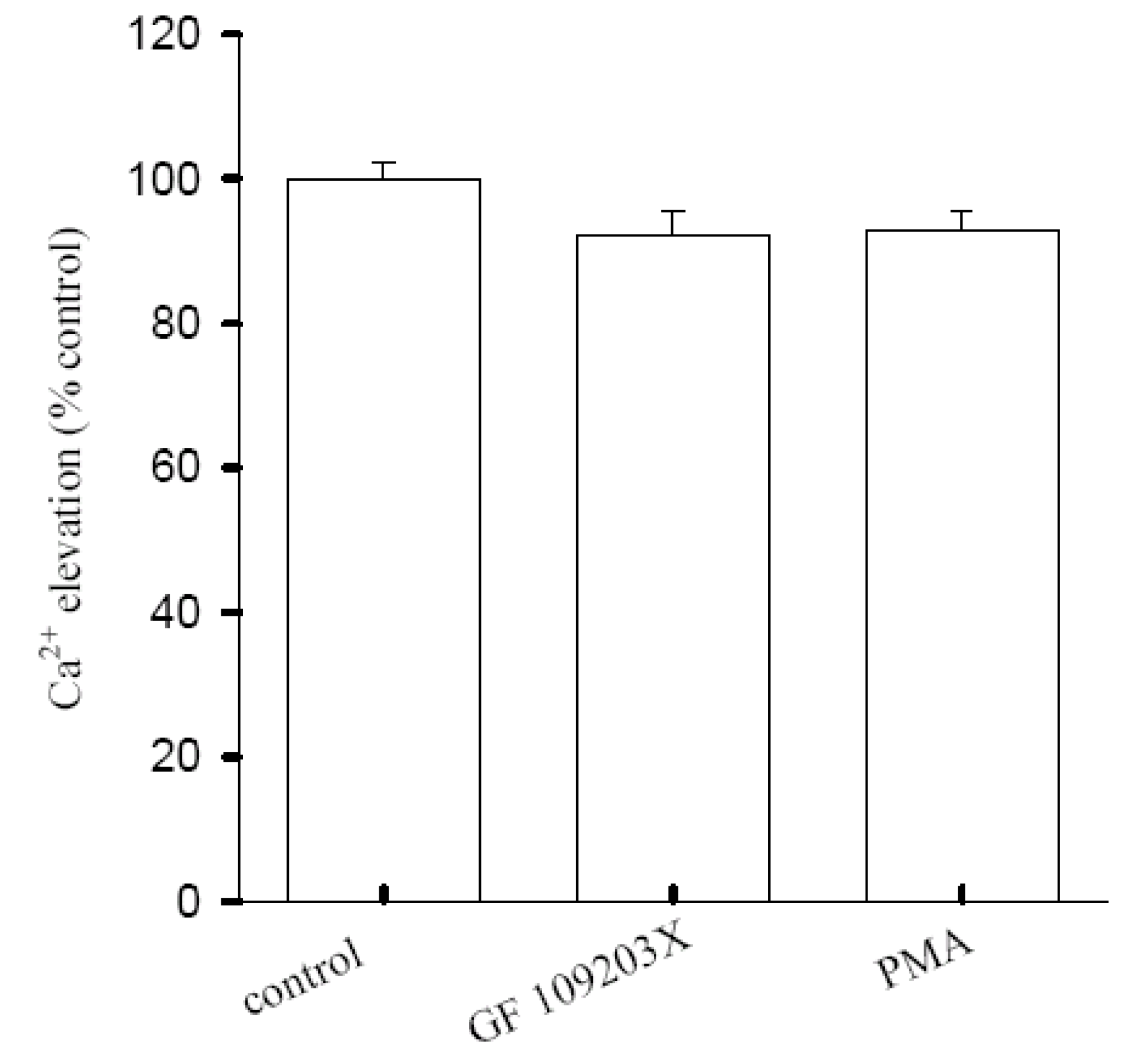
Regulation of [10]-gingerol-induced Ca2+ influx by Protein Kinase C

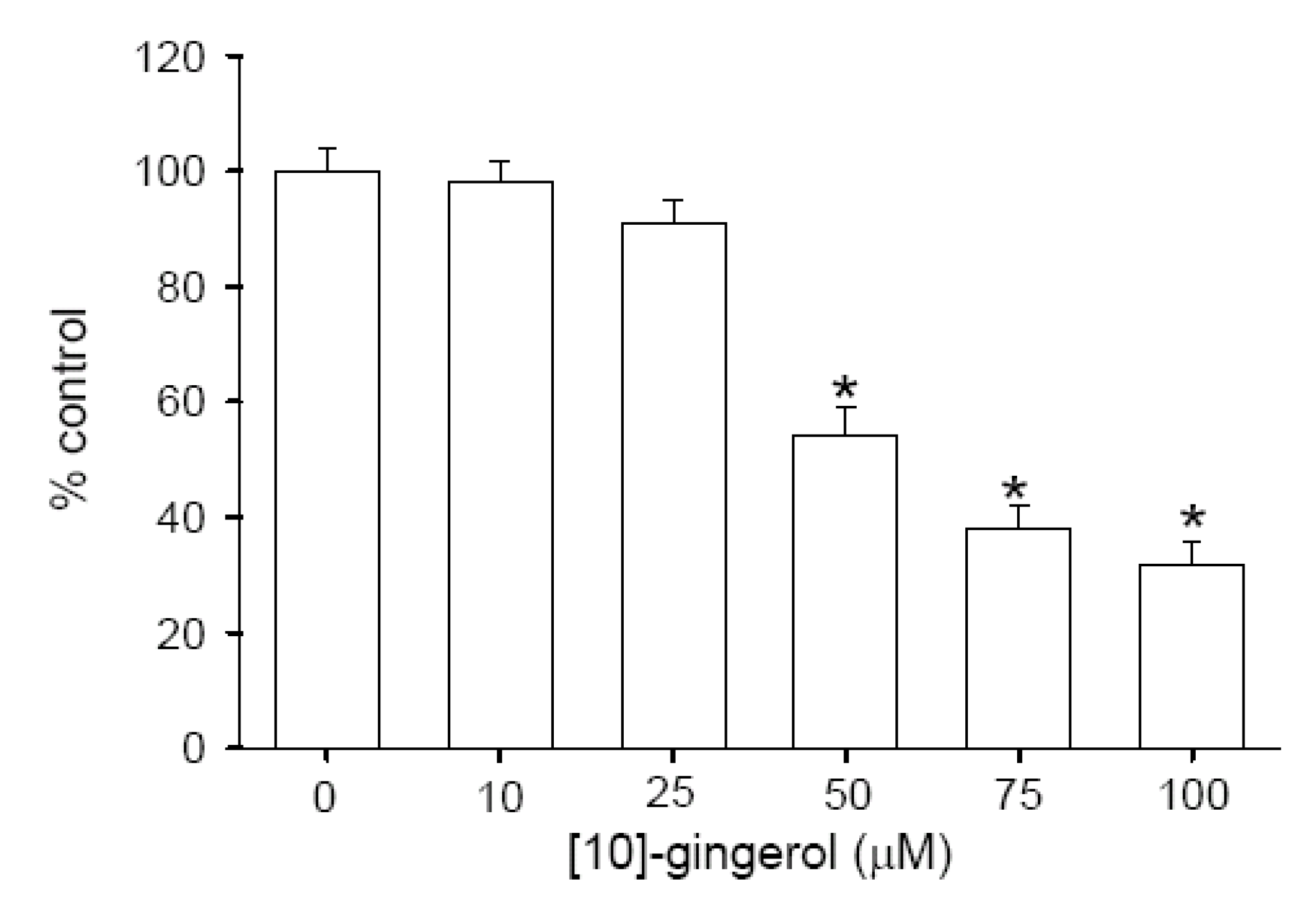
Effect of [10]-gingerol on Cell Viability

Experimental
General
Plant Material
Extraction and Isolation
Cell Culture and Test Compound Treatment
Measurement of [Ca2+]i
Cell viability assay
Data analysis
Acknowledgements
References
- Surh, Y. Molecular mechanisms of chemopreventive effects of selected dietary and medicinal phenolic substances. Muta. Res. 1999, 428, 305–327. [Google Scholar] [CrossRef]
- Altman, R.D.; Marcussen, K.C. Effects of a ginger extract on knee pain in patients with osteoarthritis. Arth. Rheum. 2001, 44, 2531–2538. [Google Scholar] [CrossRef]
- Grant, K.L.; Lutz, R.B. Alternative therapies: ginger. Am. J. Health-Syst. Pharm. 2000, 57, 945–947. [Google Scholar]
- Dorai, T.; Aggarwal, B.B. Role of chemopreventive agents in cancer therapy. Cancer Lett. 2004, 215, 129–140. [Google Scholar] [CrossRef]
- Mahmoud, N.N.; Carothers, A.M.; Grunberger, D.; Bilinski, R.T.; Churchill, M.R.; Martucci, C.; Newmark, H.L.; Bertagnolli, M.M. Plant phenolics decrease intestinal tumors in an animal model of familial adenomatous polyposis. Carcinogenesis 2000, 21, 921–927. [Google Scholar] [CrossRef]
- Kim, S.O.; Chun, K.S.; Kundu, J.K.; Surh, Y.J. Inhibitory effects of [6]-gingerol on PMA-induced COX-2 expression and activation of NFκB and p38 MAPK in mouse skin. Biofactors 2004, 21, 27–31. [Google Scholar] [CrossRef]
- Murakami, A.; Tanaka, T.; Lee, J.Y.; Surh, Y.J.; Kim, H.W.; Kawabata, K.; Nakamura, Y.; Jiwajinda, S.; Ohigashi, H. Zerumbone, a sesquiterpene in subtropical ginger, suppresses skin tumor initiation and promotion stages in ICR mice. Inter. J. Cancer 2004, 110, 481–490. [Google Scholar] [CrossRef]
- Surh, Y.J. Anti-tumor promoting potential of selected spice ingredients with antioxidative and anti-inflammatory activities: a short review. Food Chem. Toxicol. 2002, 40, 1091–1097. [Google Scholar] [CrossRef]
- Surh, Y.J.; Lee, E.; Lee, J.M. Chemoprotective properties of some pungent ingredients present in red pepper and ginger. Mut. Res. 1998, 402, 259–267. [Google Scholar] [CrossRef]
- Surh, Y.J.; Park, K.K.; Chun, K.S.; Lee, L.J.; Lee, E.; Lee, S.S. Anti-tumor- promoting activities of selected pungent phenolic substances present in ginger. J. Environ. Path. Toxicol. Oncol. 1999, 18, 131–139. [Google Scholar]
- Lee, E.; Surh, Y.J. Induction of apoptosis in HL-60 cells by pungent vanilloids, [6]-gingerol and [6]-paradol. Cancer Lett. 1998, 134, 163–168. [Google Scholar] [CrossRef]
- Kim, E.C.; Min, J.K.; Kim, T.Y.; Lee, S.J.; Yang, H.O.; Han, S.; Kim, Y.M.; Kwon, Y.G. [6]-Gingerol, a pungent ingredient of ginger, inhibits angiogenesis in vitro and in vivo. Biochem. Biophys. Res. Commun. 2005, 335, 300–308. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chen, C.H.; Kung, C.H.; Kuo, S.H.; Kuo, S.Y. [6]-Gingerol Induces Ca2+ Mobilization in Madin-Darby Canine Kidney Cells. J. Nat. Prod. 2008, 71, 137–140. [Google Scholar] [CrossRef]
- Ohizumi, Y.,; Sasaki, S.; Shibusawa, K.; Ishikawa, K.; Ikemoto, F. Stimulation of sarcoplasmic reticulum Ca(2+)-ATPase by gingerol analogues. Biol. Pharm. Bull. 1996, 19, 1377–1379. [Google Scholar] [CrossRef]
- Jemal, A.; Siegel, R.; Ward, E.; Murray, T.; Xu, J.; Thun, M.J. Cancer statistics. CA Cancer J. Clin. 2007, 57, 43–66. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signaling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef]
- Liantonio, A.; Giannuzzi, V.; Picollo, A.; Babini, E.; Pusch, M.; Conte Camerino, D. Niflumic acid inhibits chloride conductance of rat skeletal muscle by directly inhibiting the CLC-1 channel and by increasing intracellular calcium. Br. J. Pharmacol. 2007, 150, 235–247. [Google Scholar] [CrossRef]
- Zhang, F.; Wen, Q.; Mergler, S.; Yang, H.; Wang, Z.; Bildin, V.N.; Reinach, P.S. PKC isoform-specific enhancement of capacitative calcium entry in human corneal epithelial cells. Invest. Ophthalmol. Vis. Sci. 2006, 47, 3989–4000. [Google Scholar] [CrossRef]
- Berridge, M.J. Elementary and global aspects of calcium signaling. J. Physiol. 1997, 499, 291–306. [Google Scholar]
- Annunziato, L.; Amoroso, S.; Pannaccione, A.; Cataldi, M.; Pignataro, G.; D’Alessio, A.; Sirabella, R.; Secondo, A.; Sibaud, L.; Di Renzo, G.F. Apoptosis induced in neuronal cells by oxidative stress: role played by caspases and intracellular calcium ions. Toxicol. Lett. 2003, 139, 125–133. [Google Scholar] [CrossRef]
- Ma, Y.; Hendershot, L.M. The role of the unfolded protein response in tumour development: friend or foe? Nat. Rev. Cancer 2004, 4, 966–977. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: the calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Soboloff, J.; Berger, S.A. Sustained ER Ca2+ depletion suppresses protein synthesis and induces activation-enhanced cell death in mast cells. J. Biol. Chem. 2002, 277, 13812–13820. [Google Scholar] [CrossRef]
- Orrenius, S. Oxidative stress studied in intact mammalian cells. Phil. Trans R. Soc. Lond., B Biol. Sci. 1985, 311, 673–677. [Google Scholar] [CrossRef]
- Goldhaber, J.I.; Qayyum, M.S. Oxygen free radicals and excitation-contraction coupling. Antioxi. Redox. Signal. 2000, 2, 55–64. [Google Scholar] [CrossRef]
- Chakraborti, T.; Das, S.; Mondal, M.; Roychoudhury, S.; Chakraborti, S. Oxidant, mitochondria and calcium: an overview. Cell. Signal. 1999, 11, 77–85. [Google Scholar] [CrossRef]
- Putney, J.W. A model for receptor-regulated calcium entry. Cell Calcium 1986, 7, 1–12. [Google Scholar] [CrossRef]
- McFadzean, I.; Gibson, A. The developing relationship between receptor-operated and store-operated calcium channels in smooth muscle. Br. J. Pharmacol. 2002, 135, 1–13. [Google Scholar] [CrossRef]
- Shoji, N.; Iwasa, A.; Takemoto, T.; Ishida, Y. Cardiotonic principles of ginger (Zingiber officinale Roscoe). J. Pharm. Sci. 1982, 71, 1174–1175. [Google Scholar] [CrossRef]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar]
- Baliga, R.; Zhang, Z.; Shah, S.V. Role of cytochrome P-450 in hydrogen peroxide-induced cytotoxicity to LLC-PK cells. Kidney Int. 1996, 50, 1118–1124. [Google Scholar] [CrossRef]
- Andersson, D.A.; Zygmunt, P.M.; Movahed, P.; Andersson, T.L.; Hogestatt, E.D. Effects of inhibitors of small- and intermediate-conductance calcium-activated potassium channels, inwardly-rectifying potassium channels and Na+/K+ ATPase on EDHF relaxations in the rat hepatic artery. Br. J. Pharmacol. 2000, 129, 1490–1496. [Google Scholar] [CrossRef]
- Das, S.; Bhattacharyya, S.; Ghosh, S.; Majumdar, S. TNF-alpha induced altered signaling mechanism in human neutrophil. Mol. Cell. Biochem. 1999, 197, 97–108. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, S.I.; Park, H.W.; Yang, J.H.; Shin, T.Y.; Kim, Y.C.; Baek, N.I.; Kim, S.H.; Choi, S.U.; Kwon, B.M.; Leem, K.H.; Jung, M.Y.; Kim, D.K. Cytotoxic components from the dried rhizomes of Zingiber officinale Roscoe. Arch Pharm Res. 2008, 31, 415–418. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, C.-Y.; Li, Y.-W.; Kuo, S.-Y. Effect of [10]-Gingerol on [Ca2+]i and Cell Death in Human Colorectal Cancer Cells. Molecules 2009, 14, 959-969. https://doi.org/10.3390/molecules14030959
Chen C-Y, Li Y-W, Kuo S-Y. Effect of [10]-Gingerol on [Ca2+]i and Cell Death in Human Colorectal Cancer Cells. Molecules. 2009; 14(3):959-969. https://doi.org/10.3390/molecules14030959
Chicago/Turabian StyleChen, Chung-Yi, Yi-Wen Li, and Soong-Yu Kuo. 2009. "Effect of [10]-Gingerol on [Ca2+]i and Cell Death in Human Colorectal Cancer Cells" Molecules 14, no. 3: 959-969. https://doi.org/10.3390/molecules14030959
APA StyleChen, C. -Y., Li, Y. -W., & Kuo, S. -Y. (2009). Effect of [10]-Gingerol on [Ca2+]i and Cell Death in Human Colorectal Cancer Cells. Molecules, 14(3), 959-969. https://doi.org/10.3390/molecules14030959