Interest of Fluorescence Derivatization and Fluorescence Probe Assisted Post-column Detection of Phospholipids: A Short Review

Abstract
:Abbreviations
| GPA | Glycerophosphatidic acid |
| GPSer | Glycerophosphatidylserine |
| GPIns | Glycerophosphatidylinositol |
| GPEtn | Glycerophosphatidylethanolamine |
| GPCho | Glycerophosphatidylcholine |
| GPGro | Glycerophosphatidylglycerol |
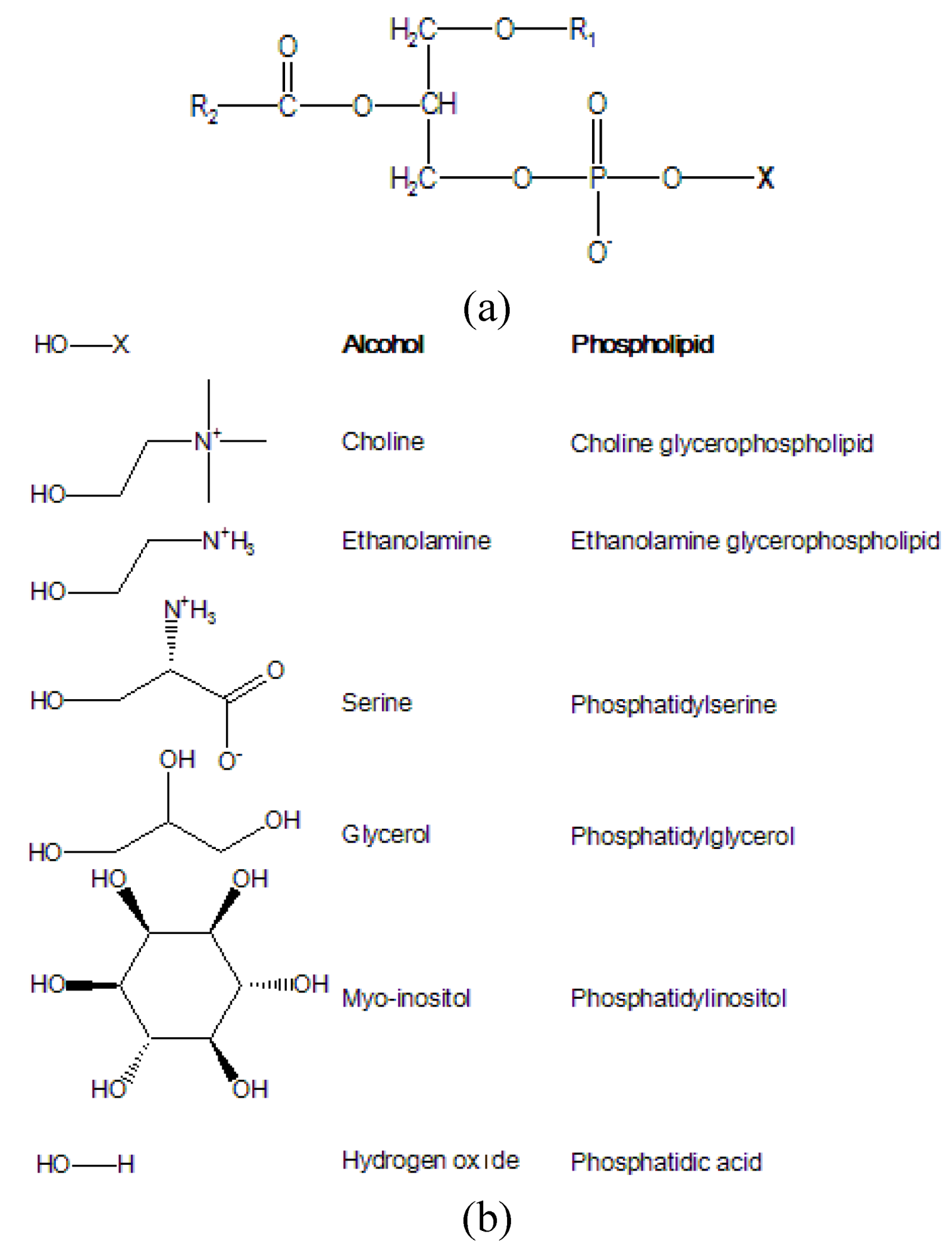
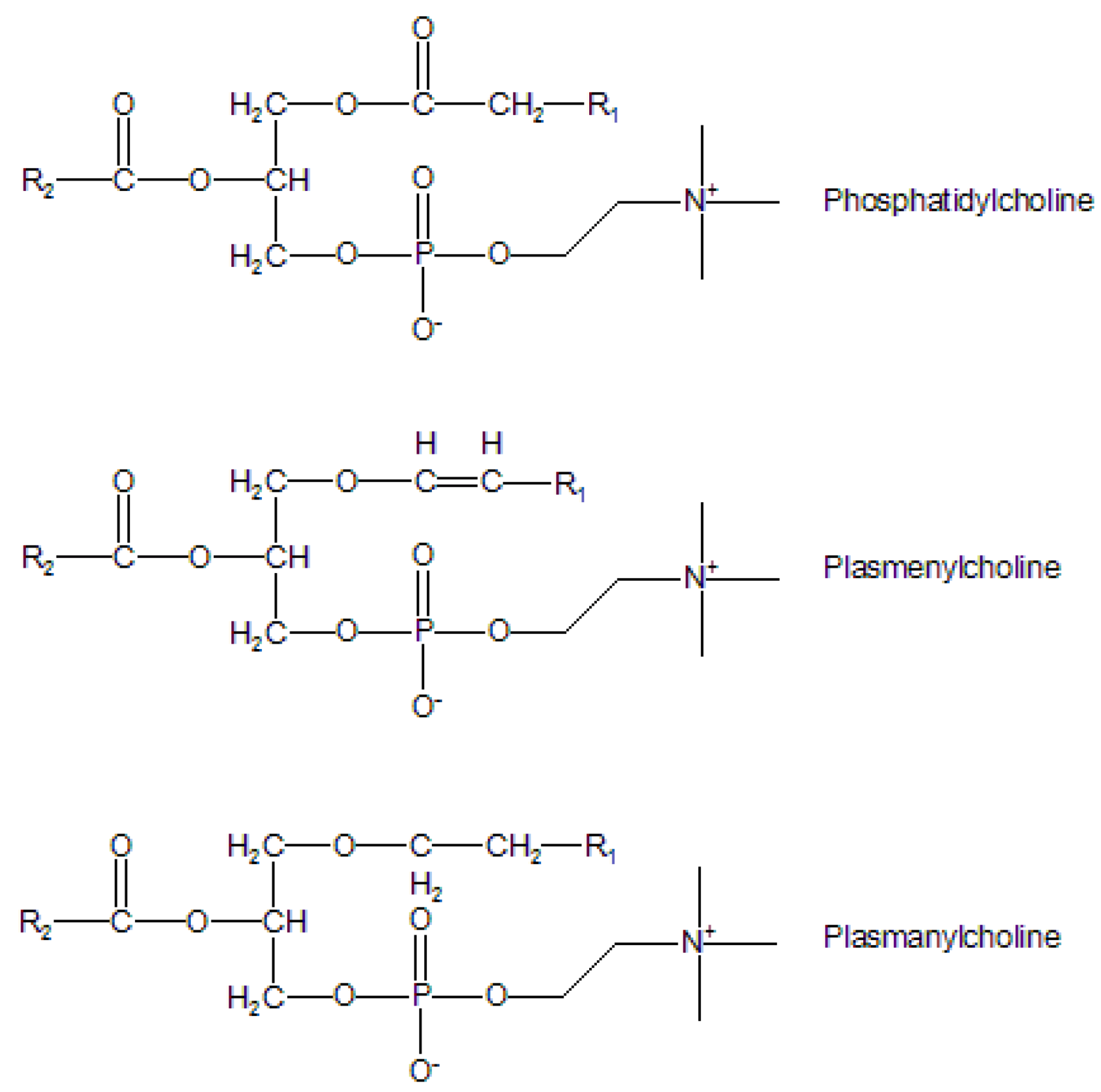
1. Introduction
Lipids: A complex world


2. Detection of Phospholipids: An Analytical Challenge
2.1. Pre-column derivatization of phospholipids
2.2. Fluorescence probe assisted post-column detection of phospholipids
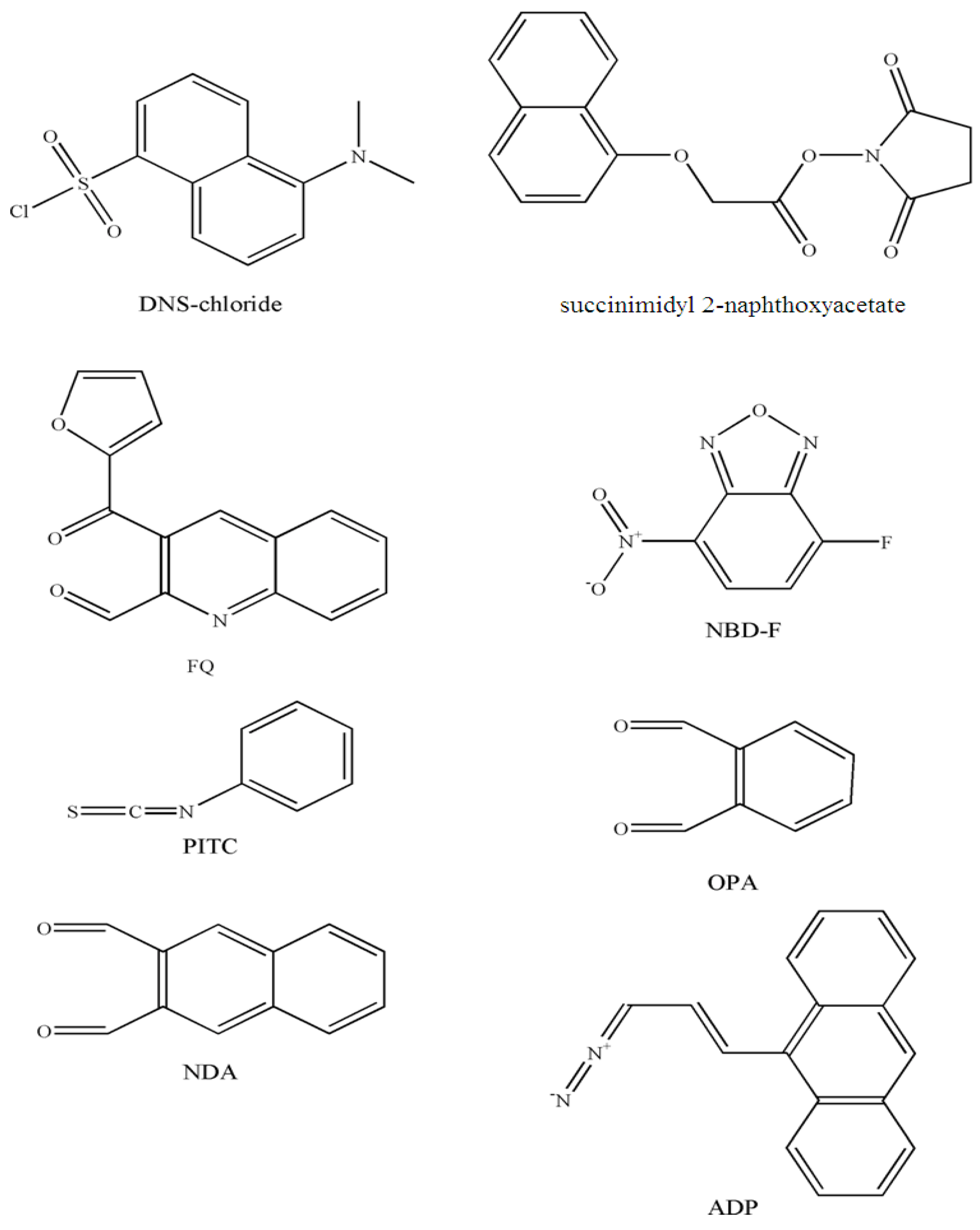
3. Pre-Column Fluorescent Covalent Derivatization of Phospholipids
3.1. Derivatization without phospholipid hydrolysis
3.1.1. Amino phospholipids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Matrices | Derivatizing reagent | Separation Type | Temperature condition (°C) | Reaction time | λex-λem (nm) | Limit of detection | Refs. |
|---|---|---|---|---|---|---|---|---|
| Phospholipids with amino group | ||||||||
| GPEtn, lyso-GPEtn, GPSer, lyso-GPSer | Rat brain | DNS-chloride | HPLC | 50 | 3h | 342-500 | 20 pmol | [29] |
| GPEtn | Egg | DNS-chloride | HPLC | 50 | 2-3h | 360-> 420 | - | [30,31] |
| GPEtn, lyso- GPEtn, GPSer, lyso- GPSer | Rat brain | succinimidyl 2-naphtoxyacetate | HPLC | Room temperature | 2h | 228-342 | 2 pmol | [32] |
| GPEtn, lyso- GPEtn, PS, lysoPS | - | FQ | MECC | 55 | 15min | 488-630 | 0.65-1.1 fg | [12] |
| PE, lysoPE, PS, lysoPS | - | NBD-F | MECC | 55°C | 5min | 488-535 | 0.18-0.87 fg | [12] |
| Sphingoid base 1-phosphates | Human plasma, serum, platelets | OPA | HPLC | Room temperature | 20 min | 340-455 | < 5 pmol | [33] |
| Sphingosine 1-phosphate | Human plasma | OPA | HPLC | Room temperature | 20 min | 340-455 | < 50 ng/mL | [34] |
| Sphingosine 1-phosphate | Plasma (human, horse, mouse), mousse tissues | NDA | HPLC | 50°C | 10 min | 252-483 | 20.9 fmol | [35] |
| Phosphatidic acid | ||||||||
| GPA | Egg | DNS-ethanolamine from DNS-Cl | HPLC | Room temperature | 2-3h | 360-420 | - | [31] |
| GPA | Rabbit platelets | 3-(9 anthroyl) diazo-2-propene | HPLC | 4°C | 8h | 254-430 | 0.05ng/mL | [36] |

3.1.2. Phosphatidic acid
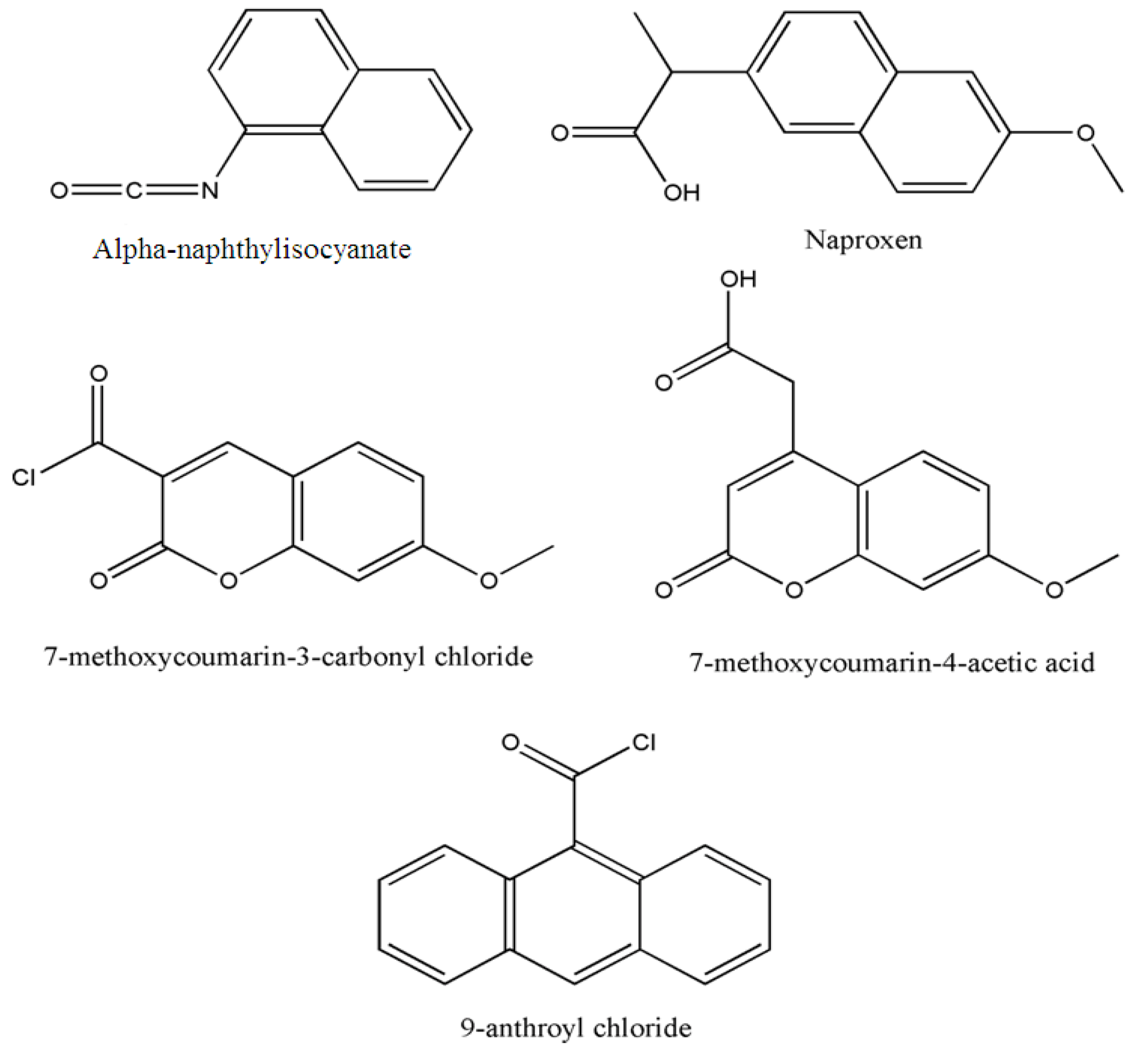
3.2. Derivatization after phospholipid hydrolysis
3.2.1. Diacylglycerols obtained after hydrolysis by phospholipase C
| Analyte | Derivatizing reagent | Matrices | Separation | Temperature condition (°C) | Reaction time | λex- λem (nm) | Limit of detection | Refs. |
|---|---|---|---|---|---|---|---|---|
| Hydrolysis by phospholipase C | ||||||||
| GPCho | alpha-naphthylisocyanate | Rat liver microsomes | HPLC | 85 | 2 h | 280-360 | 10 pmol | [39] |
| GPCho, GPEtn | DNS-ethanolamine phosphate from DNS-Cl | Egg | HPLC | 60-80 | 24 h | 360-420 | - | [30] |
| GPEtn | Naproxen chloride | rat brain cerebrum and cerebellum | HPLC | 80 | 15 min | 332-352 | 1 pmol | [40] |
| GPCho, GPA | 9-anthroyl chloride | Rat thymocytes | HPLC | 60 | 10 min | 360-460 | 0.1 pmol | [41] |
| PAF | 7-methoxycoumarin-3-carbonyl chloride or 7-methoxycoumarin-4- acetic acid | Human leucocytes | HPLC | - | - | - | 100 pg | [42] |
| Hydrolysis by phospholipase D | ||||||||
| GPCho | DNS-ethanolamine from DNS-Cl | Egg | HPLC | Room temperature | 2-3 h | 360-420 | - | [31] |
| Hydrolysis by alkaline phosphatase | ||||||||
| Sphingoid base 1-phosphates | OPA | Serum, cultured cells, rat tissues | HPLC | Room temperature | 20 min | 340-455 | 0.5 pmol | [43] |
| Sphingosine 1-phosphate and dihydro Sphingosine 1-phosphate | OPA | Cultured cells, plasma | HPLC | Room temperature | 20 min | 340-455 | <0.5 pmol | [44] |

3.2.2. Phosphatidic acid obtained after hydrolysis by phospholipase D
3.2.3. Sphingoid base obtained after hydrolysis by alkaline phosphatase
4. Probe Assisted Fluorescence Detection after Native Separation of Phospholipids
4.1. Fluorescence detection after TLC
4.2. Post-column detection after HPLC

4.3. Surfactant
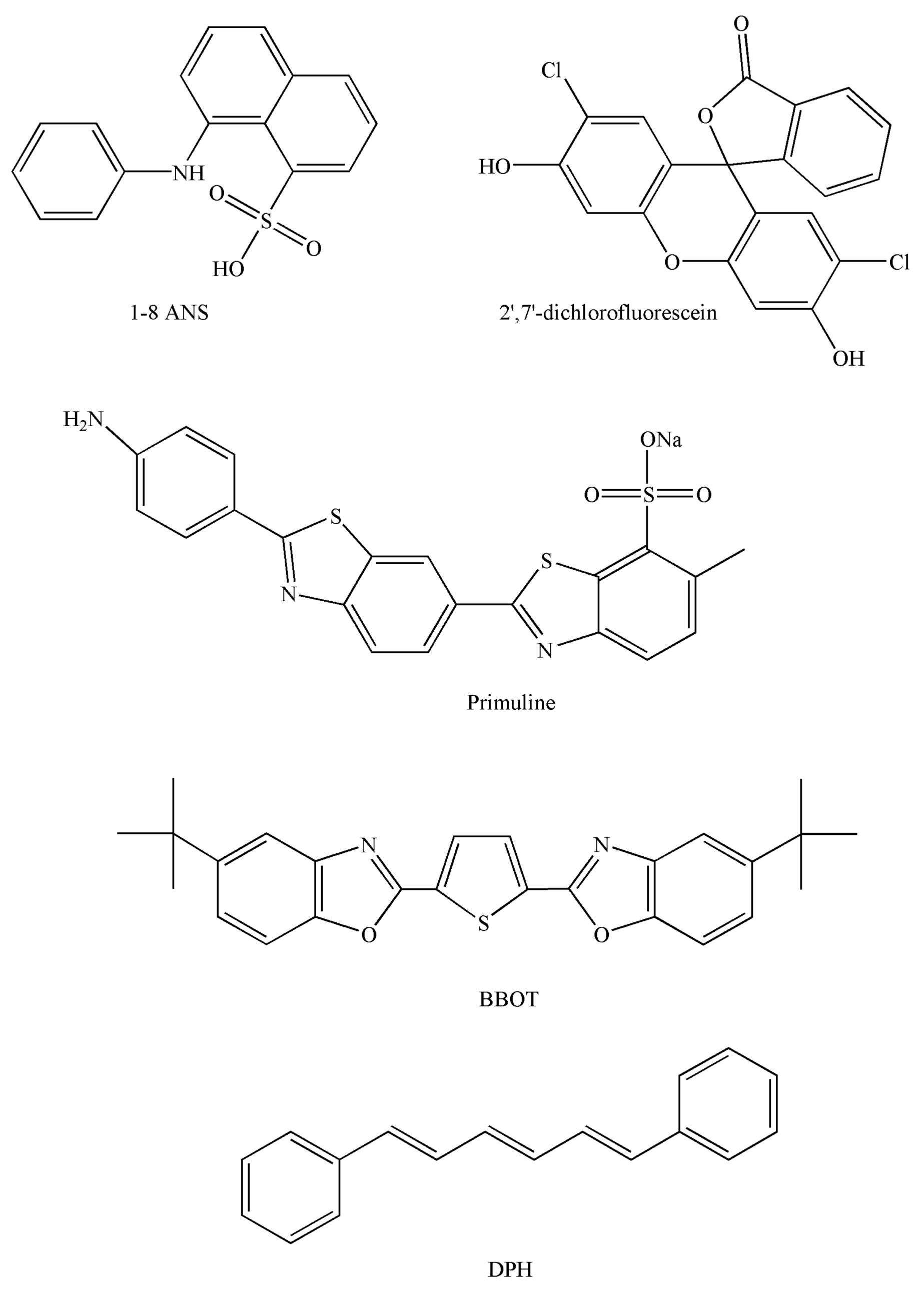
4.4. Probes
| Refs. | [51] | [53] | [61] | [62] | [63] | [64] | [54] |
|---|---|---|---|---|---|---|---|
| Molecular probe | BBOT | DPH | DPH | DPH | DPH | DPH | DPH |
| Concentration before | 0.023 | 0.15 | 0.45 | 0.00045 | 0.45 | 10.5 | 3.35 |
| (and after) mixing (µmol/L) | -0.006 | -0.13 | -0.34 | -0.00025 | -0.37 | -7 | -2.79 |
| λex-λem (nm) | 372–440 | 365–460 | 340–460 | 340–460 | 340–460 | 340–460 | 350–430 |
| Surfactant | - | - | Tween 20 | Tween 20 | Brij 35 | Brij 35 | Brij 35 |
| Concentration (v/v) | 0.00% | 0.00% | 0.03% | 0.03% | 0.02% | ||
| Flow (mL.min-1) | |||||||
| - Chromatographic | 0.8 | 1 | 1 | 1 | 1 | 1 | 0.1 |
| - Post-column phase | 0.25 | 6 | 3 | 1,2 | 4.5 | 2 | 0.5 |
| % aqueous phase | 1.5 | 87.9 | 75 | 54.5 | 82.4 | 66.7 | 83.3 |
| Tubing L(m) × id(mm) | 0.3 × 0.25 | - | 3 × 0.5 | 0.3 × 0.5 | 2 × 0.8 | 3 × 0.25 | 1.4 × 0.5 |
| Time (s) | <1 | 60 | 9 | 2 | 11 | 3 | 27 |
| Temperature (°C) | - | 40 | 50 | 50 | 50 | 50 | 35 |
4.5. Post-column flow rate
4.6. Post column tubing length (or reaction time)
4.7. Post-column system temperature
4.8. Advantage of micro-LC with post-column device
5. Conclusions
- Sample Availability: Not available.
References
- Spector, A.A.; Yorek, M.A. Membrane lipid composition and cellular function. J. Lipid. Res. 1985, 26, 1015–1035. [Google Scholar]
- Ghosh, S.; Strum, J.C.; Bell, R.M. Lipid biochemistry: functions of glycerolipids and sphingolipids in cellular signaling. Faseb. J. 1997, 11, 45–50. [Google Scholar]
- Shimizu, T. Lipid mediators in health and disease: enzymes and receptors as therapeutic targets for the regulation of immunity and inflammation. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 123–150. [Google Scholar] [CrossRef]
- Roberts, J.L. I, II Lipids as regulators of cell functions. Cell Mol. Life Sci. 2002, 59, 727–728. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Shotgun lipidomics: Electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass. Spectrom. Rev. 2005, 24, 367–412. [Google Scholar] [CrossRef]
- Mueller, H.W.; O'Flaherty, J.T.; Greene, D.G.; Samuel, M.P. 1-O-alkyl-linked glycerophospholipids of human neutrophils: Distribution of arachidonate and other acyl residues in the ether-linked and diacyl species. J. Lipid. Res. 1984, 25, 383–388. [Google Scholar]
- Scherrer, L.A.; Gross, R.W. Subcellular distribution, molecular dynamics and catabolism of plasmalogens in myocardium. Mol. Cell. Biochem. 1989, 88, 97–105. [Google Scholar]
- Schiller, J.; Suss, R.; Arnhold, J.; Fuchs, B.; Lessig, J.; Muller, M.; Petkovic, M.; Spalteholz, H.; Zschornig, O.; Arnold, K. Matrix-assisted laser desorption and ionization time-of-flight (MALDI-TOF) mass spectrometry in lipid and phospholipid research. Prog. Lipid. Res. 2004, 43, 449–488. [Google Scholar] [CrossRef]
- Daleke, D.L. Regulation of transbilayer plasma membrane phospholipid asymmetry. J. Lipid. Res. 2003, 44, 233–242. [Google Scholar] [CrossRef]
- Abidi, S.L.; Mounts, T.L. High-performance liquid chromatography of phosphatidic acid. J. Chromatogr. B 1995, 671, 281–297. [Google Scholar] [CrossRef]
- Gao, F.; Dong, J.; Li, W.; Wang, T.; Liao, J.; Liao, Y.; Liu, H. Separation of phospholipids by capillary zone electrophoresis with indirect ultraviolet detection. J. Chromatogr. A 2006, 1130, 259–264. [Google Scholar] [CrossRef]
- Zhang, L.; Krylov, S.N.; Hu, S.; Dovichi, N.J. Methyl-beta-cyclodextrin modified micellar electrokinetic capillary chromatography with laser-induced fluorescence for separation and detection of phospholipids. J. Chromatogr. A 2000, 894, 129–134. [Google Scholar] [CrossRef]
- Myher, J.J.; Kuksis, A. General strategies in chromatographic analysis of lipids. J. Chromatogr. B 1995, 671, 3–33. [Google Scholar] [CrossRef]
- Nissen, H.P.; Kreysel, H.W. Analysis of phospholipids in human semen by high-performance liquid chromatography. J. Chromatogr. 1983, 276, 29–35. [Google Scholar] [CrossRef]
- Pulfer, M.; Murphy, R.C. Electrospray mass spectrometry of phospholipids. Mass. Spectrom. Rev. 2003, 22, 332–364. [Google Scholar] [CrossRef]
- Murphy, E.J.; Rosenberger, T.A.; Horrocks, L.A. Separation of neutral lipids by high-performance liquid chromatography: quantification by ultraviolet, light scattering and fluorescence detection. J. Chromatogr. B 1996, 685, 9–14. [Google Scholar] [CrossRef]
- Hoving, E.B. Chromatographic methods in the analysis of cholesterol and related lipids. J. Chromatogr. B 1995, 671, 341–362. [Google Scholar] [CrossRef]
- Peterson, B.L.; Cummings, B.S. A review of chromatographic methods for the assessment of phospholipids in biological samples. Biomed. Chromatogr. 2006, 20, 227–243. [Google Scholar] [CrossRef]
- Stith, B.J.; Hall, J.; Ayres, P.; Waggoner, L.; Moore, J.D.; Shaw, W.A. Quantification of major classes of Xenopus phospholipids by high performance liquid chromatography with evaporative light scattering detection. J. Lipid. Res. 2000, 41, 1448–1454. [Google Scholar]
- Jungalwala, F.B.; Evans, J.E.; McCluer, R.H. High-performance liquid chromatography of phosphatidylcholine and sphingomyelin with detection in the region of 200 nm. Biochem. J. 1976, 155, 55–60. [Google Scholar]
- Patton, G.M.; Fasulo, J.M.; Robins, S.J. Separation of phospholipids and individual molecular species of phospholipids by high-performance liquid chromatography. J. Lipid. Res. 1982, 23, 190–196. [Google Scholar]
- Abidi, S.L.; Mounts, T.L. Reversed-phase separations of nitrogenous phospholipids on an octadecanoyl poly(vinyl alcohol) phase. J. Chromatogr. A 1997, 773, 93–101. [Google Scholar] [CrossRef]
- Grizard, G.; Sion, B.; Bauchart, D.; Boucher, D. Separation and quantification of cholesterol and major phospholipid classes in human semen by high-performance liquid chromatography and light-scattering detection. J. Chromatogr. B 2000, 740, 101–107. [Google Scholar] [CrossRef]
- Ramos, R.G.; Libong, D.; Rakotomanga, M.; Gaudin, K.; Loiseau, P.M.; Chaminade, P. Comparison between charged aerosol detection and light scattering detection for the analysis of Leishmania membrane phospholipids. J. Chromatogr. A 2008, 1209, 88–94. [Google Scholar] [CrossRef]
- Vervoort, N.; Daemen, D.; Torok, G. Performance evaluation of evaporative light scattering detection and charged aerosol detection in reversed phase liquid chromatography. J. Chromatogr. A 2008, 1189, 92–100. [Google Scholar] [CrossRef]
- Fukushima, T.; Usui, N.; Santa, T.; Imai, K. Recent progress in derivatization methods for LC and CE analysis. J. Pharm. Biomed. Anal. 2003, 30, 1655–1687. [Google Scholar] [CrossRef]
- Lingeman, H.; Underberg, W.J.M.; Takadate, A.; Hulshoff, A. Fluorescence detection in High-performance liquid chromatography. J. Liq. Chromatogr. 1985, 8, 789–874. [Google Scholar] [CrossRef]
- Wang, Y.; Krull, I.S.; Liu, C.; Orr, J.D. Derivatization of phospholipids. J. Chromatogr. B 2003, 793, 3–14. [Google Scholar] [CrossRef]
- Chen, S.S.; Kou, A.Y.; Chen, H.H. Measurement of ethanolamine- and serine-containing phospholipids by high-performance liquid chromatography with fluorescence detection of their Dns derivatives. J. Chromatogr. 1981, 208, 339–346. [Google Scholar] [CrossRef]
- Ryan, P.J.; Honeyman, T.W. Formation and separation of fluorescent derivatives of diacylglycerols. J. Chromatogr. 1995, 331, 177–182. [Google Scholar]
- Ryan, P.J.; McGoldrick, K.; Stickney, D.; Honeyman, T.W. Formation and separation of fluorescent derivatives of phosphatidic acid. J. Chromatogr. 1995, 320, 421–425. [Google Scholar]
- Chen, S.S.; Kou, A.Y.; Chen, H.H. Quantitative analysis of aminophospholipids by high-performance liquid chromatography using succinimidyl 2-naphthoxyacetate as a fluorescent label. J. Chromatogr. 1983, 276, 37–44. [Google Scholar] [CrossRef]
- Caligan, T.B.; Peters, K.; Ou, J.; Wang, E.; Saba, J.; Merrill, A.H., Jr. A high-performance liquid chromatographic method to measure sphingosine 1-phosphate and related compounds from sphingosine kinase assays and other biological samples. Anal. Biochem. 2000, 281, 36–44. [Google Scholar]
- Butter, J.J.; Koopmans, R.P.; Michel, M.C. A rapid and validated HPLC method to quantify sphingosine 1-phosphate in human plasma using solid-phase extraction followed by derivatization with fluorescence detection. J. Chromatogr. B 2005, 824, 65–70. [Google Scholar] [CrossRef]
- He, X.; Huang, C.L.; Schuchman, E.H. Quantitative analysis of sphingosine-1-phosphate by HPLC after napthalene-2,3-dicarboxaldehyde (NDA) derivatization. J. Chromatogr. B 2009, 877, 983–990. [Google Scholar] [CrossRef]
- Yamada, K.; Abe, S.; Katayama, K.; Sato, T. Sensitive high-performance liquid chromatographic method for the determination of phosphatidic acid. J. Chromatogr. 1988, 424, 367–372. [Google Scholar] [CrossRef]
- Haddadian, E.; Shamsi, S.A.; Schaeper, J.P.; Danielson, N.D. Capillary electrophoresis of phospholipids with indirect photometric detection. J. Chromatogr. Sci. 1998, 36, 395–400. [Google Scholar]
- Stark, M.; Johansson, J. Determination of proteins, phosphatidylethanolamine, and phosphatidylserine in lipid-rich materials by analysis of phenylthiocarbamyl derivative. Methods Mol. Biol. 2000, 159, 177–182. [Google Scholar]
- Kruger, J.; Rabe, H.; Reichmann, G.; Rustow, B. Separation and determination of diacylglycerols as their naphthylurethanes by high-performance liquid chromatography. J. Chromatogr. 1984, 307, 387–392. [Google Scholar] [CrossRef]
- Rastegar, A.; Pelletier, A.; Duportail, G.; Freysz, L.; Leray, C. Sensitive analysis of phospholipid molecular species by high-performance liquid chromatography using fluorescent naproxen derivatives of diacylglycerols. J. Chromatogr. 1990, 518, 157–165. [Google Scholar] [CrossRef]
- Thevenon, C.; El-Bawab, S.; Chantegrel, B.; Lagarde, M. Highly sensitive measurement of lipid molecular species from biological samples by fluorimetric detection coupled to high-performance liquid chromatography. J. Chromatogr. B 1998, 708, 39–47. [Google Scholar] [CrossRef]
- Mita, H.; Yasueda, H.; Hayakawa, T.; Shida, T. Quantitation of platelet-activating factor by high-performance liquid chromatography with fluorescent detection. Anal. Biochem. 1989, 180, 131–135. [Google Scholar]
- Min, J.K.; Yoo, H.S.; Lee, E.Y.; Lee, W.J.; Lee, Y.M. Simultaneous quantitative analysis of sphingoid base 1-phosphates in biological samples by o-phthalaldehyde precolumn derivatization after dephosphorylation with alkaline phosphatase. Anal. Biochem. 2002, 303, 167–175. [Google Scholar]
- Lee, Y.M.; Venkataraman, K.; Hwang, S.I.; Han, D.K.; Hla, T. A novel method to quantify sphingosine 1-phosphate by immobilized metal affinity chromatography (IMAC). Prostaglandins Other Lipid Mediat. 2007, 84, 154–162. [Google Scholar] [CrossRef]
- Takamura, H.; Kito, M. A highly sensitive method for quantitative analysis of phospholipid molecular species by high-performance liquid chromatography. J. Biochem. 1991, 109, 436–439. [Google Scholar]
- Kraus, R.; Wuthe, J.; Rufer, R. Method for determination of different phospholipids by circular high-performance thin-layer chromatography. J. Chromatogr. 1987, 413, 257–263. [Google Scholar] [CrossRef]
- Moe, M.K.; Anderssen, T.; Strom, M.B.; Jensen, E. Vicinal hydroxylation of unsaturated fatty acids for structural characterization of intact neutral phospholipids by negative electrospray ionization tandem quadrupole mass spectrometry. Rapid. Commun. Mass. Spectrom. 2004, 18, 2121–2130. [Google Scholar] [CrossRef]
- Moe, M.K.; Anderssen, T.; Strom, M.B.; Jensen, E. Total structure characterization of unsaturated acidic phospholipids provided by vicinal di-hydroxylation of fatty acid double bonds and negative electrospray ionization mass spectrometry. J. Am. Soc. Mass. Spectrom. 2005, 16, 46–59. [Google Scholar] [CrossRef]
- White, T.; Bursten, S.; Federighi, D.; Lewis, R.A.; Nudelman, E. High-resolution separation and quantification of neutral lipid and phospholipid species in mammalian cells and sera by multi-one-dimensional thin-layer chromatography. Anal. Biochem. 1998, 258, 109–117. [Google Scholar]
- Richter, G.; Schober, C.; Suss, R.; Fuchs, B.; Muller, M.; Schiller, J. The reaction between phosphatidylethanolamines and HOCl investigated by TLC: fading of the dye primuline is induced by dichloramines. J. Chromatogr. B 2008, 867, 233–237. [Google Scholar] [CrossRef]
- Ouhazza, M.; Siouffi, A.M. Liquid chromatography analysis of some phospholipids with fluorescence detection. Analusis 1992, 20, 185–188. [Google Scholar]
- Haugland, R.P. Handbook of Fluorescent Probes and Research Chemicals, 6th ed; M.T.Z Spence: Eugene, OR, USA, 1996. [Google Scholar]
- Gebhardt, D.O.; Soederhuizen, W.; Feyen, J.H. The fluorimetric determination of the lecithin/sphingomyelin ratio of amniotic fluid after HPLC. Ann. Clin. Biochem. 1985, 22, 321–323. [Google Scholar]
- Caudron, E.; Zhou, J.Y.; Chaminade, P.; Baillet, A.; Prognon, P. Fluorescence probe assisted post-column detection for lipid analysis in microbore-LC. J. Chromatogr. A 2005, 1072, 149–157. [Google Scholar] [CrossRef]
- Shinitzky, M.; Barenholz, Y. Fluidity parameters of lipid regions determined by fluorescence polarization. Biochim. Biophys. Acta 1978, 515, 367–394. [Google Scholar] [CrossRef]
- Lentz, B.R.; Barenholz, Y.; Thompson, T.E. Fluorescence depolarization studies of phase transitions and fluidity in phospholipid bilayers. 1. Single component phosphatidylcholine liposomes. Biochemistry 1976, 15, 4521–4528. [Google Scholar] [CrossRef]
- Lentz, B.R.; Barenholz, Y.; Thompson, T.E. Fluorescence depolarization studies of phase transitions and fluidity in phospholipid bilayers. 2 Two-component phosphatidylcholine liposomes. Biochemistry 1976, 15, 4529–4537. [Google Scholar] [CrossRef]
- Repakova, J.; Capkova, P.; Studenovsky, M.; Ilavsky, M. Characterization of molecular structures and properties of polyurethanes using molecular dynamics simulations. J. Mol. Model. 2004, 10, 240–249. [Google Scholar] [CrossRef]
- London, E.; Feligenson, G.W. A convenient and sensitive fluorescence assay for phospholipid vesicles using diphenylhexatriene. Anal. Biochem. 1978, 88, 203–211. [Google Scholar]
- Hernando, V.; Zhou, J.Y.; Nouaille-Degorce, B.; Prognon, P.; Brion, F.; Rieutord, A. Quantitative determination of fat emulsion using 1,6-diphenyl-1,3,5-hexatriene as fluorescence probe: application to the compounding of all-in-one parenteral nutrition admixtures. J. Pharm. Biomed. Anal. 2001, 26, 487–493. [Google Scholar] [CrossRef]
- Postle, A.D. Method for the sensitive analysis of individual molecular species of phosphatidylcholine by high-performance liquid chromatography using post-column fluorescence detection. J. Chromatogr. 1987, 415, 241–251. [Google Scholar] [CrossRef]
- Kitsos, M.; Gandini, C.; Massolini, G.; De Lorenzi, E.; Caccialanza, G. High-performance liquid chromatography post-column derivatization with fluorescence detection to study the influence of ambroxol on dipalmitoylphosphatidylcholine levels in rabbit eustachian tube washings. J. Chromatogr. 1991, 553, 1–6. [Google Scholar] [CrossRef]
- Bernhard, W.; Linck, M.; Creutzburg, H.; Postle, A.D.; Arning, A.; Martin-Carrera, I.; Sewing, K.F. High-performance liquid chromatographic analysis of phospholipids from different sources with combined fluorescence and ultraviolet detection. Anal. Biochem. 1994, 220, 172–180. [Google Scholar]
- Hradec, J.; Dufek, P.; Matucha, M. Quantitation of individual molecular species of phosphatidylcholines by reversed-phase high-performance liquid chromatography with fluorometric detection. J. Chromatogr. B 1998, 714, 145–151. [Google Scholar] [CrossRef]
- Caudron, E.; Zhou, J.Y.; England, P.; Ollivon, M.; Prognon, P. Some insights about 1,6-diphenyl-1,3,5-hexatriene-lipid supramolecular assemblies by steady-state fluorescence measurements. Appl. Spectrosc. 2007, 61, 963–969. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Chaminade, P.; Gaudin, K.; Prognon, P.; Baillet, A.; Ferrier, D. Postcolumn fluorescence as an alternative to evaporative light scattering detection for ceramide analysis with gradient elution in non-aqueous reversed-phase liquid chromatography. J. Chromatogr. A 1999, 859, 99–105. [Google Scholar] [CrossRef]
- Novotny, M. Recent advances in microcolumn liquid chromatography. Anal. Chem. 1988, 60, 500–510. [Google Scholar]
- Menet, H.; Gareil, P.; Caude, M.; Rosset, R. Microchromatographie en phase liquide. Analusis 1983, 11, 141–155. [Google Scholar]
- Vissers, J.P. Recent developments in microcolumn liquid chromatography. J. Chromatogr. A 1999, 856, 117–143. [Google Scholar] [CrossRef]
- IUPAC-IUB Commission on biochemical nomenclature. The nomenclature of lipids. Biochem. J. 1978, 171, 21–35.
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ibrahim, H.; Caudron, E.; Kasselouri, A.; Prognon, P. Interest of Fluorescence Derivatization and Fluorescence Probe Assisted Post-column Detection of Phospholipids: A Short Review. Molecules 2010, 15, 352-373. https://doi.org/10.3390/molecules15010352
Ibrahim H, Caudron E, Kasselouri A, Prognon P. Interest of Fluorescence Derivatization and Fluorescence Probe Assisted Post-column Detection of Phospholipids: A Short Review. Molecules. 2010; 15(1):352-373. https://doi.org/10.3390/molecules15010352
Chicago/Turabian StyleIbrahim, Hanadi, Eric Caudron, Athena Kasselouri, and Pratrice Prognon. 2010. "Interest of Fluorescence Derivatization and Fluorescence Probe Assisted Post-column Detection of Phospholipids: A Short Review" Molecules 15, no. 1: 352-373. https://doi.org/10.3390/molecules15010352
APA StyleIbrahim, H., Caudron, E., Kasselouri, A., & Prognon, P. (2010). Interest of Fluorescence Derivatization and Fluorescence Probe Assisted Post-column Detection of Phospholipids: A Short Review. Molecules, 15(1), 352-373. https://doi.org/10.3390/molecules15010352