Seasonal Variation of Bioactive Alkaloid Contents in Macleaya microcarpa (Maxim.) Fedde

Abstract
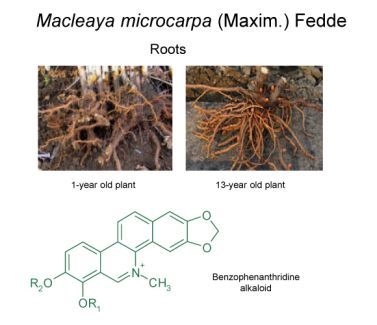
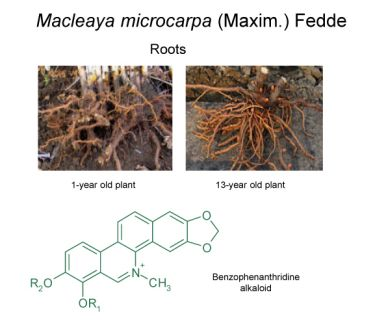
:
1. Introduction
2. Results and Discussion
2.1. Analyses of aerial parts

{kind=link}
{kind=link}
| Year of harvest | Age of the plant | PRO (n = 3) | ALL (n = 3) | SA (n = 3) | CHE (n = 3) | ||||
|---|---|---|---|---|---|---|---|---|---|
| mg/g ± SD | % | mg/g ± SD | % | mg/g ± SD | % | mg/g ± SD | % | ||
| 2008 | 1 | 1.559 ± 0.056 | 9.0 | 5.655 ± 0.239 | 32.6 | 4.370 ± 0.225 | 25.2 | 5.745 ± 0.199 | 33.2 |
| 2009 | 1 | 2.814 ± 0.051 | 18.0 | 6.500 ± 0.093 | 41.1 | 3.115 ± 0.107 | 19.7 | 3.371 ± 0.148 | 21.2 |
| 2010 | 1 | 2.610 ± 0.185 | 18.4 | 6.224 ± 0.620 | 43,8 | 2.433 ± 0.468 | 17.1 | 2.927 ± 0.239 | 20.7 |
| 2010 | 2 | 1.123 ± 0.053 | 9.8 | 3.815 ± 0.157 | 33.2 | 2.812 ± 0.316 | 22.4 | 3.757 ± 0.062 | 34.6 |
| 2009 | 12 | 2.698 ± 0.050 | 16.1 | 7.286 ± 0.140 | 43.5 | 2.961 ± 0.154 | 17.7 | 3.798 ± 0,142 | 22.7 |
| 2010 | 13 | 5.131 ± 0.030 | 20.1 | 13.604 ± 0.184 | 53.4 | 2.948 ± 0.212 | 11.6 | 3.678 ± 0.056 | 14.5 |
| Month | PRO (n = 3) | ALL (n = 3) | SA (n = 3) | CHE (n = 3) | ||||
|---|---|---|---|---|---|---|---|---|
| mg/g ± SD | % | mg/g ± SD | % | mg/g ± SD | % | mg/g ± SD | % | |
| April | 1.050 ± 0.070 | 4.2 | 3.040 ± 0.185 | 1.2 | 10.135 ± 1.726 | 40.6 | 10.767 ± 1.279 | 43.1 |
| May | 1.016 ± 0.088 | 4.8 | 2.789 ± 0.287 | 13.2 | 7.463 ± 0.855 | 35.3 | 9.877 ± 1.059 | 46.7 |
| June | 2.341 ± 0.083 | 10.0 | 7.225 ± 0.148 | 31.1 | 5.439 ± 0.628 | 23.4 | 8.248 ± 0.347 | 35.5 |
| July | 5.830 ± 0.721 | 16.7 | 18.229 ± 2.337 | 52.5 | 4.266 ± 0.512 | 12.3 | 6.384 ± 0.981 | 18.4 |
| August | 2.553 ± 0.098 | 13.4 | 8.134 ± 0.033 | 42.7 | 3.707 ± 0.082 | 19.5 | 4.654 ± 0.257 | 24.4 |
| September | 0.872 ± 0.228 | 3.7 | 14.643 ± 0.639 | 61.9 | 3.436 ± 0.159 | 14.5 | 4.700 ± 0.175 | 19.9 |
| October | 5.131 ± 0.030 | 20.1 | 13.604 ± 0.184 | 53.4 | 2.948 ± 0.212 | 11.6 | 3.678 ± 0.056 | 14.5 |
2.2. Analyses of roots
| Year of harvest | Age of the plant | PRO (n = 3) | ALL (n = 3) | SA (n = 3) | SR (n = 3) | CHE (n = 3) | CR (n = 3) | CL (n = 3) | MA (n = 3) |
|---|---|---|---|---|---|---|---|---|---|
| mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | ||
| 2008 | 1 | 11.955 ± 3.700 | 48.933 ± 1.572 | 1.791 ± 0.124 | 0.041 ± 0.011 | 2.798 ± 0.043 | 0.764 ± 0.102 | 0.179 ± 0.003 | 0.130 ± 0.013 |
| 2009 | 1 | 11.877 ± 3.454 | 44.806 ± 5.404 | 1.679± 0.423 | 0.043 ± 0.022 | 2.595 ± 0.013 | 0.764 ± 0.128 | 0.227 ± 0.016 | 0.194 ± 0.029 |
| 2010 | 1 | 12.269 ± 3.706 | 32.280 ± 9.697 | 3.028 ± 0.793 | 0.031 ± 0.001 | 4.971 ± 1.440 | 1.090 ± 0.284 | 0.181 ± 0.064 | 0.153 ± 0.007 |
| 2010 | 2 | 12.910 ± 0.532 | 41.991 ± 2.435 | 2.587 ± 0.107 | 0.047 ± 0.005 | 3.857 ± 0.238 | 1.235 ± 0.040 | 0.172 ± 0.003 | 0.134 ± 0.015 |
| 2009 | 12 | 7.567 ± 1.173 | 24.207 ± 0.632 | 1.955 ± 0.462 | 0.254 ± 0.005 | 3.027 ± 0.268 | 3.669 ± 0.348 | 1.501 ± 0.008 | 3.021 ± 0.328 |
| 2010 | 13 | 8.958 ± 0.348 | 34.947 ± 1.399 | 3.106 ± 0.570 | 0.140 ± 0.019 | 4.702 ± 0.469 | 2.720 ± 0.642 | 0.883 ± 0.197 | 1.340 ± 0.413 |
| Month | PRO (n = 3) | ALL (n = 3) | SA (n = 3) | SR (n = 3) | CHE (n = 3) | CR (n = 3) | CL (n = 3) | MA (n = 3) |
|---|---|---|---|---|---|---|---|---|
| mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | mg/g ± SD | |
| April | 11.843 ± 0.536 | 47.810 ± 2.558 | 2.216 ± 0.075 | 0.098 ± 0.009 | 4.573 ± 0.196 | 2.878 ± 0,097 | 0.735 ± 0.072 | 0.908 ± 0.153 |
| May | 12.999 ± 0.283 | 43.691 ± 0.235 | 3.268 ± 0.107 | 0.107 ± 0.002 | 6.093 ± 0.041 | 2.115 ± 0.041 | 0.650 ± 0.051 | 0.764 ± 0.063 |
| June | 12.686 ± 1.115 | 61.848 ± 4.725 | 4.290 ± 0.308 | 0.221 ± 0.036 | 9.854 ± 0.939 | 5.376 ± 0.939 | 1.134 ± 0.052 | 1.634 ± 0.263 |
| July | 11.754 ± 0.353 | 66.116 ± 1.421 | 5.583 ± 1.381 | 0.289 ± 0.027 | 15.052 ± 1.381 | 5.038 ± 0.155 | 1.783 ± 0.051 | 2.475 ± 0.192 |
| August | 9.256 ± 0.380 | 42.748 ± 1.983 | 2.789 ± 0.176 | 0.207 ± 0.016 | 5.600 ± 0.362 | 2.244 ± 0.187 | 1.133 ± 0.071 | 2.466 ± 0.053 |
| September | 6.676 ± 0.622 | 34.001 ± 2.477 | 2.941 ± 0.252 | 0.193 ± 0.050 | 6.936 ± 0.764 | 3.303 ± 0.723 | 1.520 ± 0.316 | 2. 311 ± 0.672 |
| October | 8.958 ± 0.348 | 34.947 ± 1.399 | 3.106 ± 0.570 | 0.140 ± 0.019 | 4.702 ± 0.469 | 2.720 ± 0.642 | 0.883 ± 0.197 | 1.340 ± 0.413 |
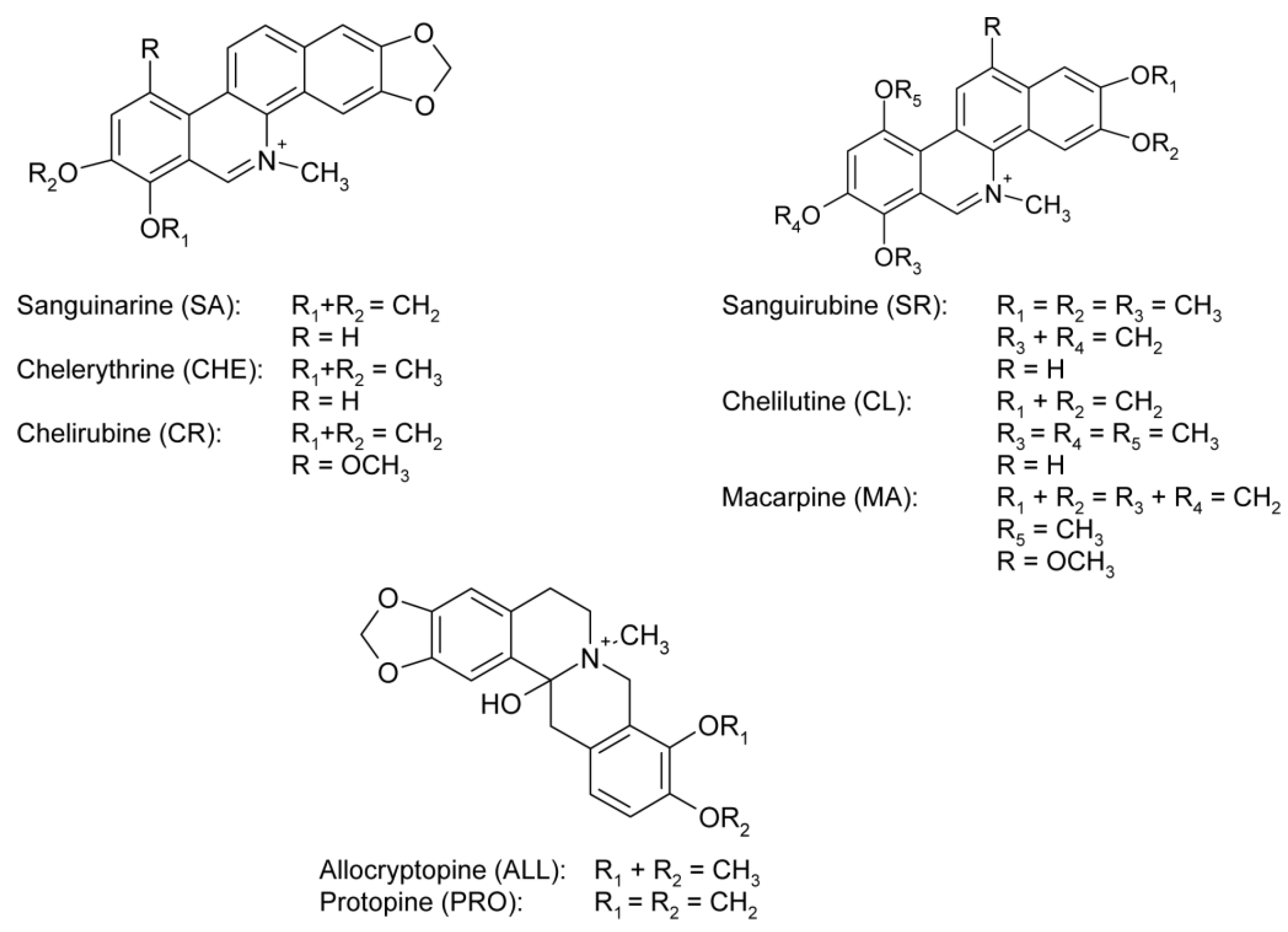
2.3. Alkaloids of the plant

3. Experimental
3.1. General
3.2. Alkaloids and chemicals
3.3. Plant material and cultivated technique
3.4. Extract preparation
4. Conclusions
Acknowledgments
References and Notes
- Slavík, J.; Slavíková, L. Papaveraceae alkaloids VI. On alkaloids of Macleaya microcarpa (Maxim) Fedde. Chem. Listy 1954, 48, 106–110, in Czech.. [Google Scholar]
- Zhang, M.; Grey-Wilson, C. Bo Lu Hui Shu (in Chinese). Flora China. 2008, 7, pp. 287–288. Available online: http://flora.huh.harvard.edu/china/PDF/PDF07/Macleaya.pdf (Accessed: December,2010).
- Katz, A.W. Perennials for Midwestern Gardens: Proven Plants for the Heartland; Timber Press: Portland, Oregon, 2008. [Google Scholar]
- Kosina, P.; Gregorova, J.; Gruz, J.; Vacek, J.; Kolard, M.; Vogele, M.; Roose, W.; Naumannf, K.; Simaneka, V.; Ulrichovaa, J. Phytochemical and antimicrobial characterization of Macleaya cordata herb. Fitoterapia 2010, 81, 1006–1012. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, Y.; Guo, M.; Lou, X.; Yao, S. A simple and sensitive Metod of nonaqueouscapillary electrophoresis with laser-induced native fluorescence detection for the analysisof chelerythrine and sanguinarine in Chinese herbal medicines. Talanta 2006, 70, 202–207. [Google Scholar] [CrossRef]
- Pi, G.; Ren, P.; Yu, J.; Shi, R.; Yuan, Z.; Wang, C. Separation of sanguinarine and chelerythrine in Macleaya cordata (Willd) R. Br. based on methyl acrylate-co-divinylbenzen macroporous adsorbents. J. Chromatogr. A 2008, 1192, 17–24. [Google Scholar]
- Zhang, F.; Chen, B.; Xiao, S.; Yao, S. Optimatization and comparsion of different extraction techniques for sanguinarine and chelerythrine in fruits of Macleaya cordata (Willd) R. Br. Sep. Purif. Technol. 2005, 42, 283–290. [Google Scholar] [CrossRef]
- Faddeieva, M.D.; Beliaeva, T.N. Sanguinarine and ellipticine: cytotoxic alkaloids isolated from well-known antitumor plants. Intracellular targets of their action. Tsitologiya 1997, 39, 181–208. [Google Scholar]
- Zdarilova, A.; Vrablova, E.; Vostalova, J.; Klejdus, B.; Stejskal, D.; Proskova, J.; Kosina, P.; Svobodova, A.; Vecera, R.; Hrbac, J.; Cernochova, D.; Vicar, J.; Ulrichova, J.; Šimánek, V. Natural feed additive of Macleaya cordata: Safety assessment in rats a 90-day Frediny experiment. Food Chem. Toxicol. 2008, 46, 3721–3726. [Google Scholar]
- Zdunczyk, Z.; Gruzauskas, R.; Juskiewicz, J.; Semaskaite, A.; Jankowski, J.; Godycka, I.; Klos, J.; Jarule, V.; Miezeliene, A.; Alencikiene, G. Growth performance, gastrointestinal tractresponses, and meat characteristics of broiler chickens fed a diet containing the naturalalkaloid sanguinarine from Macleaya cordata. J. Appl. Poult. Res. 2010, 19, 393–400. [Google Scholar] [CrossRef]
- Rawling, M.D.; Merrifield, D.L.; Davies, S.J. Preliminary assessment of dietary supplementation of Sangrovit (R) on red tilapia (Oreochromis niloticus) growth performance and health. Aquaculture 2009, 294, 118–122. [Google Scholar] [CrossRef]
- Newman, S.E.; Roll, M.J.; Harkrader, R.J. A naturally occuring compound for controllingpowdery mildew of greenhouse roses. Hort Sci. 1999, 34, 686–689. [Google Scholar]
- Kosina, P.; Walterová, D.; Ulrichová, J.; Lichnovský, V.; Stiborová, M.; Rýdlová, H.; Vicar, J.; Krecman, V.; Brabec, M.J.; Simánek, V. Sanguinarine and chelerythrine: assessment of safety on pigs in ninety days feeding experiment. Food Chem. Toxicol. 2004, 42, 85–91. [Google Scholar] [CrossRef]
- Deng, A.-J.; Hai-Lin, Q. Cytotoxic dihydrobenzophenanthridine alkaloids from the roots of Macleaya microcarpa. Phytochemistry 2010, 71, 816–822. [Google Scholar] [CrossRef]
- Feng, G.; Zhang, J.; Li, X.W.; Feng, J.T.; Zhang, X. Insecticidal activity of alkaloids from Macleaya microcarpa against several species of insect pests. Agric. Life Sci. 2008, 34 (in Chinese with English abstract)., 187–192. [Google Scholar]
- Wang, G.-X.; Zhou, Z.; Jiang, D.-X.; Han, J.; Wang, J.-F. In vivo anthelmintic activity of five alkaloids from Macleaya microcarpa. Vet. Parasitol. 2010, 171, 305–313. [Google Scholar] [CrossRef]
- Dostál, J.; Slavík, J. Novější poznatky o sanguinarinu a přbuzných alkaloidech. Chem. Listy 2000, 94, 15–20. [Google Scholar]
- Suchomelová, J.; Bochořáková, H.; Paulová, H.; Musil, P.; Táborská, E. HPLC quantification of seven quaternary benzo[c]phenanthridine alkaloids in six species of the family Papaveraceae. J. Pharm. Biomed. Anal. 2007, 44, 283–287. [Google Scholar] [CrossRef]
- Slaninová, I.; Slanina, J.; Táborská, E. Quaternary benzo[c]phenanthridine alkaloids-novel cell permeant and red fluorescing DNA probes. Cytom. Part A 2007, 71A, 700–708. [Google Scholar] [CrossRef]
- Zdarilova, A.; Malikova, J.; Dvorak, Z.; Ulrichova, J.; Šimánek, V. Quaternary isoquinoline alkaloids sanguinarine and chelerythrine. In vitro and in vivo effects. Chem. Listy 2006, 100, 30–41. [Google Scholar]
- Colombo, M.L.; Bosisio, E. Pharmacological activities of Chelidonium majus L. (Papaveraceae). Pharmacol. Res. 1996, 33, 127–134. [Google Scholar]
- Ahmad, N.; Gupta, S.; Husain, M.M.; Heiskanen, K.M.; Mukhtar, H. Differential Antiproliferative and Apoptotic Response of Sanguinarine for Cancer Cells versus Normal Cells. Clin. Cancer Res. 2000, 6, 1524–1528. [Google Scholar]
- Barreto, M.C.; Pinto, R.E.; Arrabaça, J.D.; Pavlo, M.L. Inhibition of mouse liver respiration by Chelidonium majus isoquinoline alkaloids. Toxicol. Lett. 2003, 146, 37–47. [Google Scholar] [CrossRef]
- Lopus, M.; Panda, D. The benzophenanthridine alkaloid sanguinarine perturbsmicrotubule assembly dynamics through tubulin binding. FEBS J. 2006, 273, 2139–2150. [Google Scholar] [CrossRef]
- Kaminskyy, V.O.; Lootsik, M.D.; Stoika, R.S. Correlation of the cytotoxic activity of four different alkaloids, from Chelidonium majus (greater celandine), with their DNA intercalating properties and ability to induce breaks in the DNA of NK/Ly murine lymphoma cells. Centr. Eur. J. Biol. 2006, 1, 2–15. [Google Scholar] [CrossRef]
- Adhami, V.M.; Aziz, M.H.; Mukhtar, H.; Ahmad, N. Activation of prodeath Bcl-2 family proteins and mitochondrial apoptosis pathway by sanguinarine in immortalized human HaCaT keratinocytes. Clin. Cancer Res. 2003, 9, 3176–3182. [Google Scholar]
- Ishikawa, T. Benzo[c]phenanthridine bases and their antituberculosis activity. Med. Res. Rev. 2001, 21, 61–72. [Google Scholar] [CrossRef]
- Slaninová, I.; Slunská, Z.; Sinkora, J.; Vlková, M.; Táborská, E. Screening of minorbenzo(c)phenanthridine alkaloids for antiproliferative and apoptotic activities. Cell Biol. Toxicol. 2007, 45, 131–139. [Google Scholar]
- Slaninová, I., Táborská; Bochořáková, H.; Slanina, J. Interaction of benzo[c]phenanthridine and protoberberine alkaloids with animal and yeast cells. Cell Biol. Toxicol. 2001, 17, 51–63. [Google Scholar] [CrossRef]
- Urbanová, J.; Lubal, P.; Slaninová, I.; Táborská, E.; Táborský, P. Fluorescence proparties ofselected benzo[c]phenanthridine alkaloids and studies of their interaction with CT DNA. Anal. Bioanal. Chem. 2009, 394, 997–1002. [Google Scholar] [CrossRef]
- Slunská, Z.; Gelnarová, E.; Hammerová, J.; Táborská, E.; Slaninová, I. Effect of quaternarybenzophenanthridine alkaloids sanguilutine and chelilutine on normal and cancer cells. Toxicol. In Vitro 2010, 24, 697–706. [Google Scholar] [CrossRef]
- Zenk, M.H. The formation of benzophenanthridine alkaloids. Pure Appl. Chem. 1994, 66, 2023–2028. [Google Scholar] [CrossRef]
- Abizov, E.A.; Tolkachev, O.N.; Kopylova, I.E.; Luferov, A.N. Distribution of the sum of sanguinarine and chelerythrine in the above-ground part of Macleaya microcarpa. Pharm. Chem. J. 2003, 37, 413–414. [Google Scholar] [CrossRef]
- Dostál, J.; Slavik, J. Some aspects of the chemistry of quaternary benzo[c]phenanthridine alkaloids. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier Science B.V.: Oxford, UK, 2002; Volume 27, pp. 155–184. [Google Scholar]
- Gregorova, J.; Babica, J.; Marek, R.; Paulova, H.; Táborská, E.; Dostal, J. Extraction ofisoquinoline alkaloids with butanol and octanol. Fitoterapia 2010, 81, 565–568. [Google Scholar] [CrossRef]
- Sample Availability: Not Available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pěnčíková, K.; Urbanová, J.; Musil, P.; Táborská, E.; Gregorová, J. Seasonal Variation of Bioactive Alkaloid Contents in Macleaya microcarpa (Maxim.) Fedde. Molecules 2011, 16, 3391-3401. https://doi.org/10.3390/molecules16043391
Pěnčíková K, Urbanová J, Musil P, Táborská E, Gregorová J. Seasonal Variation of Bioactive Alkaloid Contents in Macleaya microcarpa (Maxim.) Fedde. Molecules. 2011; 16(4):3391-3401. https://doi.org/10.3390/molecules16043391
Chicago/Turabian StylePěnčíková, Kristýna, Jana Urbanová, Pavel Musil, Eva Táborská, and Jana Gregorová. 2011. "Seasonal Variation of Bioactive Alkaloid Contents in Macleaya microcarpa (Maxim.) Fedde" Molecules 16, no. 4: 3391-3401. https://doi.org/10.3390/molecules16043391
APA StylePěnčíková, K., Urbanová, J., Musil, P., Táborská, E., & Gregorová, J. (2011). Seasonal Variation of Bioactive Alkaloid Contents in Macleaya microcarpa (Maxim.) Fedde. Molecules, 16(4), 3391-3401. https://doi.org/10.3390/molecules16043391