Fungitoxicity against Botrytis cinerea of a Flavonoid Isolated from Pseudognaphalium robustum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
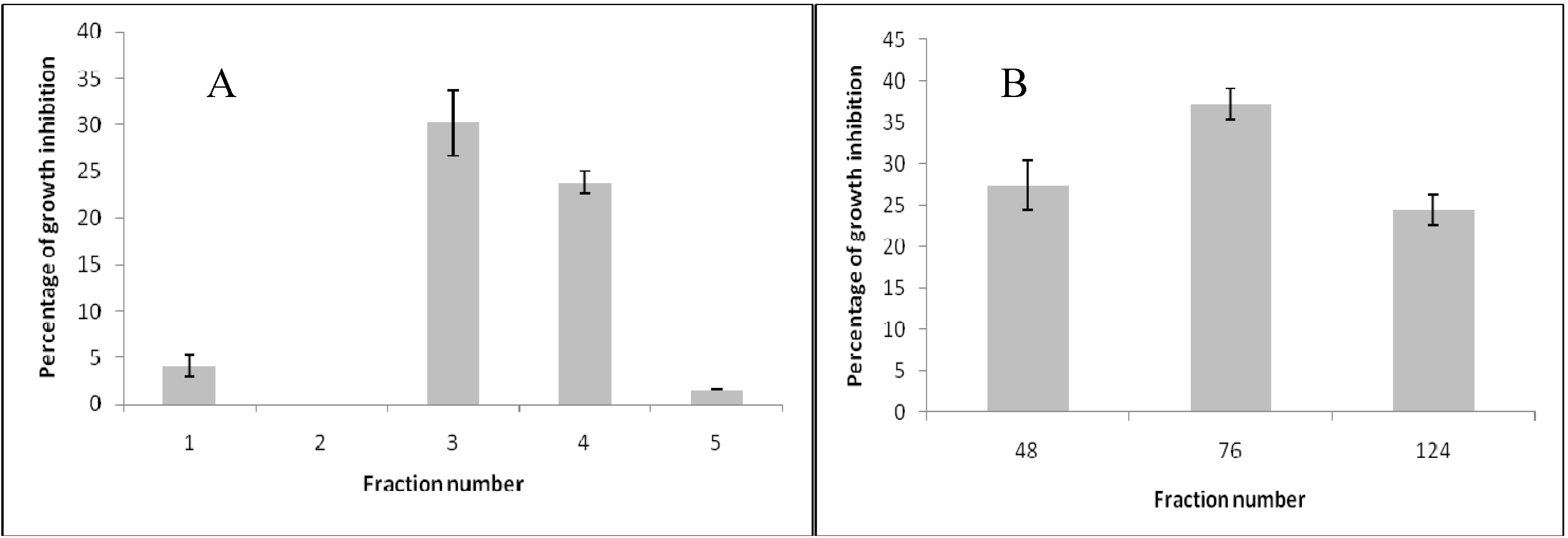
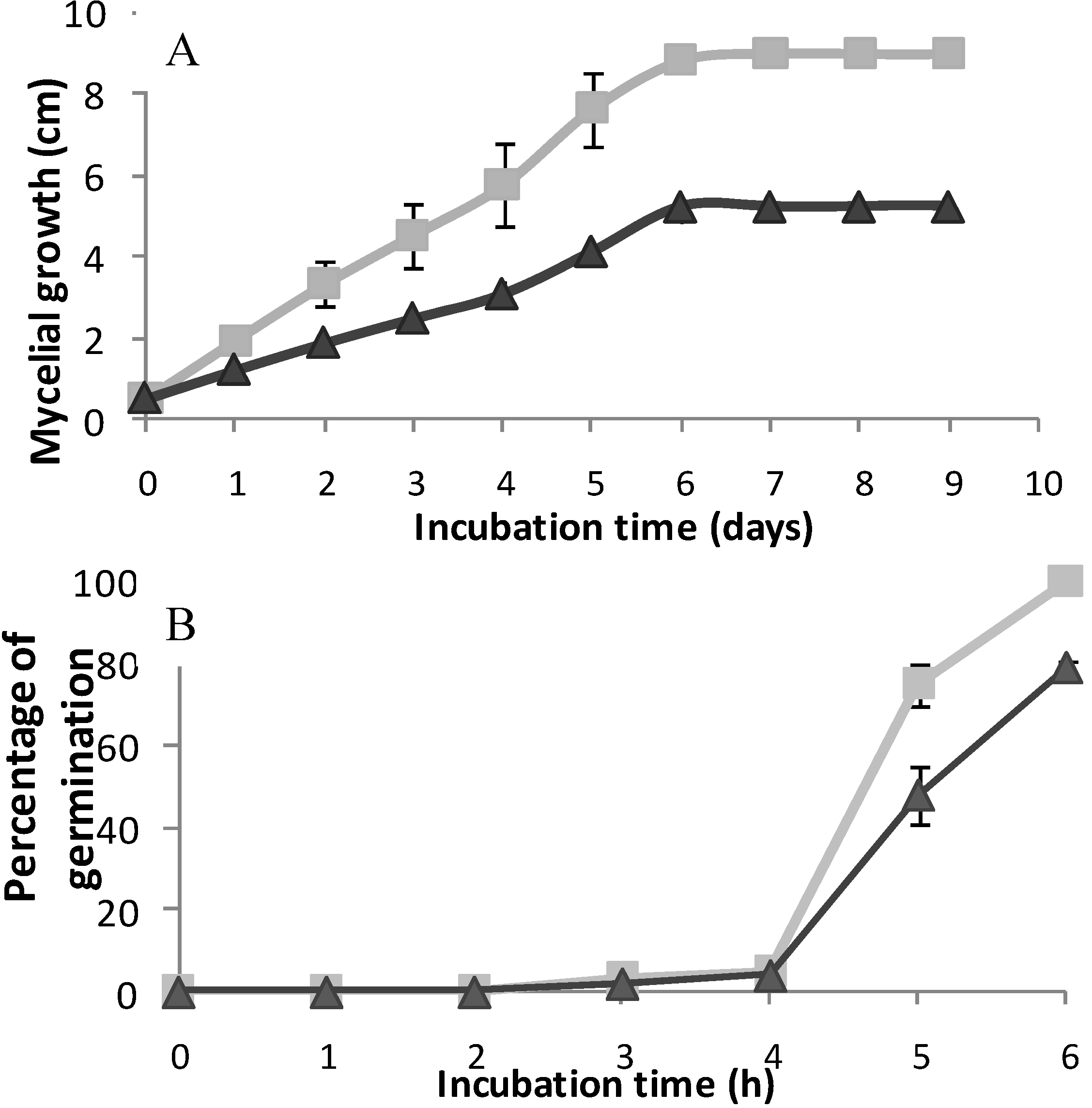
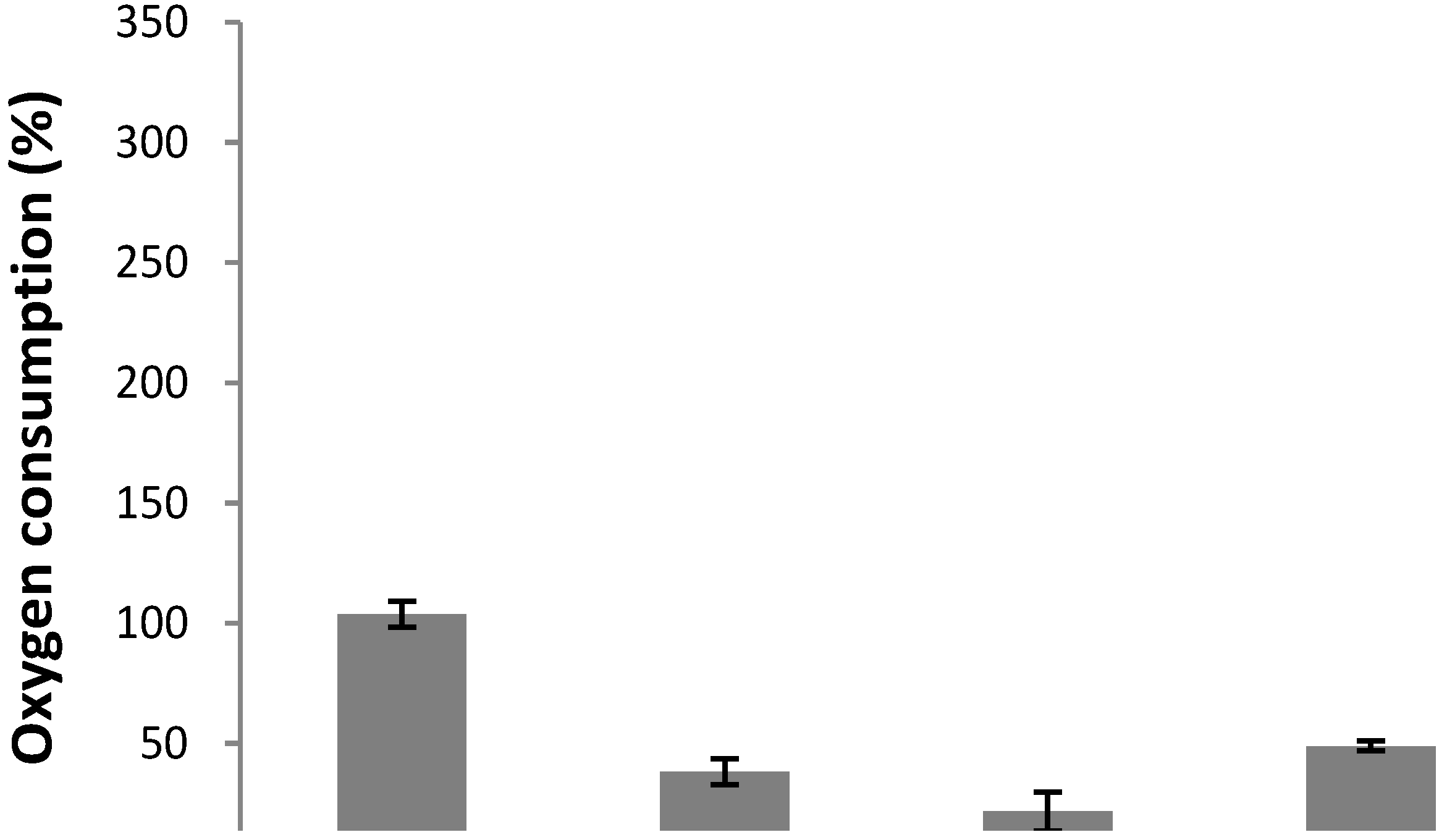
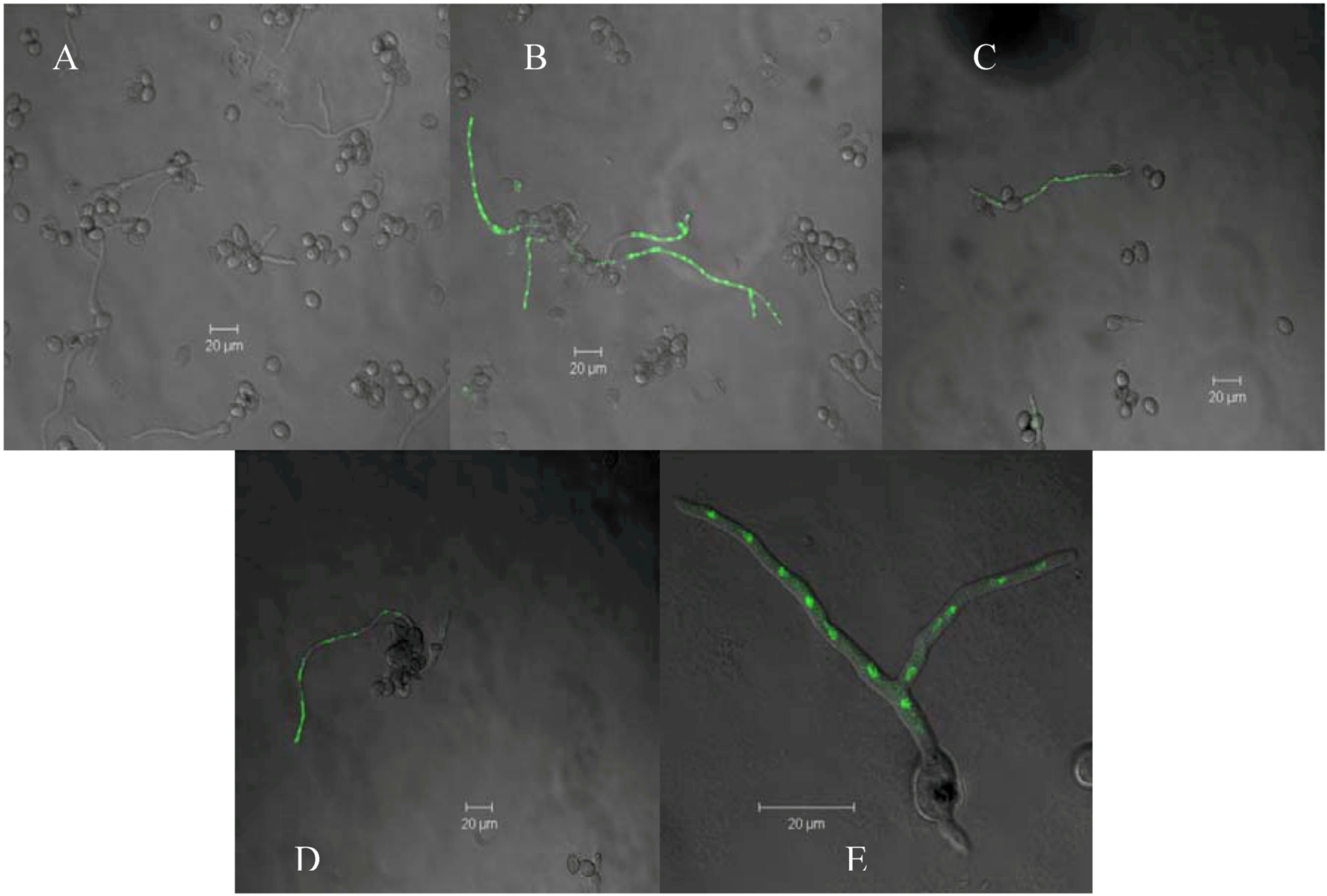
2. Results and Discussion


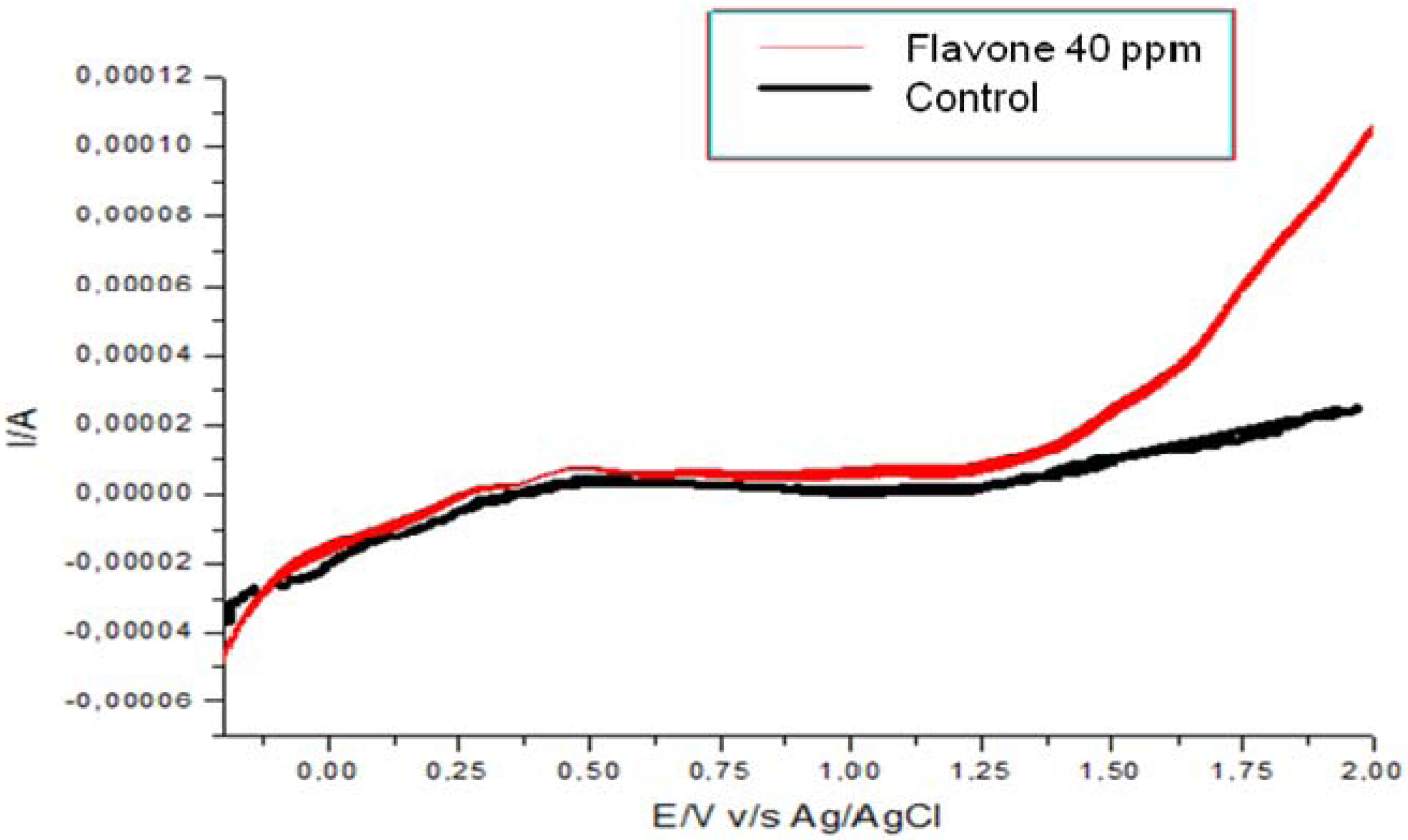
 ) and treatment assays. Each point represents the mean of at least three independent experiments ± standard deviation.
) and treatment assays. Each point represents the mean of at least three independent experiments ± standard deviation.
) and treatment assays. Each point represents the mean of at least three independent experiments ± standard deviation.
) and treatment assays. Each point represents the mean of at least three independent experiments ± standard deviation.



3. Experimental
3.1. Bioassay-guided Fractionation of Resinous Exudate from Pseudognaphalium Robustum
3.2. Fungal Isolate and Culture Conditions
3.3. Effect of Column Fractions and Pure Compound on B. cinerea
3.4. Effect of Pure Compound on the Membrane Integrity of B. cinerea
3.5. Effect of Pure Compound on the Oxygen Consumption of B. cinerea Conidia
3.6. Pro-oxidant Activity
4. Conclusions
Acknowledgements
References
- Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N. Botrytis spp. and Diseases They Cause in Agricultural Systems – an. In troduction. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 1–6. [Google Scholar]
- Leroux, P. Chemical Control of Botrytis cinerea and Its Resistance to Chemical Fungicides. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 195–222. [Google Scholar]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Simic, A.; Manojlovic, D.; Segan, D.; Todorovic, M. Electrochemical behaviour and antioxidant and prooxidant activity of natural phenolics. Molecules 2007, 12, 2327–2340. [Google Scholar] [CrossRef]
- Padmavati, M.; Sakthivel, N.; Thara, K.V.; Reddy, A.R. Differential sensitivity of rice pathogens to growth inhibition by flavonoids. Phytochemistry 1997, 46, 499–502. [Google Scholar]
- Grayer, R.; Harbone, J. A survey of antifungal compounds from higher plants. Phytochemistry 1994, 37, 19–42. [Google Scholar]
- Langcake, P.; McCarthy, W. The relationship of resveratrol production to infection of grapevine leaves by Botrytis cinerea. Vitis 1979, 18, 244–253. [Google Scholar]
- Urzúa, A.; Cuadra, P. Flavonoids from the resinous exudate of Gnaphalium robustum. Bol. Soc. Chil. Quim. 1989, 34, 247–251. [Google Scholar]
- Urzúa, A.; Cuadra, P. Acylated flavonoids aglycones from Gnaphalium robustum. Phytochemistry 1990, 29, 1342–1345. [Google Scholar]
- Cotoras, M.; García, C.; Lagos, C.; Folch, C.; Mendoza, L. Antifungal activity on Botrytis cinerea of flavonoids and diterpenoids isolated from the surface of Pseudognaphalium spp. Bol. Soc. Chil. Quim. 2001, 46, 433–440. [Google Scholar]
- Urzúa, A.; Torres, R.; Bueno, C.; Mendoza, L. Flavonoids and diterpenoids in the trichome resinous exudates from Pseudognaphalium cheiranthifolium, P heterotrichium and P. vira vira. Biochem. Syst. Ecol. 1995, 23, 459. [Google Scholar] [CrossRef]
- Mendoza, L.; Urzúa, A. Minor flavonoids and diterpenoids in the trichome resinous exudates from Pseudognaphalium cheiranthifolium, P heterotrichium, P. vira vira and P. robustum. Biochem. Syst. Ecol. 1998, 26, 469–471. [Google Scholar] [CrossRef]
- Schinella, G.R.; Tournier, H.A.; Máñez, S.; de Buschiazzo, P.M.; Recio, M.C.; Ríos, J.L. Tiliroside and gnaphaliin inhibit human low density lipoprotein oxidation. Fitoterapia 2007, 78, 1–6. [Google Scholar] [CrossRef]
- Sala, A.; Recio, M.C.; Schinella, G.R.; Máñez, S.; Giner, R.M.; Cerdá-Nicolás, M.; Rosí, J.L. Assessment of the anti-inflammatory activity and free radical scavenger activity of tiliroside. Eur. J. Pharmacol. 2003, 461, 53–61. [Google Scholar] [CrossRef]
- Aleu, J.; Collado, I.G. Biotransformations by Botrytis species. J. Mol. Catal. B Enzym. 2001, 123, 77–93. [Google Scholar]
- Schouten, A.; Wagemakers, L.; Stefanato, F.L.; van der Kaaij, R.M.; van Kan, J.A.L. Resveratrol acts as a natural profungicide and induces self-intoxication by a specific laccase. Mol. Microbiol. 2002, 43, 883–894. [Google Scholar] [CrossRef]
- Slawecki, R.; Ryan, E.; Young, D. Novel fungitoxic assay for inhibition of germination-associated adhesion of Botrytis cinerea and Puccinia recondite spores. Appl. Environ. Microbiol. 2002, 68, 597–601. [Google Scholar] [CrossRef]
- Dorta, D.J.; Pigoso, A.A.; Mingatto, F.E.; Rodrigues, T.; Prado, I.M.R.; Helena, A.F.C.; Uyemura, S.A.; Santos, A.C.; Curti, C. The interaction of flavonoids with mitochondria: Effects on energetic processes. Chem. Biol. Interact. 2005, 152, 67–78. [Google Scholar] [CrossRef]
- Laks, P.; Pruner, M. Flavonoid biocides; structure/activity relations of flavonoid phytoalexin analogues. Phytochemistry 1989, 28, 87–91. [Google Scholar]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar]
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part I: Chemical diversity, impacts on plant biology and human health. Biotechnol. J. 2007, 2, 1214–1234. [Google Scholar]
- Plochmann, K.; Korte, G.; Koutsiliere, E.; Richling, E.; Riederer, P.; Rethwilm, A.; Schreier, P.; Scheller, C. Structure-activity relationships of flavonoid-induced cytotoxicity on human leukemia cells. Arch. Biochem. Biophys. 2007, 460, 1–9. [Google Scholar] [CrossRef]
- Hodnick, W.F.; Milosavljevic, E.B.; Nelson, J.H.; Pardini, R.S. Electrochemistry of flavonoids. Relationships between redox potentials, inhibition of mitochondrial respiration and production of oxygen radicals by flavonoids. Biochem. Pharmacol. 1988, 37, 2607–2611. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Mochizuki, M.; Yamazaki, S.; Kano, K.; Ikeda, T. Kinetic analysis and mechanistic aspects of autoxidation of catehins. Biochim. Biophys. Acta 2002, 1569, 35–44. [Google Scholar] [CrossRef]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. Flavonoids as antioxidants: Determination of radical scavenging efficiencies. Methods Enzymol. 1990, 186, 343–355. [Google Scholar]
- Muñoz, G.; Hinrichsen, P.; Brygoo, Y.; Giraud, T. Genetic characterisation of Botrytis cinerea populations in Chile. Mycol. Res. 2002, 106, 594–601. [Google Scholar] [CrossRef]
- Mendoza, L.; Espinoza, P.; Urzúa, A.; Vivanco, M.; Cotoras, M. In vitro antifungal activity of the diterpenoid 7α-Hidroxy-8(17)-labden-15-oic acid and its derivates against Botrytis cinerea. Molecules 2009, 14, 1966–1979. [Google Scholar] [CrossRef]
- Thevissen, K.; Terras, F.R.G.; Broekaert, W.F. Permeabilization of fungal membranes by plant defensins inhibit fungal growth. Appl. Environ. Microbiol. 1999, 65, 5451–5458. [Google Scholar]
- Sample Availability: Samples of the compound 5,7-dihydroxy-3,8-dimethoxyflavone are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cotoras, M.; Mendoza, L.; Muñoz, A.; Yáñez, K.; Castro, P.; Aguirre, M. Fungitoxicity against Botrytis cinerea of a Flavonoid Isolated from Pseudognaphalium robustum. Molecules 2011, 16, 3885-3895. https://doi.org/10.3390/molecules16053885
Cotoras M, Mendoza L, Muñoz A, Yáñez K, Castro P, Aguirre M. Fungitoxicity against Botrytis cinerea of a Flavonoid Isolated from Pseudognaphalium robustum. Molecules. 2011; 16(5):3885-3895. https://doi.org/10.3390/molecules16053885
Chicago/Turabian StyleCotoras, Milena, Leonora Mendoza, Alexis Muñoz, Karen Yáñez, Paulo Castro, and María Aguirre. 2011. "Fungitoxicity against Botrytis cinerea of a Flavonoid Isolated from Pseudognaphalium robustum" Molecules 16, no. 5: 3885-3895. https://doi.org/10.3390/molecules16053885
APA StyleCotoras, M., Mendoza, L., Muñoz, A., Yáñez, K., Castro, P., & Aguirre, M. (2011). Fungitoxicity against Botrytis cinerea of a Flavonoid Isolated from Pseudognaphalium robustum. Molecules, 16(5), 3885-3895. https://doi.org/10.3390/molecules16053885