Cytotoxicity and Pro-Apoptotic Activity of 2,2´-Bis[4,5-bis(4-hydroxybenzyl)-2-(4-hydroxyphenyl)cyclopent-4-en-1,3-dione], a Phenolic Cyclopentenedione Isolated from the Cyanobacterium Strain Nostoc sp. str. Lukešová 27/97

Abstract
:1. Introduction

2. Results and Discussion



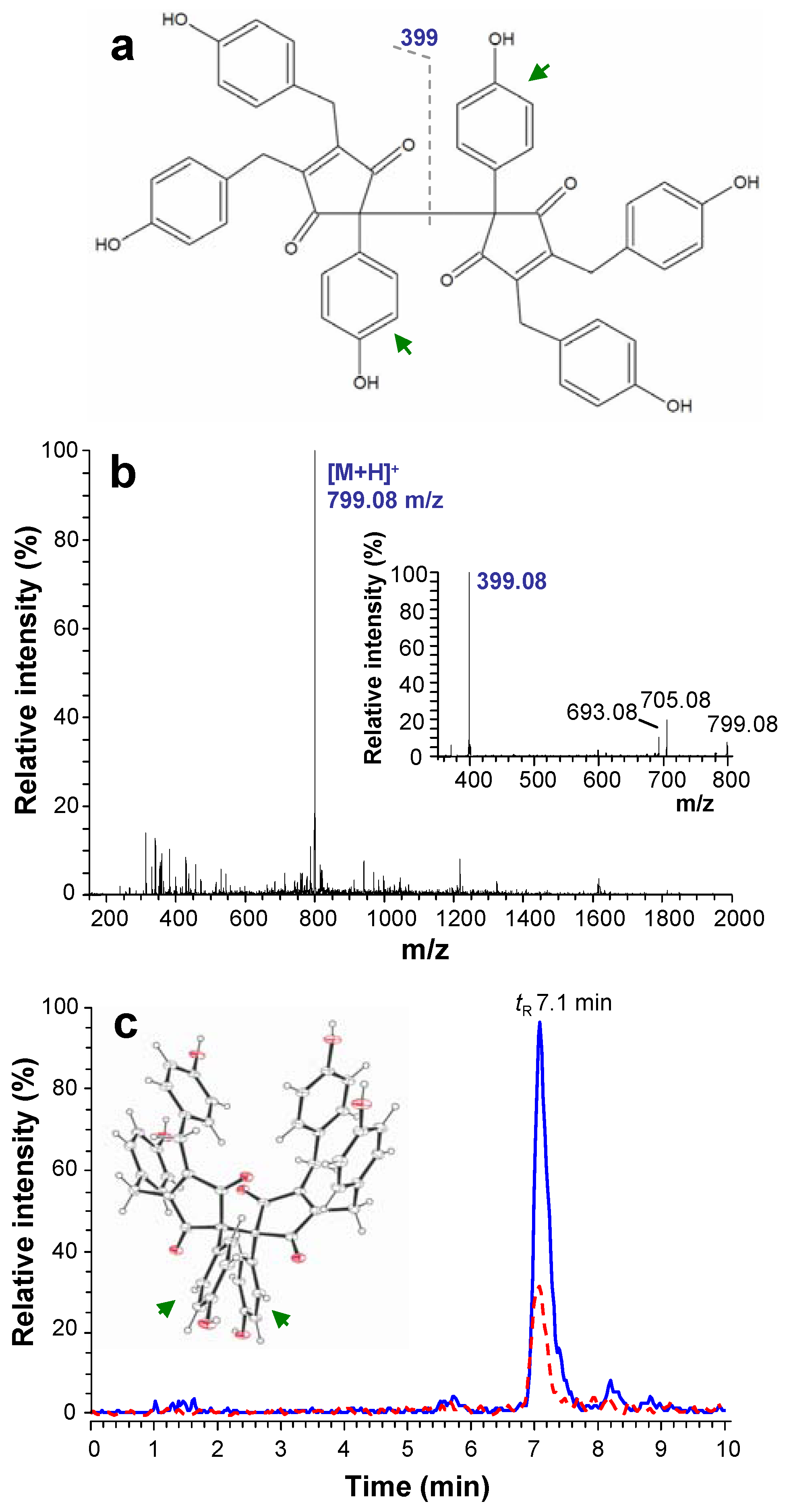{kind=link}
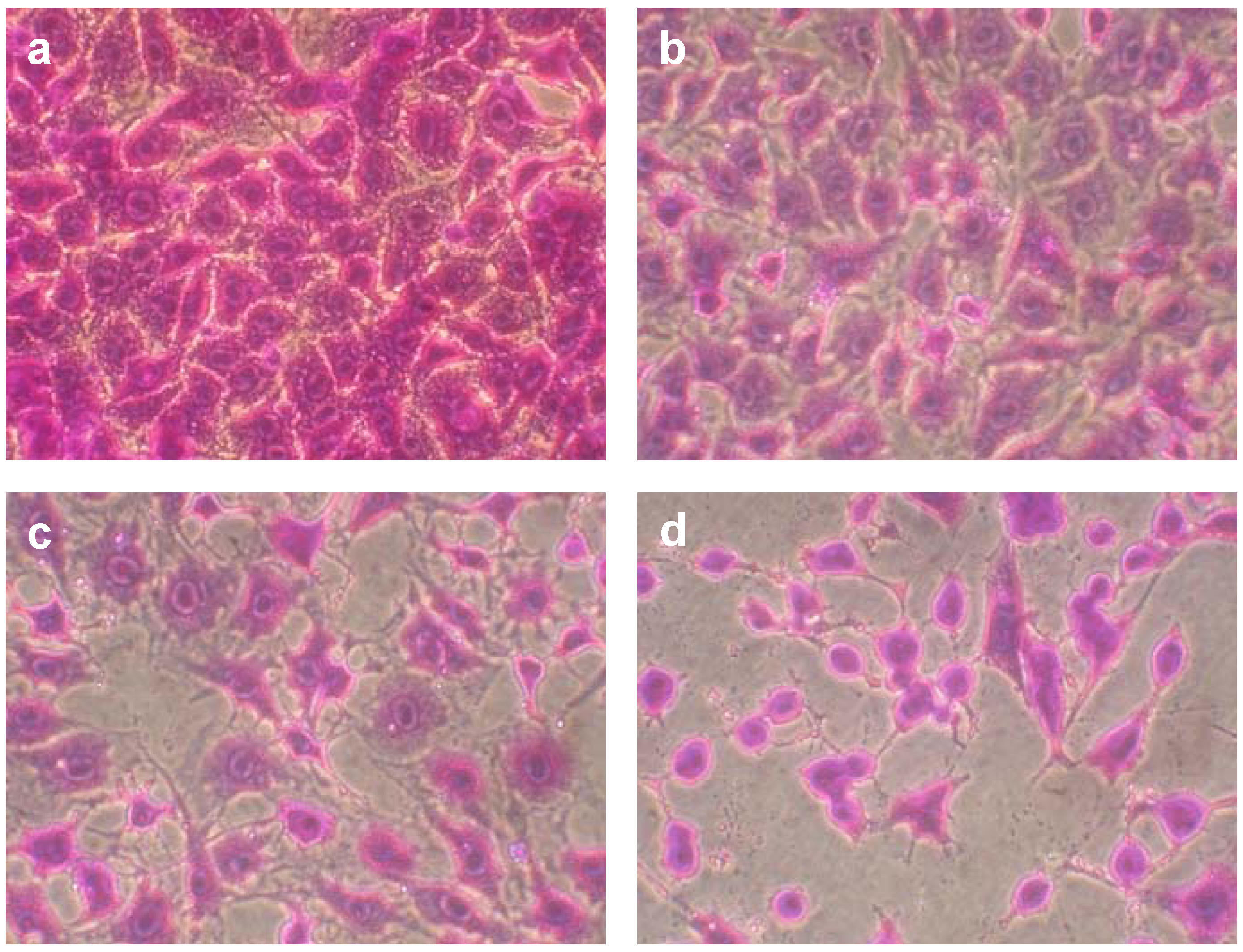{kind=link}
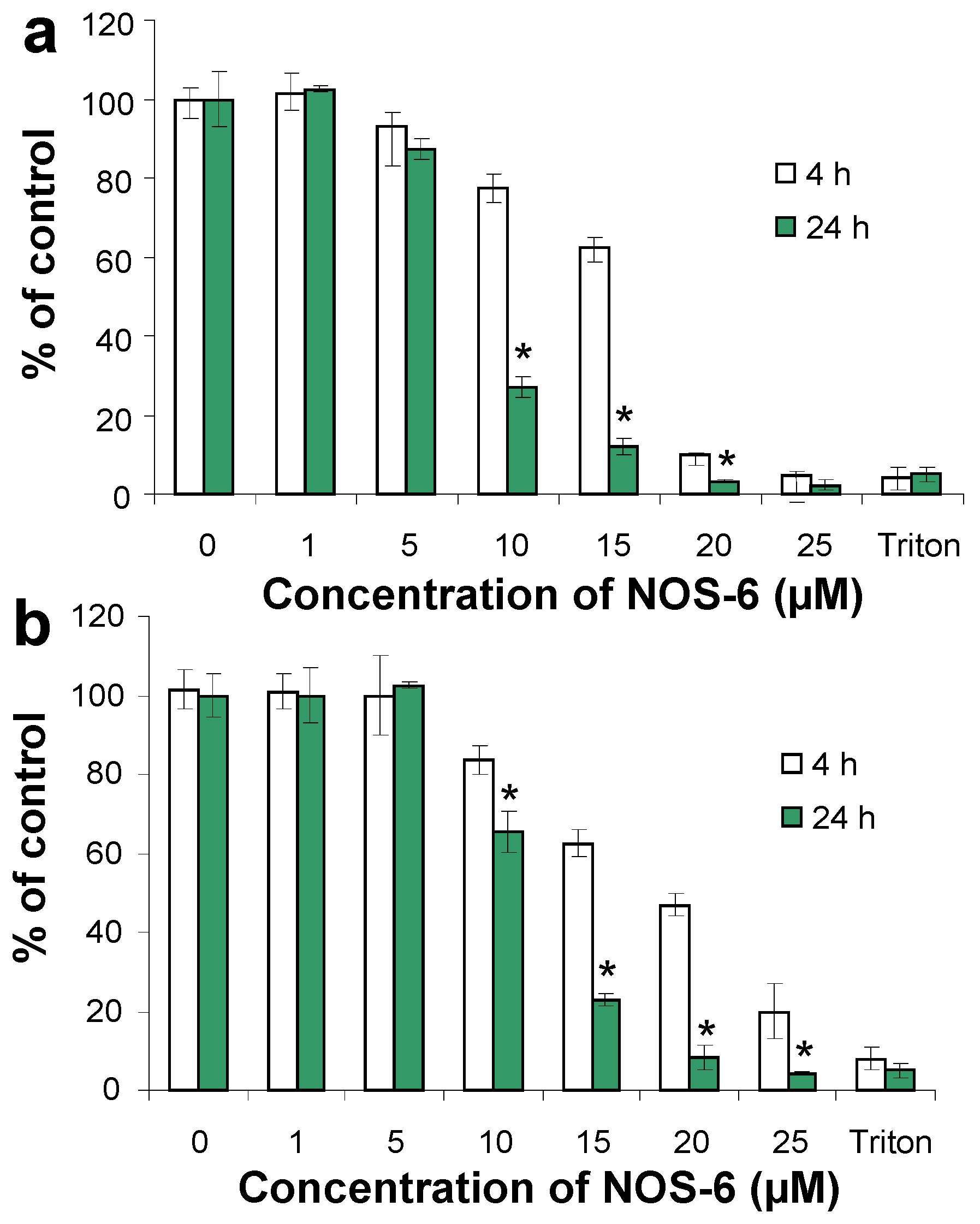{kind=link}
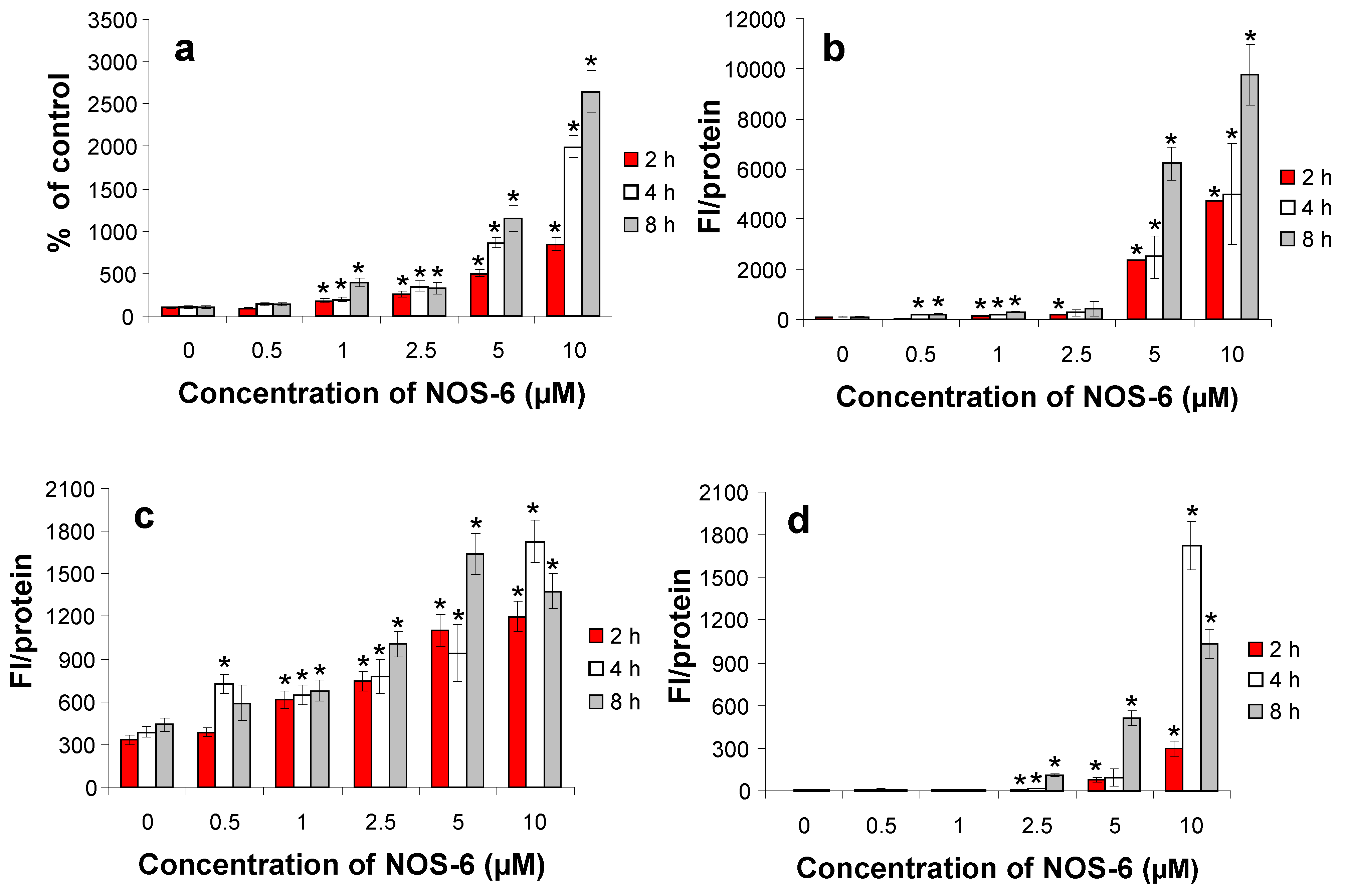{kind=link}
| Compound | IC50 (µM) | |
|---|---|---|
| NR assay | MTT assay | |
| NOS-6 | 8.48 ± 0.16 | 12.15 ± 1.96 |
| Doxorubicin | 1.26 ± 0.42 | 2.04 ± 0.48 |

3. Experimental
3.1. Chemicals
3.2. Isolation of NOS-6
3.3. Cell culture
3.4. Identification of NOS-6 and its uptake into mouse fibroblasts
3.5. Cytotoxicity of NOS-6
3.6. Statistical analysis
4. Conclusions
Acknowledgements
References
- Plaza, M.; Santoyo, S.; Jaime, L.; Reina, G.G.B.; Herrero, M.; Senorans, F.J.; Ibanez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar]
- Klejdus, B.; Kopecky, J.; Benesova, L.; Vacek, J. Solid-phase/supercritical-fluid extraction for liquid chromatography of phenolic compounds in freshwater microalgae and selected cyanobacterial species. J. Chromatogr. A 2009, 1216, 763–771. [Google Scholar]
- Onofrejova, L.; Vasickova, J.; Klejdus, B.; Stratil, P.; Misurcova, L.; Kracmar, S.; Kopecky, J.; Vacek, J. Bioactive phenols in algae: The application of pressurized-liquid and solid-phase extraction techniques. J. Pharm. Biomed. Anal. 2010, 51, 464–470. [Google Scholar] [CrossRef]
- Ragan, M.A.; Glombitza, K.W. Phlorotannins, brown algal polyphenols. In Progress in Phycological Research; Round, F.E., Chapman, D.J., Eds.; Biopress: Bristol, UK, 1986; pp. 129–241. [Google Scholar]
- La Barre, S.; Potin, P.; Leblanc, C.; Delage, L. The halogenated metabolism of brown algae (phaeophyta), its biological importance and its environmental significance. Mar. Drugs 2010, 8, 988–1010. [Google Scholar] [CrossRef]
- Vacek, J.; Snoblova, M.; Klejdus, B. A short introduction to algal and cyanobacterial constituents - the occurrence of phenolic metabolites (in czech). Čes. Slov. Farm. 2009, 58, 103–108. [Google Scholar]
- Koivikko, R.; Loponen, J.; Honkanen, T.; Jormalainen, V. Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga fucus vesiculosus, with implications on their ecological functions. J. Chem. Ecol. 2005, 31, 195–212. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2010, 27, 165–237. [Google Scholar] [CrossRef]
- Zelik, P.; Kopecky, J.; Cegan, A. Acetylcholinesterase inhibitors from microalgae (in czech). Chem. Listy 2005, 99, s356–s357. [Google Scholar]
- Zelik, P.; Lukesova, A.; Cejka, J.; Budesinsky, M.; Havlicek, V.; Cegan, A.; Kopecky, J. Nostotrebin 6, a bis(cyclopentenedione) with cholinesterase inhibitory activity isolated from Nostoc sp str. Lukesova 27/97. J. Enzym. Inhib. Med. Chem. 2010, 25, 414–420. [Google Scholar] [CrossRef]
- Zelik, P.; Lukesova, A.; Voloshko, L.; Stys, D.; Kopecky, J. Screening for acetylcholinesterase inhibitory activity in cyanobacteria of the genus Nostoc. J. Enzym. Inhib. Med. Chem. 2009, 24, 531–536. [Google Scholar]
- Babu, K.S.; Li, X.C.; Jacob, M.R.; Zhang, Q.F.; Khan, S.I.; Ferreira, D.; Clark, A.M. Synthesis, antifungal activity, and structure-activity relationships of coruscanone a analogues. J. Med. Chem. 2006, 49, 7877–7886. [Google Scholar] [CrossRef]
- Gilardoni, G.; Clericuzio, M.; Tosi, S.; Zanoni, G.; Vidari, G. Antifungal acylcyclopentenediones from fruiting bodies of hygrophorus chrysodon. J. Nat. Prod. 2007, 70, 137–139. [Google Scholar] [CrossRef]
- Li, X.C.; Ferreira, D.; Jacob, M.R.; Zhang, Q.F.; Khan, S.I.; ElSohly, H.N.; Nagle, D.G.; Smillie, T.J.; Khan, I.A.; Walker, L.A.; Clark, A.M. Antifungal cyclopentenediones from piper coruscans. J. Am. Chem. Soc. 2004, 126, 6872–6873. [Google Scholar]
- Shestak, O.P.; Novikov, V.L.; Martyyas, E.A.; Anisimov, M.M. Synthesis and antimicrobial and antifungal activities of cyclopentene beta,beta'-triketones and their methyl enol esters. Pharm. Chem. J. 2009, 43, 498–501. [Google Scholar] [CrossRef]
- Sturino, C.F.; Doussot, P.; Paquette, L.A. Antimicrobial algal metabolites of unusual structure. Concise synthesis of the highly oxygenated [4.4]spirononene dimethyl gloiosiphone a by ring expansion of dimethyl squarate. Tetrahedron 1997, 53, 8913–8926. [Google Scholar]
- Wijeratne, E.M.K.; Turbyville, T.J.; Zhang, Z.G.; Bigelow, D.; Pierson, L.S.; VanEtten, H.D.; Whitesell, L.; Canfield, L.M.; Gunatilaka, A.A.L. Cytotoxic constituents of aspergillus terreus from the rhizosphere of opuntia versicolor of the sonoran desert. J. Nat. Prod. 2003, 66, 1567–1573. [Google Scholar] [CrossRef]
- Noble, M.; Noble, D.; Fletton, R.A. G2201-c, a new cyclopentenedione antibiotics, isolated from fermentation broth of streptomyces-cattleya. J. Antibiotics 1978, 31, 15–18. [Google Scholar] [CrossRef]
- Hori, H.; Nagasawa, H.; Ishibashi, M.; Uto, Y.; Hirata, A.; Saijo, K.; Ohkura, K.; Kirk, K.L.; Uehara, Y. Tx-1123: An antitumor 2-hydroxyarylidene-4-cyclopentene-1,3-dione as a protein tyrosine kinase inhibitor having low mitochondrial toxicity. Bioorg. Med. Chem. 2002, 10, 3257–3265. [Google Scholar] [CrossRef]
- Hori, H.; Nagasawa, H.; Uto, Y. Structure-based design of the antitumor 2-hydroxyarylidene-4-cyclopentene-1,3-dione tx-1123 as a protein tyrosine kinase inhibitor having low mitochondrial toxicity. Cell. Mol. Biol. Lett. 2003, 8, 528–530. [Google Scholar]
- Kong, A.N.T.; Yu, R.; Chen, C.; Mandlekar, S.; Primiano, T. Signal transduction events elicited by natural products: Role of mapk and caspase pathways in homeostatic response and induction of apoptosis. Arch. Pharm. Res. 2000, 23, 1–16. [Google Scholar] [CrossRef]
- Oh, H.M.; Choi, S.K.; Lee, J.M.; Lee, S.K.; Kim, H.Y.; Han, D.C.; Kim, H.M.; Son, K.H.; Kwon, B.M. Cyclopentenediones, inhibitors of farnesyl protein transferase and anti-tumor compounds, isolated from the fruit of lindera erythrocarpa makino. Bioorg. Med. Chem. 2005, 13, 6182–6187. [Google Scholar] [CrossRef]
- Capkova, K.; Hixon, M.S.; Pellett, S.; Barbieri, J.T.; Johnson, E.A.; Janda, K.D. Benzylidene cyclopentenediones: First irreversible inhibitors against botulinum neurotoxin a's zinc endopeptidase. Bioorg. Med. Chem. Lett. 2010, 20, 206–208. [Google Scholar] [CrossRef]
- Allen, M.B.; Arnon, D.I. Studies on nitrogen-fixing blue green algae: I. Growth and nitrogen fixation by anabaena cylindrica lemm. Plant Physiol. 1955, 30, 366–372. [Google Scholar]
- Maines, M.D. In Vitro Methods for Detecting Cytotoxicity in Current Protocols in Toxicology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1998. [Google Scholar]
- Svobodova, A.; Zdarilova, A.; Vostalova, J. Lonicera caerulea and vaccinium myrtillus fruit polyphenols protect HaCaT keratinocytes against UVB-induced phototoxic stress and DNA damage. J. Dermatol. Sci. 2009, 56, 196–204. [Google Scholar] [CrossRef]
- Antkowiak, R.; Antkowiak, W.Z.; Banczyk, I.; Mikolajczyk, L. A new phenolic metabolite, involutone, isolated from the mushroom paxillus involutus. Can. J. Chem. 2003, 81, 118–124. [Google Scholar] [CrossRef]
- Mikolajczyk, L.; Antkowiak, W.Z. Structure studies of the metabolites of paxillus involutus. Heterocycles 2009, 79, 423–426. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vacek, J.; Hrbáč, J.; Kopecký, J.; Vostálová, J. Cytotoxicity and Pro-Apoptotic Activity of 2,2´-Bis[4,5-bis(4-hydroxybenzyl)-2-(4-hydroxyphenyl)cyclopent-4-en-1,3-dione], a Phenolic Cyclopentenedione Isolated from the Cyanobacterium Strain Nostoc sp. str. Lukešová 27/97. Molecules 2011, 16, 4254-4263. https://doi.org/10.3390/molecules16054254
Vacek J, Hrbáč J, Kopecký J, Vostálová J. Cytotoxicity and Pro-Apoptotic Activity of 2,2´-Bis[4,5-bis(4-hydroxybenzyl)-2-(4-hydroxyphenyl)cyclopent-4-en-1,3-dione], a Phenolic Cyclopentenedione Isolated from the Cyanobacterium Strain Nostoc sp. str. Lukešová 27/97. Molecules. 2011; 16(5):4254-4263. https://doi.org/10.3390/molecules16054254
Chicago/Turabian StyleVacek, Jan, Jan Hrbáč, Jiří Kopecký, and Jitka Vostálová. 2011. "Cytotoxicity and Pro-Apoptotic Activity of 2,2´-Bis[4,5-bis(4-hydroxybenzyl)-2-(4-hydroxyphenyl)cyclopent-4-en-1,3-dione], a Phenolic Cyclopentenedione Isolated from the Cyanobacterium Strain Nostoc sp. str. Lukešová 27/97" Molecules 16, no. 5: 4254-4263. https://doi.org/10.3390/molecules16054254
APA StyleVacek, J., Hrbáč, J., Kopecký, J., & Vostálová, J. (2011). Cytotoxicity and Pro-Apoptotic Activity of 2,2´-Bis[4,5-bis(4-hydroxybenzyl)-2-(4-hydroxyphenyl)cyclopent-4-en-1,3-dione], a Phenolic Cyclopentenedione Isolated from the Cyanobacterium Strain Nostoc sp. str. Lukešová 27/97. Molecules, 16(5), 4254-4263. https://doi.org/10.3390/molecules16054254