Fluorescent Probes Detecting the Phagocytic Phase of Apoptosis: Enzyme-Substrate Complexes of Topoisomerase and DNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
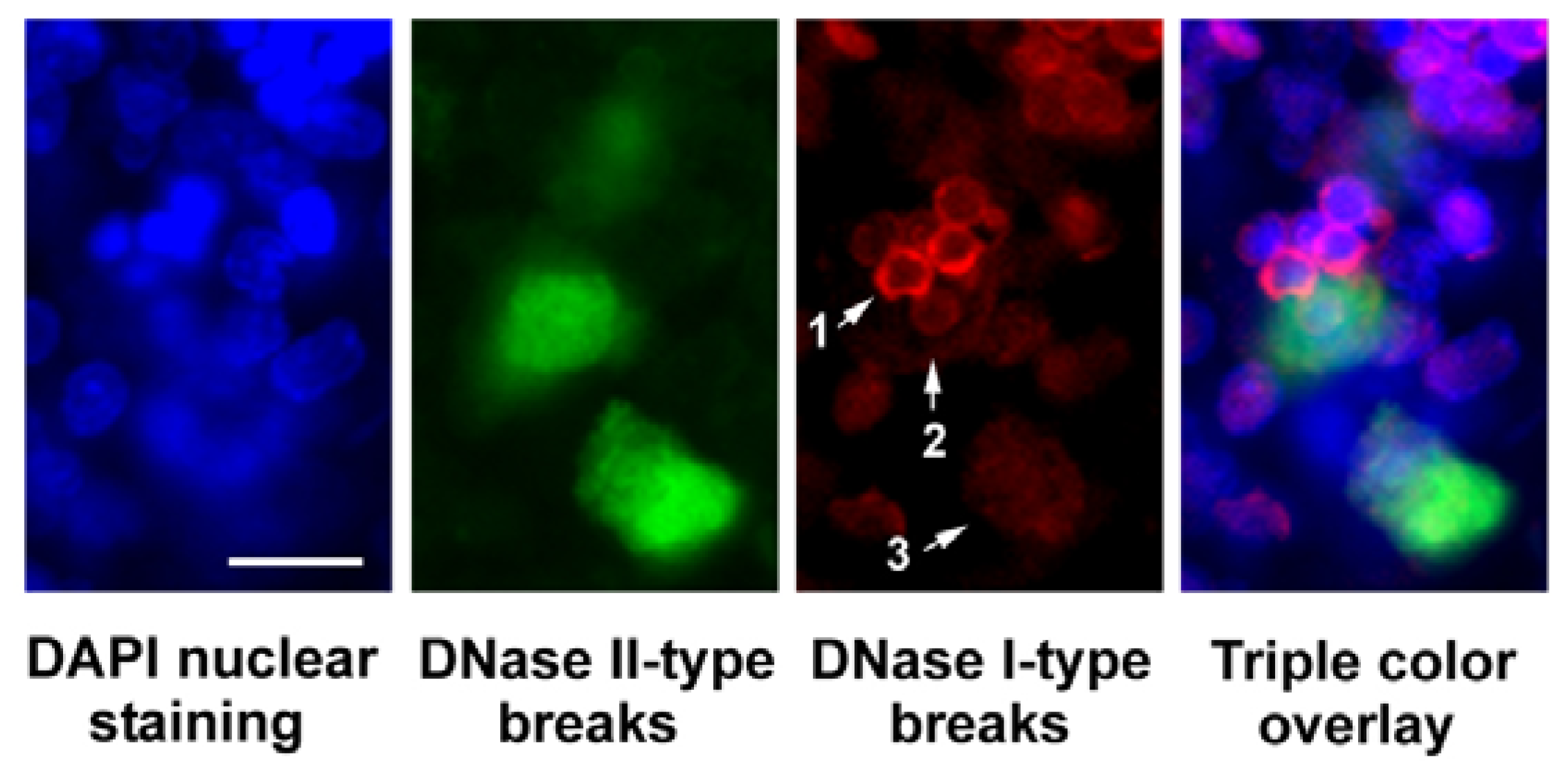
1.1. DNase II-Type Breaks and DNase I-Type Breaks in Apoptosis
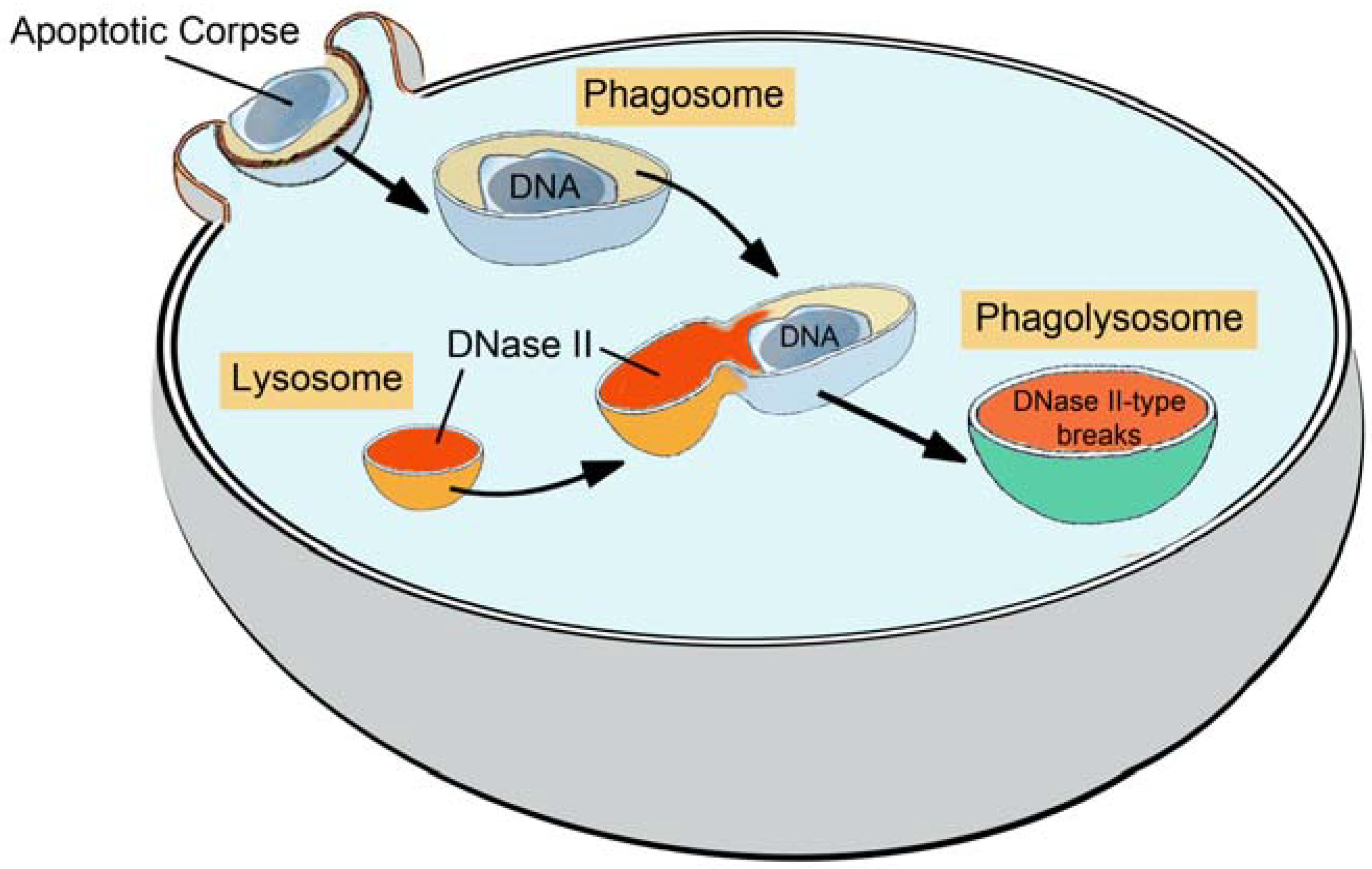
1.2. Sites of Generation of DNase II Breaks in Phagocytosis of Apoptotic Cells

1.3. Fluorescent Probes for Detection of DNase II Breaks In Situ

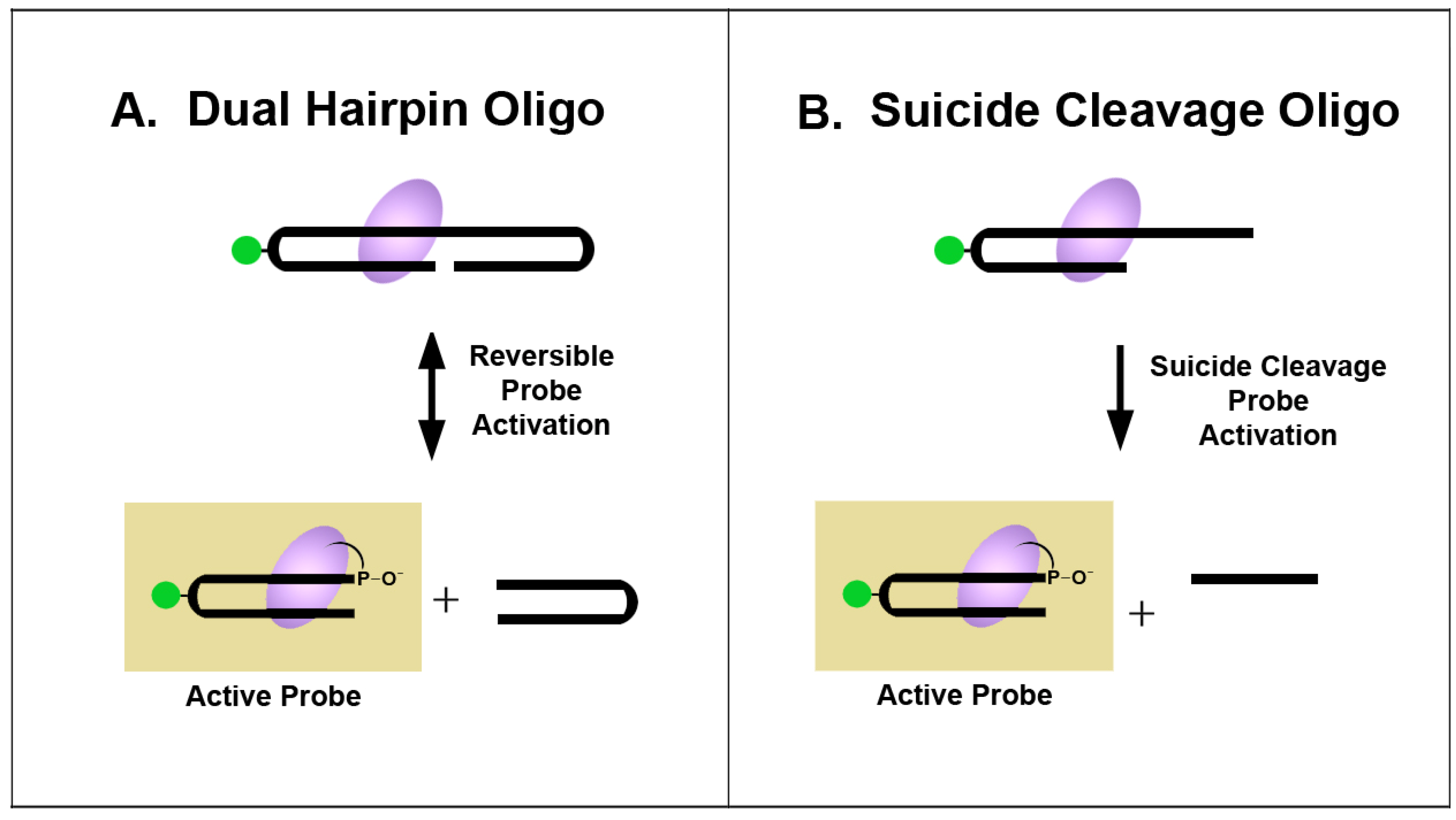
1.4. Approaches for Preparation of Reactive VACC TOPO Probes

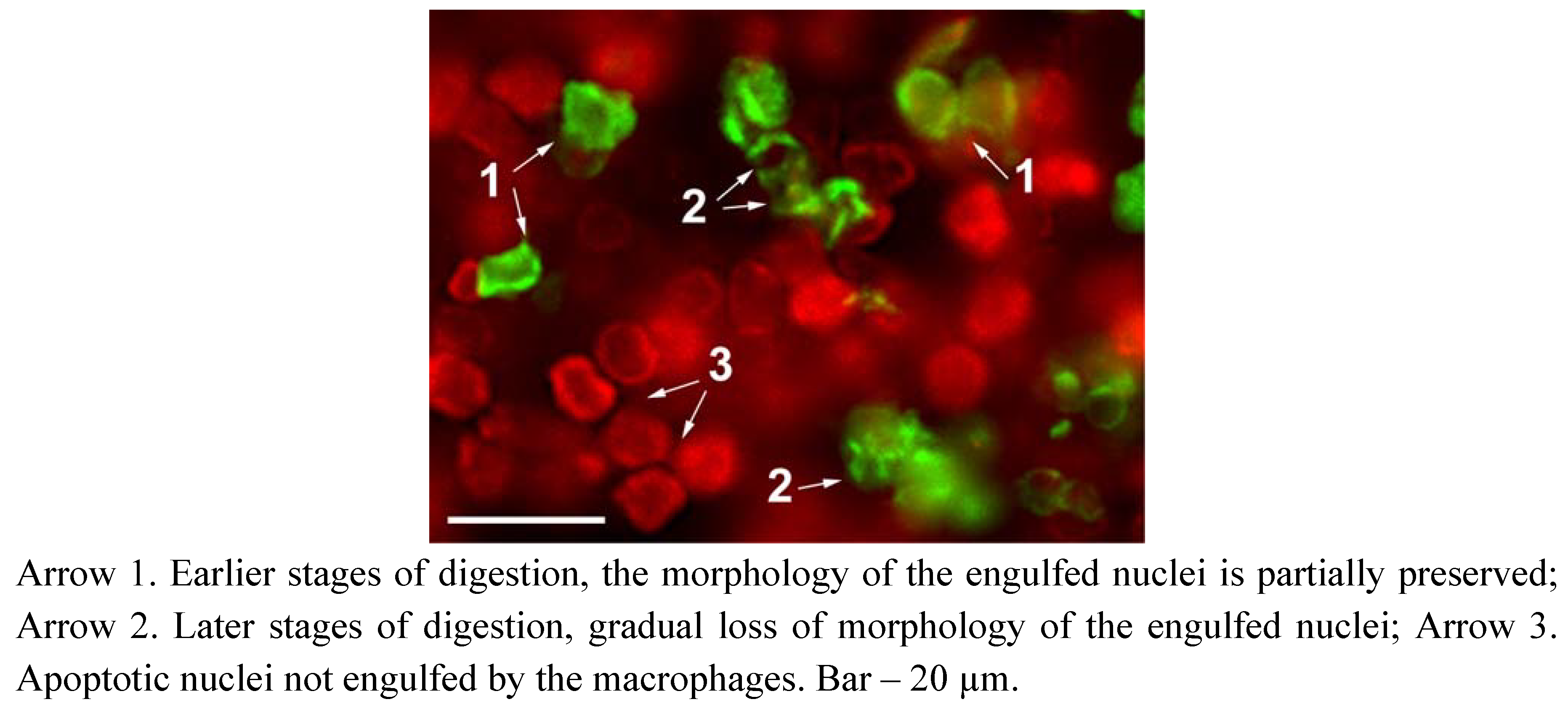
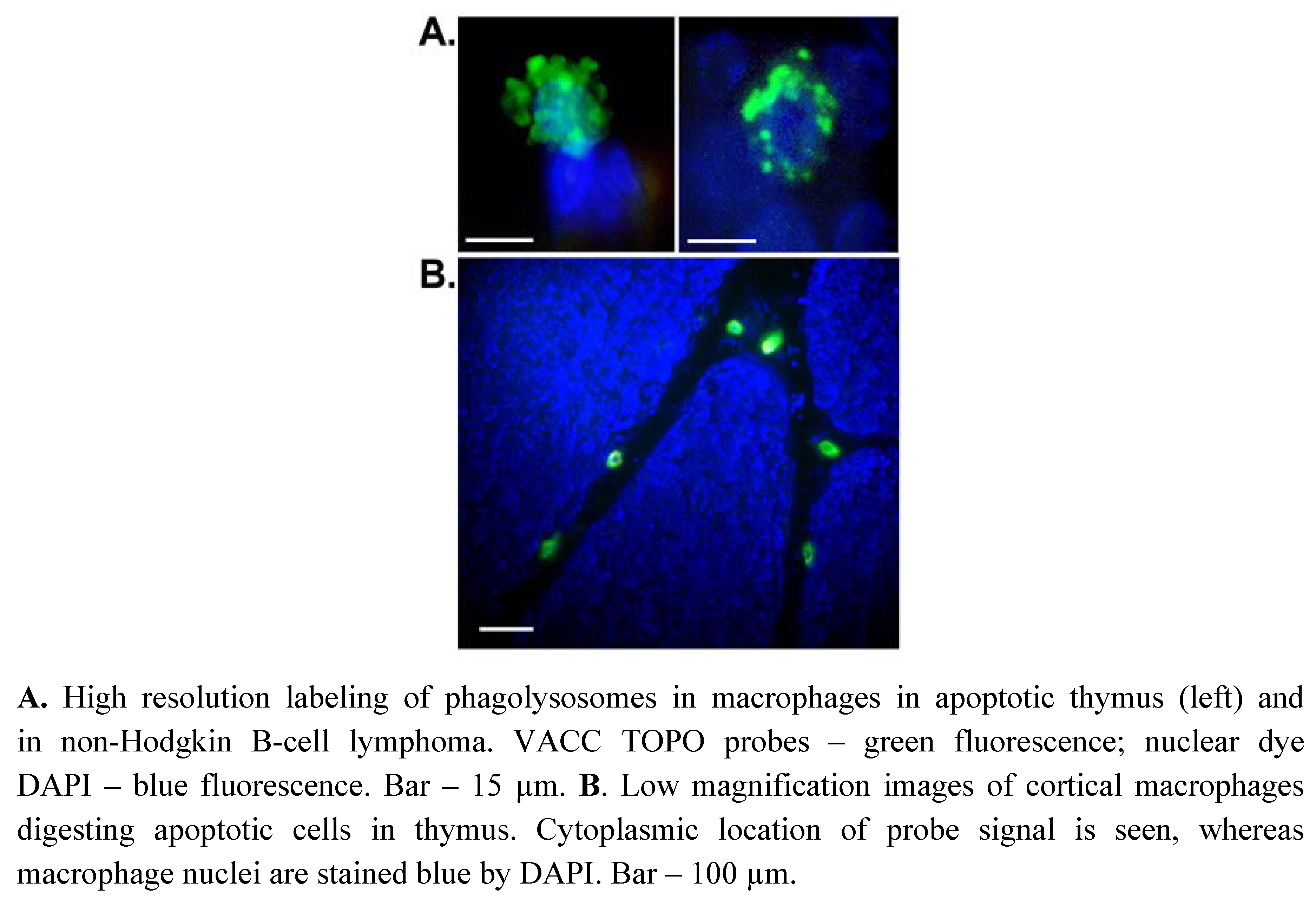
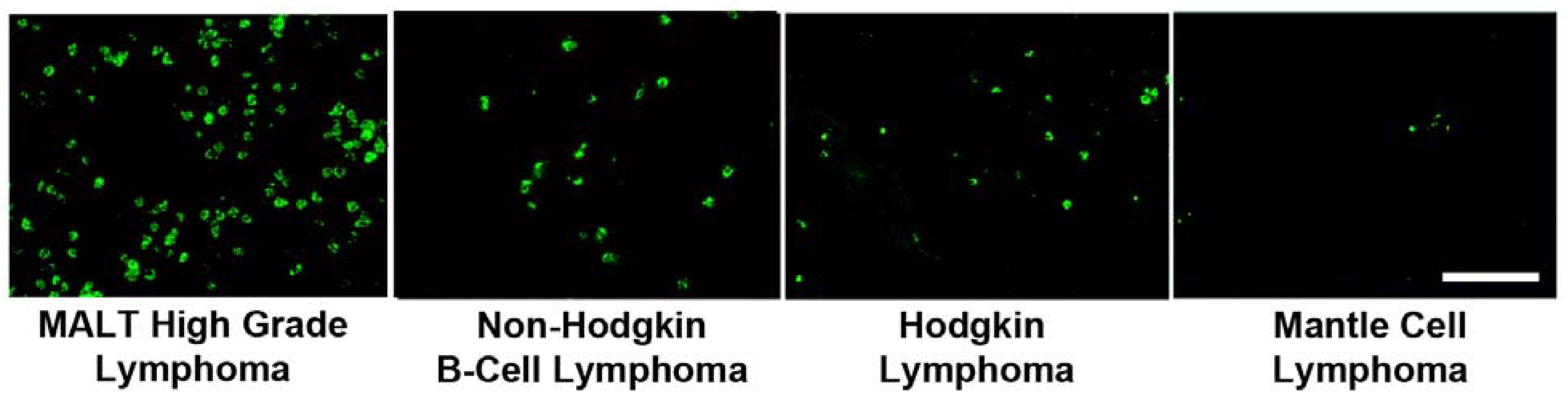
2. Results and Discussion





3. Experimental
3.1. Materials and Instruments

3.2. Preparation of Fluorescent Enzyme-Substrate Complexes and Labeling of Phagocytic Cells in Tissue Sections
4. Conclusion
Acknowledgments
References
- Samejima, K.; Earnshaw, W.C. Trashing the genome: Role of nucleases during apoptosis. Nat. Rev. Mol. Cell Biol. 2005, 6, 677–688. [Google Scholar] [CrossRef]
- Parrish, J.Z.; Xue, D. Cuts can kill: The roles of apoptotic nucleases in cell death and animal development. Chromosoma 2006, 115, 89–97. [Google Scholar] [CrossRef]
- Soehnlein, O.; Lindbom, L. Phagocyte partnership during the onset and resolution of inflammation. Nat. Rev. Immunol. 2010, 10, 427–439. [Google Scholar] [CrossRef]
- Fadeel, B.; Xue, D.; Kagan, V. Programmed cell clearance: molecular regulation of the elimination of apoptotic cell corpses and its role in the resolution of inflammation. Biochem. Biophys. Res. Commun. 2010, 396, 7–10. [Google Scholar]
- Litvack, M.L.; Palaniyar, N. Review: Soluble innate immune pattern-recognition proteins for clearing dying cells and cellular components: Implications on exacerbating or resolving inflammation. Innate Immun. 2010, 16, 191–200. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death: Recommendations of the nomenclature committee on cell death. Cell Death Differ. 2009, 16, 3–11. [Google Scholar]
- Jersmann, H.P.; Ross, K.A.; Vivers, S.; Brown, S.B.; Haslett, C.; Dransfield, I. Phagocytosis of apoptotic cells by human macrophages: Analysis by multiparameter flow cytometry. Cytometry A. 2003, 51, 7–15. [Google Scholar]
- Nagata, S.; Nagase, H.; Kawane, K.; Mukae, N.; Fukuyama, V. Degradation of chromosomal DNA during apoptosis. Cell Death Differ. 2003, 10, 108–116. [Google Scholar]
- Evans, C.J.; Aguilera, R.J. DNase II: Genes, enzymes and function. Gene 2003, 322, 1–15. [Google Scholar]
- Weir, A.F. Deoxyribonuclease I (EC 3.1.21.1) and II (EC 3.1.22.1). Methods Mol. Biol. 1993, 16, 7–16. [Google Scholar]
- Didenko, V.V. 5'OH DNA breaks in apoptosis and their labeling by topoisomerase-based approach. Methods Mol. Biol. 2011, 682, 77–87. [Google Scholar]
- Loo, D.T. In situ detection of apoptosis by the TUNEL assay: An overview of techniques. Methods Mol. Biol. 2011, 682, 3–13. [Google Scholar] [CrossRef]
- Thiry, M. In situ nick translation at the electron microscopic level. Methods Mol. Biol. 2002, 203, 121–130. [Google Scholar]
- Didenko, V.V. Detection of specific double-strand DNA breaks and apoptosis in situ using T4 DNA ligase. Methods Mol. Biol. 2002, 203, 143–151. [Google Scholar]
- Hornsby, P.J.; Didenko, V.V. In situ ligation: A decade and a half of experience. Methods Mol. Biol. 2011, 682, 49–63. [Google Scholar] [CrossRef]
- Didenko, V.V.; Hornsby, P.J. Presence of double-strand breaks with single-base 3’ overhangs in cells undergoing apoptosis but not necrosis. J. Cell Biol. 1996, 135, 1369–1376. [Google Scholar] [CrossRef]
- Widlak, P.; Li, P.; Wang, X.; Garrard, W.T. Cleavage preferences of the apoptotic endonuclease DFF40 (caspase-activated DNase or nuclease) on naked DNA and chromatin substrates. J. Biol. Chem. 2000, 275, 8226–8232. [Google Scholar]
- Haas, A. The phagosome: Compartment with a license to kill. Traffic 2007, 8, 311–330. [Google Scholar] [CrossRef]
- Kinchen, J.M.; Ravichandran, K.S. Phagosome maturation: Going through the acid test. Nat. Rev. Mol. Cell Biol. 2008, 9, 781–795. [Google Scholar] [CrossRef]
- Yates, R.M.; Hermetter, A.; Russell, D.G. The kinetics of phagosome maturation as a function of phagosome/lysosome fusion and acquisition of hydrolytic activity. Traffic 2005, 6, 413–420. [Google Scholar] [CrossRef]
- Kawane, K.; Fukuyama, H.; Kondoh, G.; Takeda, J.; Ohsawa, Y.; Uchiyama, Y.; Nagata, S. Requirement of DNase II for definitive erythropoiesis in the mouse fetal liver. Science 2001, 292, 1546–1549. [Google Scholar] [CrossRef]
- Kawane, K.; Nagata, S. Nucleases in programmed cell death. Methods Enzymol. 2008, 442, 271–287. [Google Scholar] [CrossRef]
- Shuman, S. Vaccinia virus DNA topoisomerase: A model eukaryotic type IB enzyme. Biochim. Biophys. Acta 1998, 1400, 321–337. [Google Scholar]
- Shuman, S. Site-specific interaction of vaccinia virus topoisomerase I with duplex DNA. J. Biol. Chem. 1991, 266, 11372–11379. [Google Scholar]
- Shuman, S. Site-specific cleavage by vaccinia virus DNA topoisomerase I. J. Biol. Chem. 1991, 266, 1796–1803. [Google Scholar]
- Maunders, M.J. DNA and RNA ligases (EC 6.5.1.1, EC 6.5.1.2, and EC 6.5.1.3). Methods Mol. Biol. 1993, 16, 213–230. [Google Scholar]
- Didenko, V.V.; Minchew, C.L.; Shuman, S.; Baskin, D.S. Semi-artificial fluorescent molecular machine for DNA damage detection. Nano Letters 2004, 4, 2461–2466. [Google Scholar]
- Shuman, S. Two classes of DNA end-joining reactions catalyzed by vaccinia topoisomerase I. J. Biol. Chem. 1992, 267, 16755–16758. [Google Scholar]
- Sigma-Aldrich, Nucleases, Deoxyribonuclease II. Available online: http://www.sigmaaldrich.com/life-science/metabolomics/enzyme-explorer/learning-center/nucleases.html.
- Walker, P.R.; Leblanc, J.; Carson, C.; Ribecco, M.; Sikorska, M. Neither caspase-3 nor DNA fragmentation factor is required for high molecular weight DNA degradation in apoptosis. Ann. N.Y. Acad. Sci. 1999, 887, 48–59. [Google Scholar]
- Hermann, M.; Niemitz, C.; Marafioti, T.; Schriever, F. Reduced phagocytosis of apoptotic cells in malignant lymphoma. Int. J. Cancer 1998, 75, 675–679. [Google Scholar]
- Hancock, B.W.; Bruce, L.; Richmon, J. Neutrophil function in lymphoreticular malignancy. Br. J. Cancer 1976, 33, 496–500. [Google Scholar]
- Sheagren, J.N.; Block, J.B.; Wolff, S.M. Reticuloendothelial system phagocytic function in patients with Hodgkin’s disease. J. Clin. Invest. 1967, 46, 855–862. [Google Scholar] [CrossRef]
- Du, M.; Peng, H.; Singh, N.; Isaacson, P.G.; Pan, L. The accumulation of p53 abnormalities is associated with progression of mucosa-associated lymphoid tissue lymphoma. Blood 1995, 86, 4587–4593. [Google Scholar]
- Allen, L.H. Modulating phagocyte activation: The pros and cons of Helicobacter pylori virulence factors. J. Exp. Med. 2000, 191, 1451–1454. [Google Scholar] [CrossRef]
- Rassidakis, G.Z.; Jones, D.; Thomaides, A.; Sen, F.; Lai, R.; Cabanillas, F.; McDonnell, T.J.; Medeiros, L.J. Apoptotic rate in peripheral T-cell lymphomas a study using a tissue microarray with validation on full tissue sections. Am. J. Clin. Pathol. 2002, 118, 328–334. [Google Scholar]
- Guidoboni, M.; Doglioni, C.; Laurino, L; Boiocchi, M.; Dolcetti, R. Activation of infiltrating cytotoxic T lymphocytes and lymphoma cell apoptotic rates in gastric MALT lymphomas. Differences between high-grade and low-grade cases. Am. J. Pathol. 1999, 155, 823–829. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Minchew, C.L.; Didenko, V.V. Fluorescent Probes Detecting the Phagocytic Phase of Apoptosis: Enzyme-Substrate Complexes of Topoisomerase and DNA. Molecules 2011, 16, 4599-4614. https://doi.org/10.3390/molecules16064599
Minchew CL, Didenko VV. Fluorescent Probes Detecting the Phagocytic Phase of Apoptosis: Enzyme-Substrate Complexes of Topoisomerase and DNA. Molecules. 2011; 16(6):4599-4614. https://doi.org/10.3390/molecules16064599
Chicago/Turabian StyleMinchew, Candace L., and Vladimir V. Didenko. 2011. "Fluorescent Probes Detecting the Phagocytic Phase of Apoptosis: Enzyme-Substrate Complexes of Topoisomerase and DNA" Molecules 16, no. 6: 4599-4614. https://doi.org/10.3390/molecules16064599
APA StyleMinchew, C. L., & Didenko, V. V. (2011). Fluorescent Probes Detecting the Phagocytic Phase of Apoptosis: Enzyme-Substrate Complexes of Topoisomerase and DNA. Molecules, 16(6), 4599-4614. https://doi.org/10.3390/molecules16064599