Crystallization and Characterization of an Inflammatory Lectin Purified from the Seeds of Dioclea wilsonii

Abstract
:1. Introduction
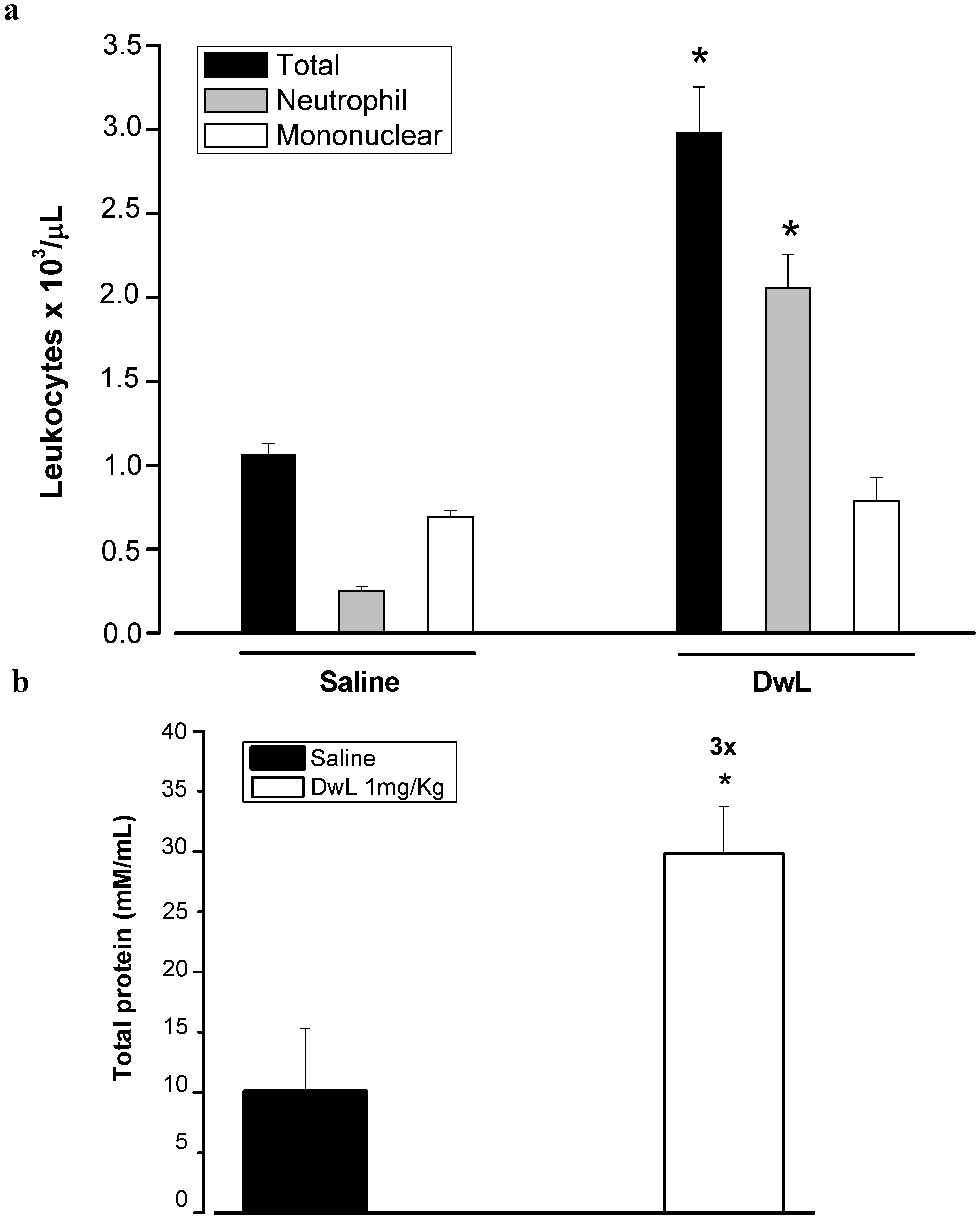
2. Results and Discussion

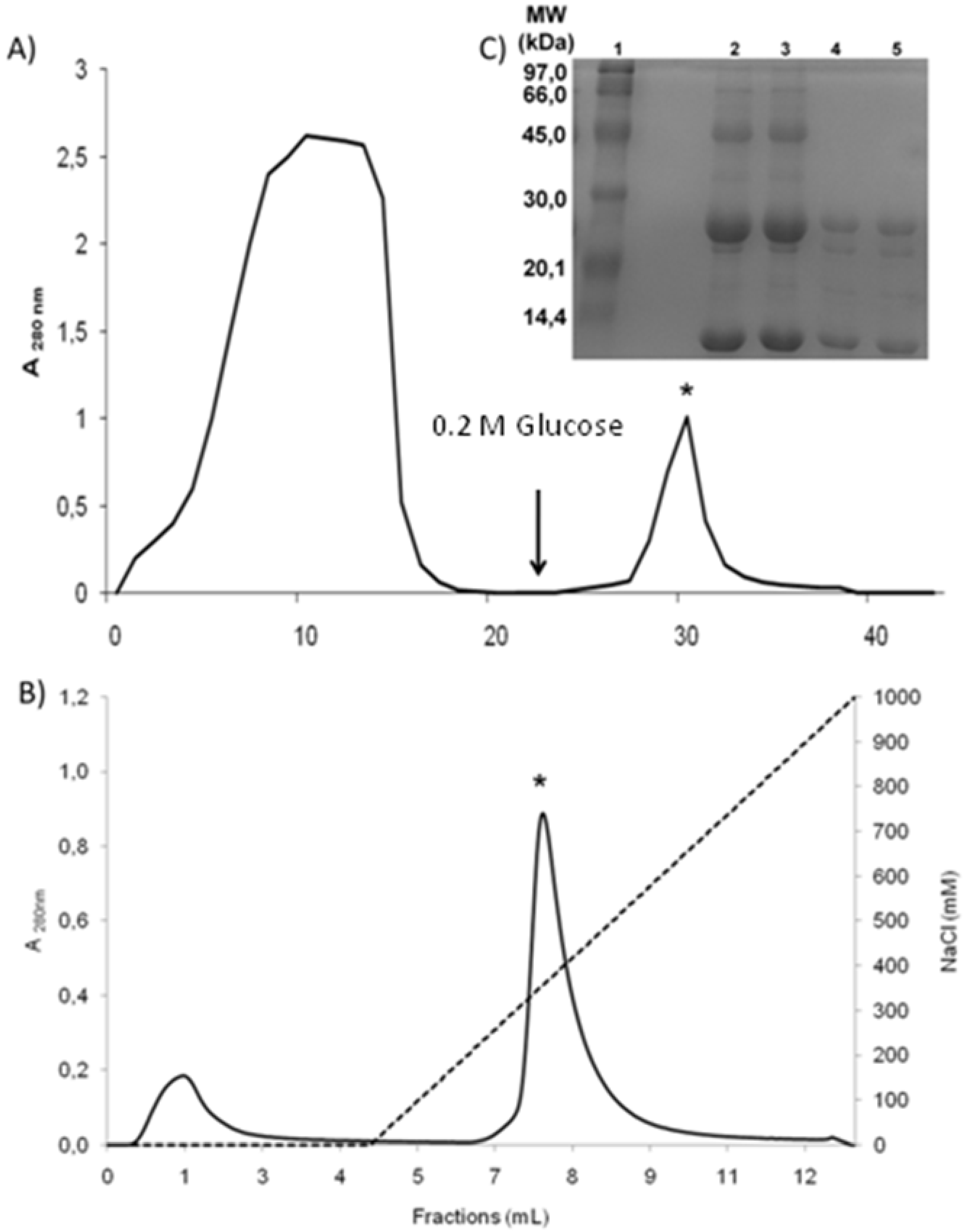{kind=link}
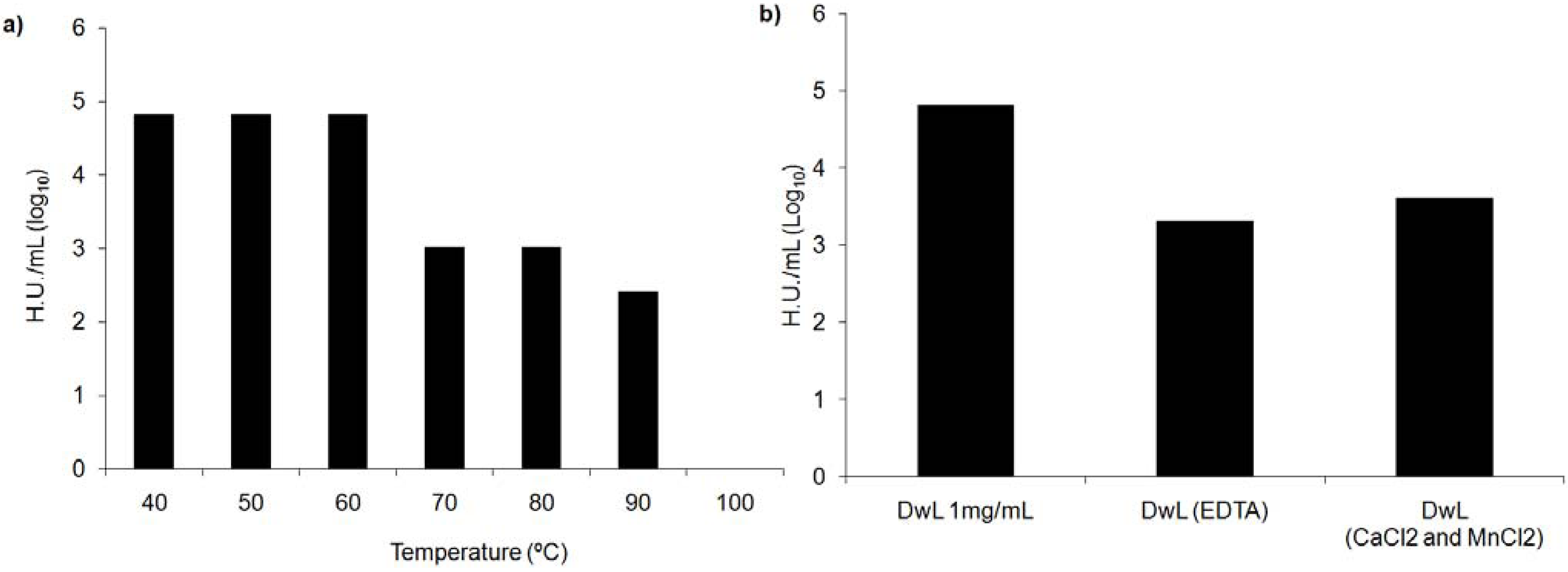{kind=link}
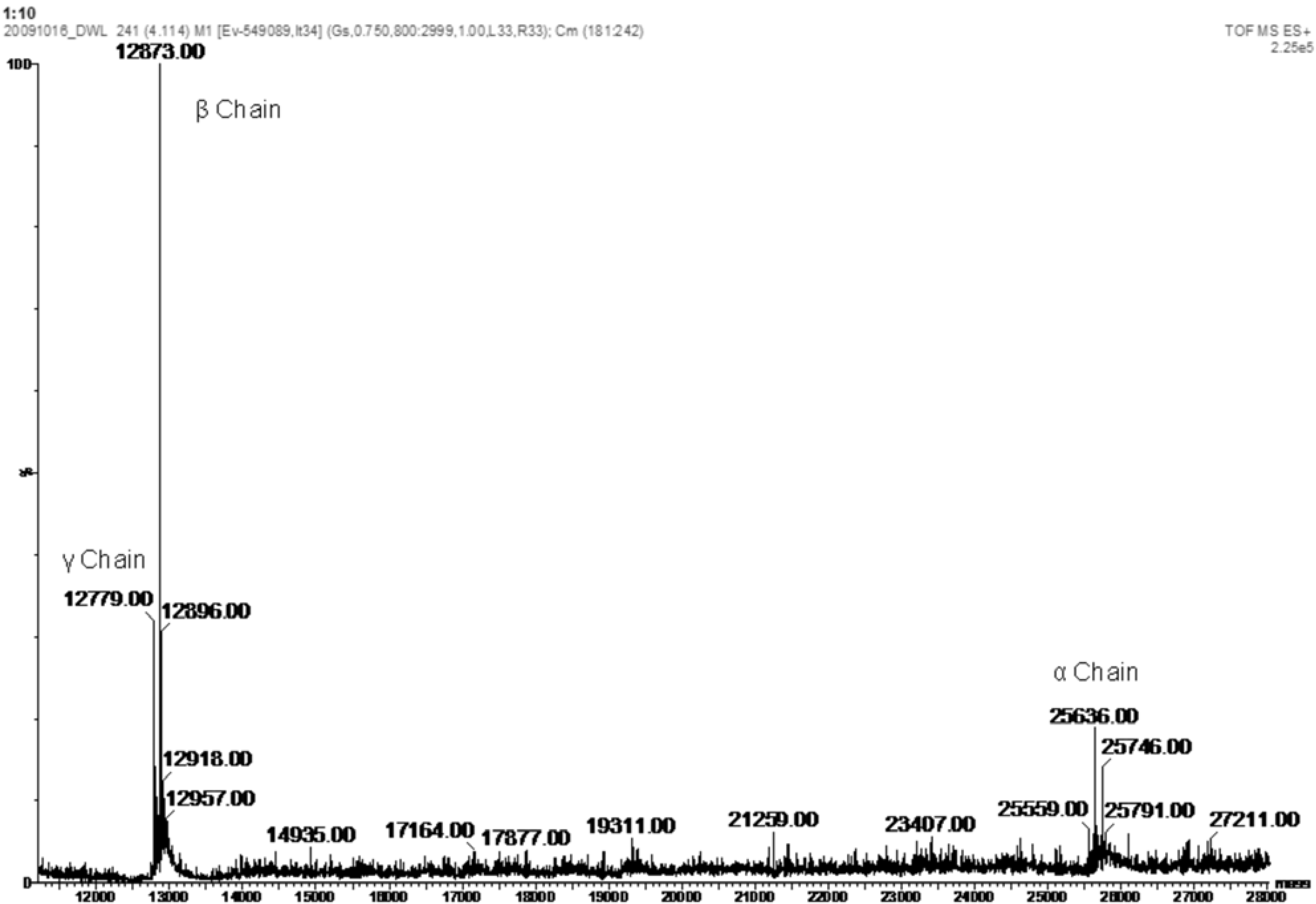{kind=link}
{kind=link}
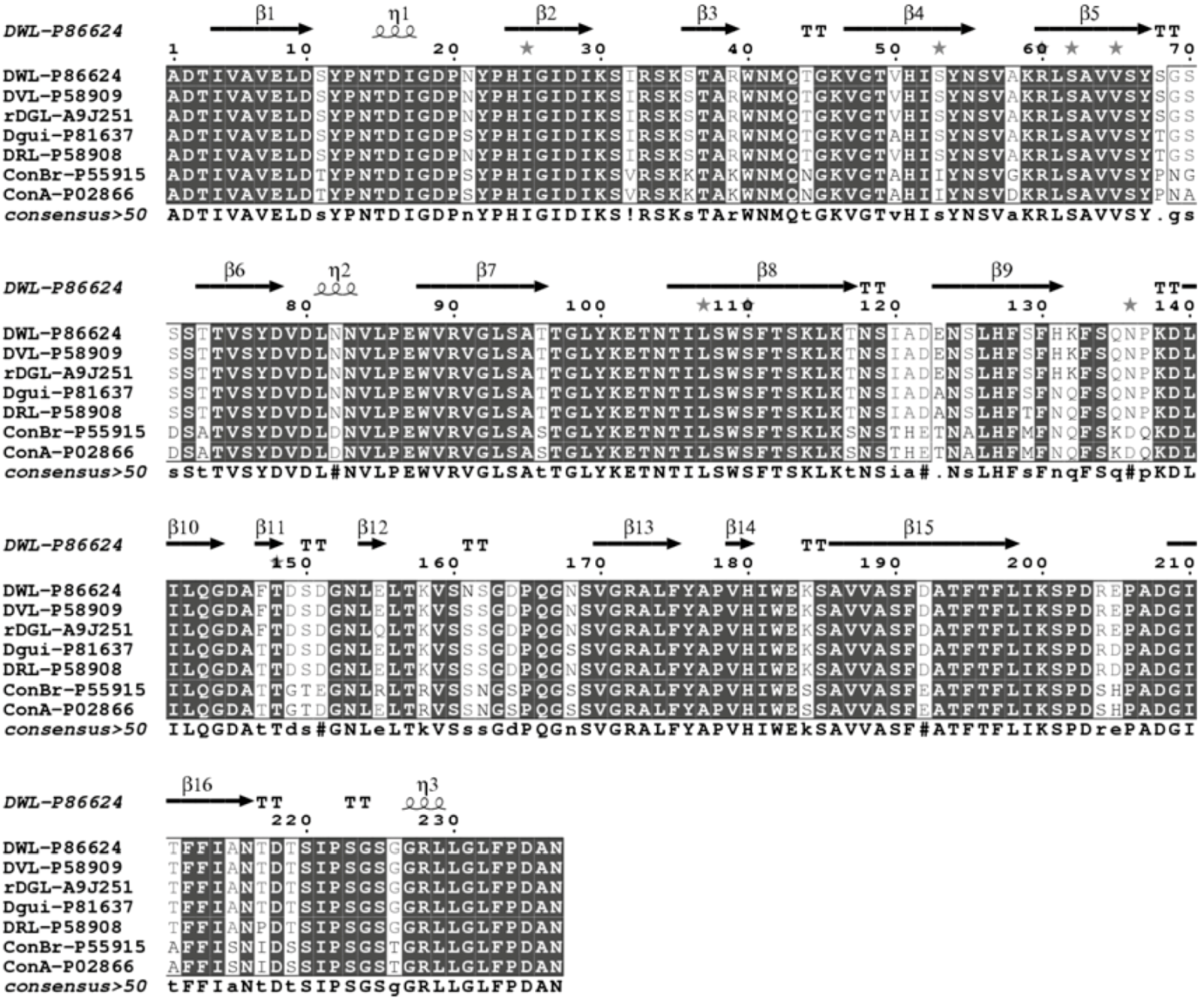{kind=link}
{kind=link}
{kind=link}
| Fractions * | V (mL) | Protein | Hemagglutinating activity † | Minimum Concentration(mg/mL) | Purification | ||||
|---|---|---|---|---|---|---|---|---|---|
| mg/mL | Total | % | U.H./mL | Total | Specific | ||||
| Total Extract | 30 | 145.62 | 4368.6 | 100 | 131072 | 3932160 | 900.09 | 1.11 × 10−3 | 1 |
| Sephadex G-50 PII | 14 | 4.26 | 59.64 | 1.36 | 32768 | 458752 | 7692.02 | 1.3 × 10−4 | 8.5 |
| HiTrap SP XL PII | 2 | 1.127 | 2.254 | 0.77 | 65536 | 131072 | 50150.85 | 1.3 × 10−5 | 55.72 |




| Peptide | Experimental Mass (Da) | Sequence |
|---|---|---|
| T1 | 3254.5566 | ADTIVAVELDSYPNTDIGDPNYPHIGIDIK |
| T2 | 1373.7165 | VGTVHISYNSVAK |
| T3 | 1108.5643 | VGLSATTGLYK |
| T4 | 1512.7244 | ETNTILSWSFTSK |
| T5 | 1845.8464 | TNSIADENSLHFSFHK |
| T6 | 2865.4045 | FSQNPKDLILQGDAFTDSDGNLELTK |
| T7 | 1472.7366 | ALFYAPVHIWEK |
| T8 | 1715.9243 | SAVVASFDATFTFLIK |
| T9MS | 2864.2444 | SPDREPADGITFFIANTDTSIPSGSGGR |
| Q1 | 3515.7151 | DSYPNTDIGDPNYPHIGIDIKSIRSKSTARW |
| Q2 | 2303.1687 | MNQTGKVGTVHISYNSVAKRL |
| Q3 | 1394.7444 | NNVLPEWVRVGL |
| Q4 | 1090.5643 | KETNTILSW |
| Q5 | 1785.8522 | TKVSNSGDPQGNSVGRAL |
| Q6 | 1544.7365 | IKSPDREPADGITF |
| Q7 | 1691.8844 | FIANTDTSIPSGSGGRL |


| Data Collection * | |
|---|---|
| Point group | I222 |
| Unit cell parameters (Å) | |
| A | 59.6 |
| B | 67.9 |
| C | 109.0 |
| Unit cell angles (°) | |
| α = β = γ | 90 |
| Total number of reflection | 50274 (6863) |
| Total number of unique observations | 10483 (1526) |
| Resolution limit (Å) | 23.29–2.2 |
| Rmerge (%) a | 8.4 (24.1) |
| Completeness (%) | 97.1 (98.5) |
| Multiplicity (%) | 4.8 (4.5) |
| (I)/σ | 5.0 (1.8) |
| Matthews coefficient (Å3.Da−1) | 2.17 |
| Solvent content (%) | 43.28 |
| Wavelenght (Å) | 1.42 |
 where I(hkl)i is the intensity of ith measurement of the reflection h and I(hkl) is the mean value of the I(hkl)i for all I measurements.
where I(hkl)i is the intensity of ith measurement of the reflection h and I(hkl) is the mean value of the I(hkl)i for all I measurements. 
3. Experimental
3.1. Materials
3.2. Protein Extraction Procedure
3.3. Hemagglutinating Activity
3.4. Sugar Specificity
3.5. Protein Purification
3.6. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
3.7. Lectin Metal Dependence
3.8. Lectin Thermostability
3.9. MW Determination by Mass Spectrometry
3.10. Protein Digestion and Tandem Mass Spectrometry Analysis
3.11. Primary Structure Analysis
3.12. Crystallization and Data Collection
3.13. Biological Assays
3.13.1. Animals, Drugs and Reagents
3.13.2. Peritonitis Model
3.13.3. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Kocourek, J.; Horejsi, V. A note of the recent discussion on definition of the term “lectin”. In Lectins: Biology, Biochemistry, Clinical Biochemistry; Bog-Hansen, T.C., Spengler, G.A., Eds.; Walter de Gruyter: Berlin, Germany, 1983; Volume 3, pp. 3–6. [Google Scholar]
- Moreira, R.A.; Ainouz, I.L.; Oliveita, J.T.A.; Cavada, B.S. Plant lectins: Chemical and biological aspects. Mem. Inst. Oswaldo Cruz 1991, 86, 212–218. [Google Scholar]
- Gabius, H.J.; Gabius, S. Glycoscience. Status and Perspectives; Chapman and Hall: Weinheim, Germany, 1997. [Google Scholar]
- Oliveira, T.M.; Delatorre, P.; Rocha, B.A.M.; Souza, E.P.; Nascimento, K.S.; Bezerra, G.A.; Moura, T.R.; Benevides, R.G.; Bezerra, E.H.S.; Moreno, F.B.M.B.; et al. Crystal structure of Dioclea rostrata: Insights into understanding the pH-dependent dimer-tetramer equilibrium and the structural basis for carbohydrate recognition in Diocleinae lectins. J. Struct. Biol. 2008, 164, 177–182. [Google Scholar] [CrossRef]
- Etzler, M.E. Distribution and function of plant lectins. In The Lectins: Properties, Functions and Applications in Biology and Medicine; Liener, J.E., Sharon, N., Goldstein, I.J., Eds.; Academic Press: New York, NY, USA, 1986; pp. 371–435. [Google Scholar]
- Cunningham, B.A.; Hemperly, J.J.; Hopp, T.P.; Edelman, G.M. Favin versus concanavalin A: Circularly permuted amino acid sequences. Proc. Natl. Acad. Sci. USA 1979, 76, 3218–3222. [Google Scholar] [CrossRef]
- Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Grangeiro, T.B. Primary structures and functions of plant lectins. Braz. J. Fis. Veg. 1993, 5, 193–201. [Google Scholar]
- Barral-Netto, M.; Santos, S.B.; Barral, A.; Moreira, L.I.; Santos, C.F.; Moreira, R.A.; Oliveira, J.T.A.; Cavada, B.S. Human lymphocyte stimulation by lectins from Brazilian leguminous seeds. Immunol. Invest. 1992, 21, 297–303. [Google Scholar] [CrossRef]
- Gomes, J.C.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A. Histamine release induced by glucose (mannose)-specific lectins isolated from brazilian plants, comparison with concanavalin A. Agents Actions 1994, 41, 132–135. [Google Scholar] [CrossRef]
- Bento, C.A.M.; Cavada, B.S.; Oliveira, J.T.A.; Moreira, R.A.; Barja-Fidalgo, C. Rat paw edema and leukocyte immigration induced by plant lectins. Agents Actions 1993, 38, 48–54. [Google Scholar] [CrossRef]
- Rodriguez, D.; Cavada, B.S.; Oliveira, J.T.A.; Moreira, R.A.; Russo, M. Differences in macrophage stimulation and leukocyte accumulation in response to intraperitoneal administration of glucose/mannose-binding plant lectins. Braz. J. Med. Biol. Res. 1992, 25, 823–826. [Google Scholar]
- Cavada, B.S.; Grangeiro, T.B.; Ramos, M.V.; Crisostomo, C.V.; Silva, L.M.; Moreira, R.A.; Oliveira, J.T.A. Lectin from Dioclea guianensis var. lasiophylla Duke seeds mobilization during germination and seedlings growth in the dark. Braz. J. Veg. Fis. 1994, 6, 21–25. [Google Scholar]
- Hora, R.C.; Soares, J.J. Estrutura fitossociológica da comunidade de lianas em uma floresta semidecidual na Fazenda Canchim, São Paulo, SP. Braz. J. Bot. 2002, 25, 323–329. [Google Scholar] [CrossRef]
- Moreira, R.A.; Monteiro, A.C.O.; Horta, A.C.G.; Oliveira, J.T.A.; Cavada, B.S. Isolation and characterization of Dioclea altissima Var. Megacarpa seed lectin. Phytochemstry 1997, 46, 139–144. [Google Scholar]
- Cavada, B.S.; Grangeiro, T.B.; Ramos, M.V.; Cordeiro, E.F.; Oliveira, J.T.A.; Moreira, R.A. Isolation and partial characterization of a lectin from Dioclea rostrata Benth seeds. Braz. J. Veg. Fis. 1996, 8, 31–36. [Google Scholar]
- Moreira, R.A.; Cordeiro, E.F.; Ramos, M.V.; Grangeiro, T.B.; Martins, J.L.; Oliveira, J.T.A.; Cavada, B.S. Isolation and partial characterization of a lectin from seeds of Dioclea violacea. Braz. J. Veg. Fis. 1996, 8, 23–29. [Google Scholar]
- Cavada, B.S.; Ramos, M.V.; Cordeiro, E.F.; Grangeiro, T.B.; Oliveira, J.T.A.; Carvalho, A.F.F.U.; Moreira, R.A. Purification and partial characterization of a lectin from Dioclea virgata Benth seeds. Braz. J. Veg. Fis. 1996, 8, 37–42. [Google Scholar]
- Vasconcellos, I.M.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A. Purification and partial characterization of a lectin from the seed of Dioclea guianensis. J. Food Biochem. 1991, 15, 137–154. [Google Scholar] [CrossRef]
- Moreira, R.A.; Barros, A.C.H.; Stewart, J.C.; Pusztai, A. Isolation and characterization of a lectin from the seeds of Dioclea grandiflora (Mart.). Planta 1983, 158, 63–69. [Google Scholar] [CrossRef]
- Moreira, R.A.; Ainouz, I.L.; Oliveira, J.T.A.; Cavada, B.S. Plant lectins: Chemical and biological aspects. Mem. Inst. Osw. Cruz 1991, 86, 212–218. [Google Scholar]
- Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Granjeiro, T.B. Primary structures and functions of plant lectins. Braz. J. Veg. Fis. 1993, 5, 193–201. [Google Scholar]
- Summer, J.B.; Howell, S.F. The identification of the hemaglutinin of the jack bean with Concavalin A. J. Bact. 1937, 32, 227–237. [Google Scholar]
- Moreira, R.A.; Cavada, B.S. Lectin from Canavalia brasiliensis (Mart.). Isolation, characterization and behavior during germination. Biol. Plantarum 1984, 26, 113–120. [Google Scholar] [CrossRef]
- Oliveira, J.T.A.; Cavada, B.S.; Moreira, R.A. Isolation and partial characterization of a lectin from Cratylia floribunda seeds. Braz. J. Bot. 1991, 14, 61–66. [Google Scholar]
- Perez, G.; Perez, C.; Cavada, B.S.; Moreira, R.A.; Richardson, M. Comparison of the amino acid sequences of the lectins from the seeds of Dioclea lehmanni and Canavalia maritima. Phytochemistry 1991, 30, 2619–2621. [Google Scholar] [CrossRef]
- Carrington, D.M.; Auffret, A.; Hanke, D.E. Polypeptide ligation occurs during post-translational modification of Concanavalin A. Nature 1985, 313, 64–67. [Google Scholar] [CrossRef]
- Naismith, J.H.; Emmerich, C.; Habash, J.; Harrop, S.J.; Helliwell, J.R.; Hunter, W.N.; Raftery, J.; Kalb, A.J.; Yariv, J. Refined structure of concanavalin A complexed with methyl-D-mannopyranoside at 2.0 Å resolution and comparison with the saccharide-free structure. Acta Crystal. D 1994, 50, 847–858. [Google Scholar] [CrossRef]
- Matthews, B.W. The solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–487. [Google Scholar] [CrossRef]
- Assreuy, A.M.; Fontenele, S.R.; Pires, A.F.; Fernandes, D.C.; Rodrigues, N.V.; Bezerra, E.H.; Moura, T.R.; do Nascimento, K.S.; Cavada, B.S. Vasodilator effects of Diocleinae lectins from Canavalia genus. N-S Arch. Pharmacol. 2009, 380, 509–521. [Google Scholar] [CrossRef]
- Andrade, J.L.; Arruda, S.; Barbosa, T.; Paim, L.; Ramos, M.V.; Cavada, B.S.; Barral-Netto, M. Lectin-induced Nitric Oxide production. Cel. Immunol. 1999, 194, 98–102. [Google Scholar] [CrossRef]
- Barbosa, T.; Arruda, S.; Cavada, B.; Grangeiro, T.B.; de Freitas, L.A.; Barral-Netto, M. In vivo lymphocyte activation and apoptosis by lectins of the Diocleinae subtribe. Mem. Inst. Osw. Cruz 2001, 96, 673–678. [Google Scholar] [CrossRef]
- Cavada, B.S.; Barbosa, T.; Arruda, S.; Grangeiro, T.B.; Barral-Netto, M. Revisiting proteus: Do minor changes in lectin structure matter in biological activity? Lessons from and potential biotechnological uses of the Diocleinae subtribe lectins. Curr. Prot. Pept. Sci. 2001, 2, 123–135. [Google Scholar] [CrossRef]
- Pelletier, M.; Lavastre, V.; Savoie, A.; Ratthé, C.; Saller, R.; Hostanska, K.; Girard, D. Modulation of interleukin-15-induced human neutrophil responses by the plant lectin Viscum album agglutinin-I. Clin. Immun. 2001, 101, 229–236. [Google Scholar] [CrossRef]
- Figueiredo, J.G.; Bitencourt, F.S.; Mota, M.R.; Silvestre, P.P.; Aguiar, C.N.; Benevides, R.G.; Nascimento, K.S.; Moura, T.R.; Dal-Secco, D.; Assreuy, A.M.; et al. Pharmacological analysis of the neutrophil migration induced by D. rostrata lectin: involvement of cytokines and nitric oxide. Toxicon 2009, 54, 736–744. [Google Scholar] [CrossRef]
- Northover, A.M.; Northover, B.J. Lectin-induced increase in microvascular permeability to colloidal carbon in vitro may involve protein kinase C activation. Agents Actions 1994, 42, 136–139. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Biochemistry 1976, 72, 248–254. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Neuhoff, V.; Arold, N.; Taube, D.; Ehrhardt, W. Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis 1988, 9, 255–262. [Google Scholar] [CrossRef]
- Ferrige, A.G.; Seddon, M.J.; Green, B.N.; Jarvis, S.A.; Skilling, J. Disentangling electrospray spectra with maximum entropy. Rapid Commun. Mass Spectrom. 1992, 6, 707–711. [Google Scholar] [CrossRef]
- Altschul, S.F.; Maddem, T.L.; Scharffer, A.A.; Zhang, L.; Zhang, Z.; Miller, W.; Lipman, D.L. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Gouet, P.; Robert, X.; Courcelle, E. ESPript/ENDscript: Extracting and rendering sequence and 3D information from atomic structure of proteins. Nucleic Acids Res. 2003, 31, 3320–3323. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Jancarik, J.; Kim, S.H. Sparse matrix sampling: a screening method for crystallization of proteins. J. Appl. Cryst. 1991, 24, 409–441. [Google Scholar] [CrossRef]
- Leslie, A.G.W. Recent changes to the MOSFLM package for processing film and image plate data. Joint CCP4 + ESF-EAMCB. Newsl. Prot. Cryst. 1992. No. 26. [Google Scholar]
- Evans, P.R. Scala. Joint CCP4 and ESF-EACBM Newsletter 1997, 33, 22–24. [Google Scholar]
- Souza, G.E.P.; Ferreira, S.H. Blockade by antimacrophages serum of the migration of PMN neutrophils into the inflamed peritoneal cavity. Agents Actions 1985, 17, 97–103. [Google Scholar] [CrossRef]
- Sample Availability: Contact the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rangel, T.B.A.; Assreuy, A.M.S.; Pires, A.d.F.; Carvalho, A.U.d.; Benevides, R.G.; Simões, R.d.C.; Silva, H.C.d.; Bezerra, M.J.B.; Nascimento, A.S.F.d.; Nascimento, K.S.d.; et al. Crystallization and Characterization of an Inflammatory Lectin Purified from the Seeds of Dioclea wilsonii. Molecules 2011, 16, 5087-5103. https://doi.org/10.3390/molecules16065087
Rangel TBA, Assreuy AMS, Pires AdF, Carvalho AUd, Benevides RG, Simões RdC, Silva HCd, Bezerra MJB, Nascimento ASFd, Nascimento KSd, et al. Crystallization and Characterization of an Inflammatory Lectin Purified from the Seeds of Dioclea wilsonii. Molecules. 2011; 16(6):5087-5103. https://doi.org/10.3390/molecules16065087
Chicago/Turabian StyleRangel, Thaiz Batista Azevedo, Ana Maria Sampaio Assreuy, Alana de Freitas Pires, Amanda Uliana de Carvalho, Raquel Guimarães Benevides, Rafael da Conceição Simões, Helton Colares da Silva, Maria Júlia Barbosa Bezerra, Antonia Samia Fernandes do Nascimento, Kyria Santiago do Nascimento, and et al. 2011. "Crystallization and Characterization of an Inflammatory Lectin Purified from the Seeds of Dioclea wilsonii" Molecules 16, no. 6: 5087-5103. https://doi.org/10.3390/molecules16065087
APA StyleRangel, T. B. A., Assreuy, A. M. S., Pires, A. d. F., Carvalho, A. U. d., Benevides, R. G., Simões, R. d. C., Silva, H. C. d., Bezerra, M. J. B., Nascimento, A. S. F. d., Nascimento, K. S. d., Nagano, C. S., Sampaio, A. H., Delatorre, P., Rocha, B. A. M. d., Fernandes, P. M. B., & Cavada, B. S. (2011). Crystallization and Characterization of an Inflammatory Lectin Purified from the Seeds of Dioclea wilsonii. Molecules, 16(6), 5087-5103. https://doi.org/10.3390/molecules16065087