Physicochemical Characteristics and Immunomodulatory Activities of Three Polysaccharide-Protein Complexes of Longan Pulp

Abstract
:1. Introduction
2. Results and Discussion
2.1. Physicochemical characteristics of LP1-3
2.1.1. Characterization of polysaccharide-protein complex


2.1.2. Chemical composition

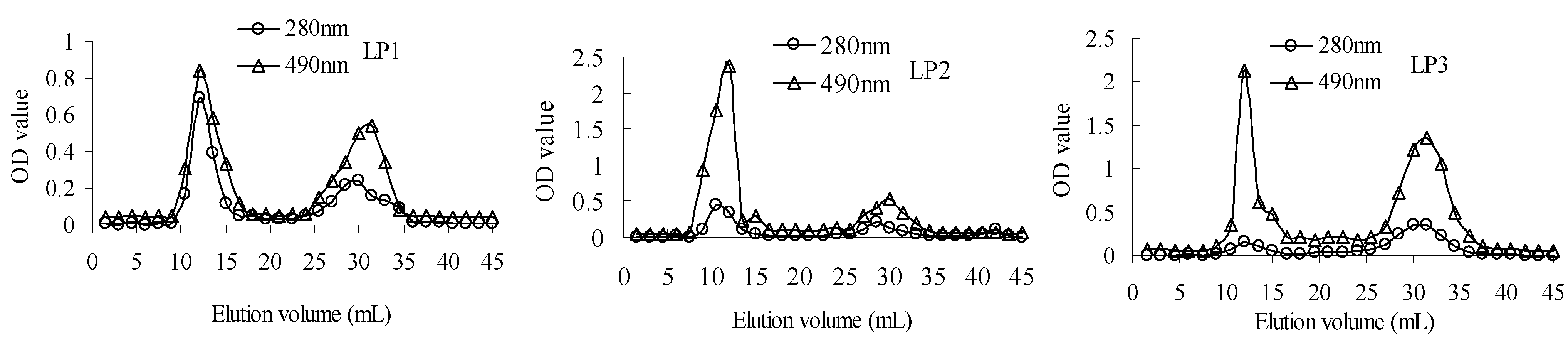{kind=link}
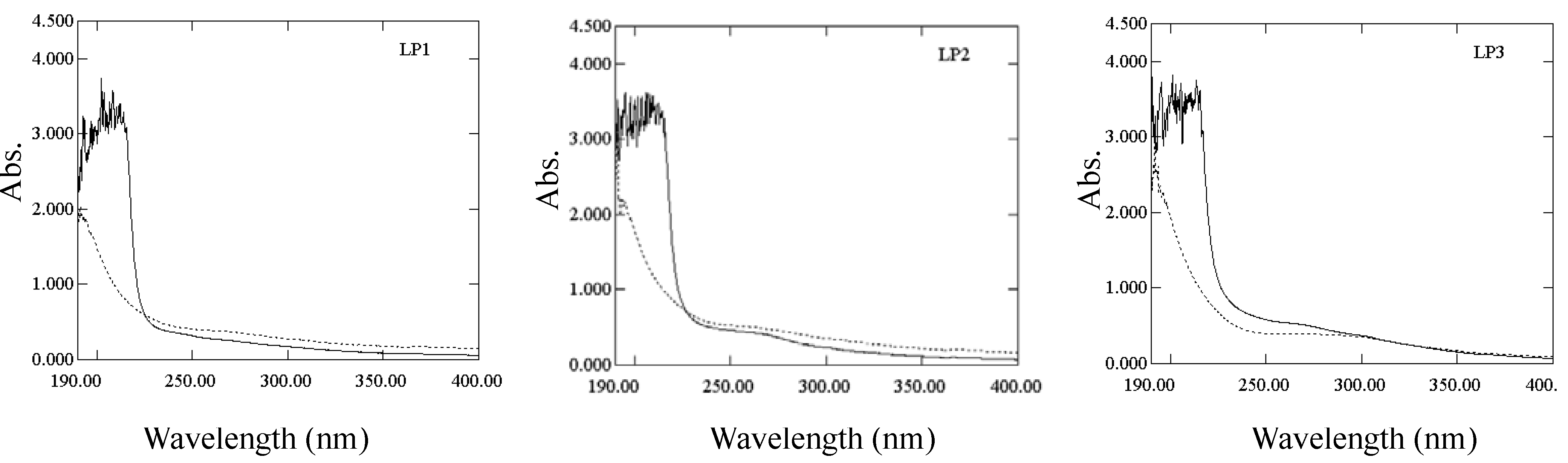{kind=link}
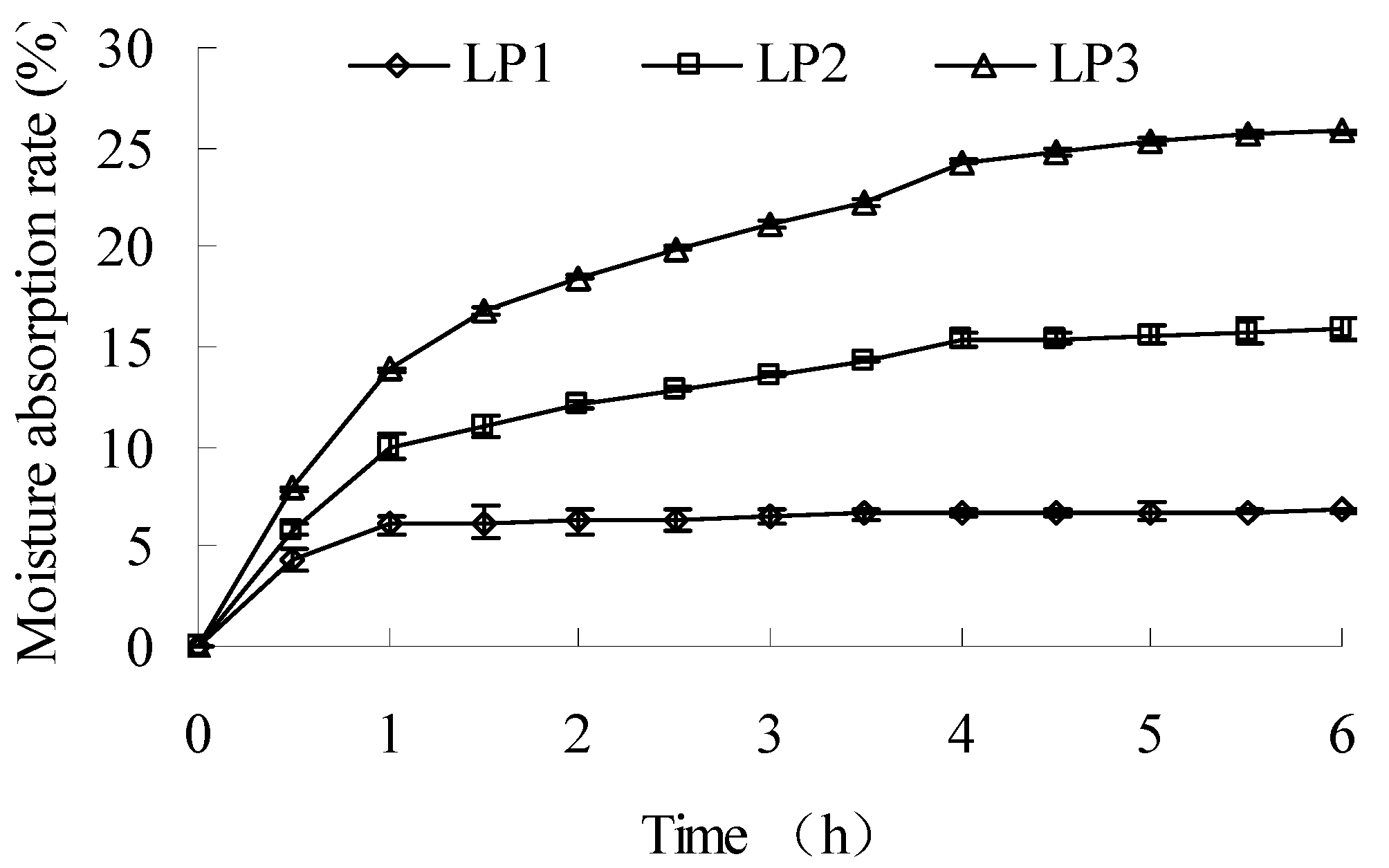{kind=link}
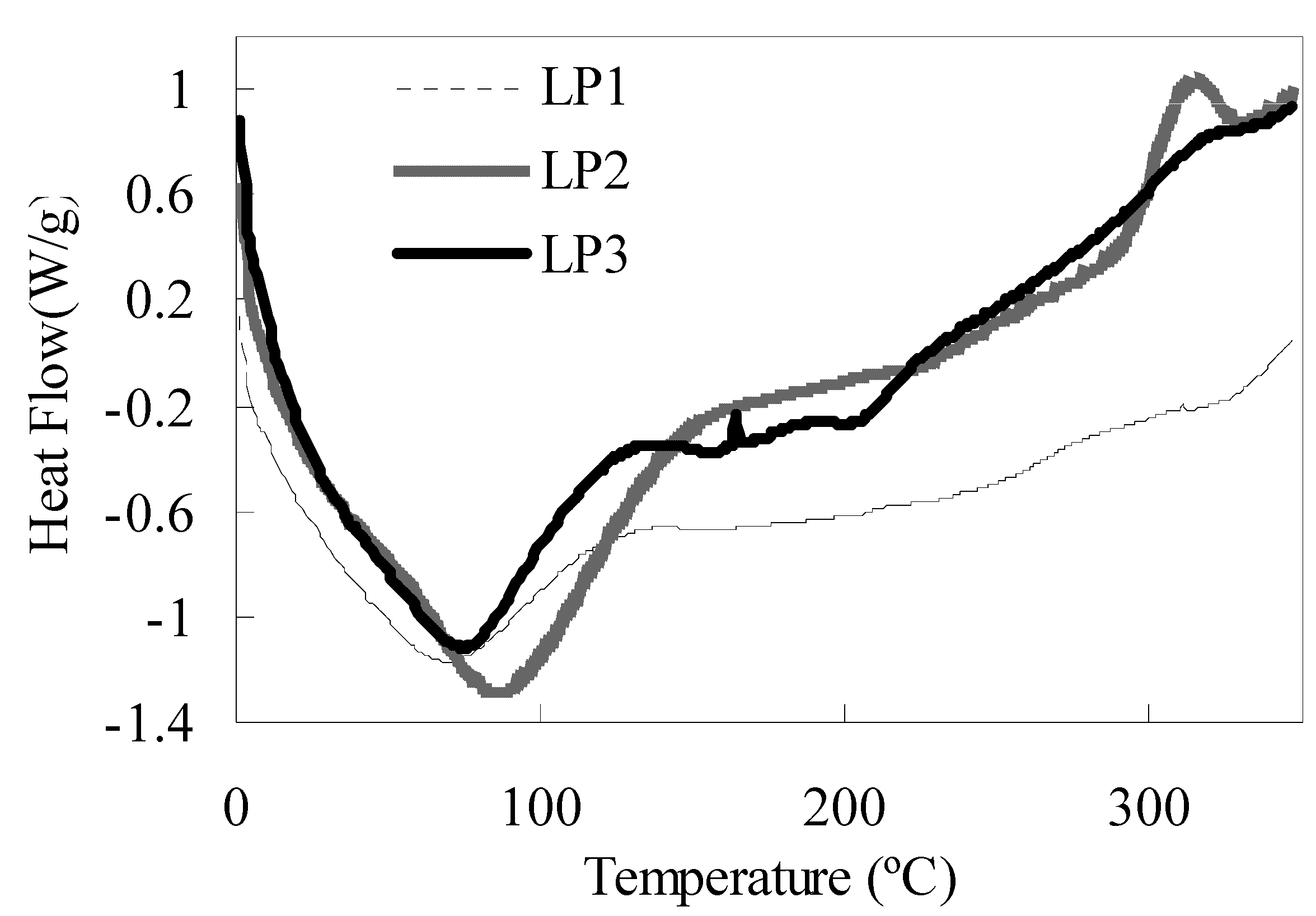{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | LP1 | LP2 | LP3 | Composition | LP1 | LP2 | LP3 |
|---|---|---|---|---|---|---|---|
| Neutral polysaccharide | 62.35 ± 2.12 a | 82.45 ± 2.42 b | 90.44 ± 2.12 c | Protein | 33.72 ± 0.16 c | 5.91 ± 0.07 b | 3.20 ± 0.06 a |
| Hexuronic acid | 1.98 ± 0.14 a | 9.03 ± 0.40 c | 4.31 ± 0.20 b | Aspartate | 3.90 | 0.73 | 0.31 |
| Percentage (%) | Glutamic | 4.34 | 0.96 | 1.00 | |||
| Ribose | 3.29 | 0.81 | 1.32 | Serine | 1.74 | 0.25 | 0.12 |
| Rhamnose | 2.91 | 0.40 | 0.12 | Glycine | 1.78 | 0.36 | 0.16 |
| Arabinose | 28.2 | 6.63 | 3.76 | Threonine | 1.86 | 0.29 | 0.12 |
| Xylose | 0.55 | 9.44 | 0.51 | Histidine | 0.86 | 0.10 | 0.05 |
| Mannose | 7.20 | 27.3 | 28.9 | Alanine * | 2.58 | 0.62 | 0.41 |
| Glucose | 46.8 | 54.6 | 63.9 | Arginine | 1.62 | 0.31 | 0.09 |
| Galactose | 11.0 | 0.87 | 1.50 | Tyrosine * | 1.22 | 0.09 | 0.02 |
| Molar ratio | Valine * | 2.49 | 0.41 | 0.16 | |||
| Ribose | 0.55 | 0.12 | 0.35 | Metione | 0.75 | 0.05 | <0.05 |
| Rhamnose | 0.44 | 0.06 | 0.03 | Phenylalanine * | 1.90 | 0.24 | 0.07 |
| Arabinose | 4.70 | 1.00 | 1.00 | Isoleucine * | 1.89 | 0.29 | 0.09 |
| Xylose | 0.09 | 1.42 | 0.14 | Leucine * | 3.34 | 0.49 | 0.20 |
| Mannose | 1.00 | 3.43 | 6.40 | Lysine | 1.98 | 0.46 | 0.16 |
| Glucose | 6.50 | 6.86 | 14.16 | Proline * | 1.48 | 0.28 | 0.19 |
| Galactose | 1.83 | 0.13 | 0.40 | ||||
2.1.3. Moisture absorption and thermodynamic property


2.1.4. FTIR spectrum characterization
| Absorption (cm−1) | Functional group | Structural characteristic | |||
|---|---|---|---|---|---|
| LP1 | LP2 | LP3 | |||
| 3398.0 | 3431.8 | 3430.9 | hydroxyl group (-OH) | stretching vibration of O-H | |
| amino group (-NH2) | stretching vibration of N-H | ||||
| 2924.6,2853.1 | 2928.0 | 2930.4 | alkyl group (-CH2-) | stretching vibration of C-H | |
| 1742.9 | carboxyl group (-COOH), aldehyde group (-CHO) or esterfunction (-COOR) | stretching vibration of C=O | |||
| 1654.5 | 1638.0 | 1647.4 | carbonyl group (-C=O or -CHO) | stretching vibration of C=O | |
| amide group (-NH2 or –COR) | bending vibration of N-H or stretching vibration of C=O | ||||
| amino group (-NH2) | bending vibration of N-H | ||||
| bound water | |||||
| 1541.3 | amino group (-NH2) or amide group (-NH2) | bending vibration of N-H | |||
| carbonyl group (-C=O) | stretching vibration of C=O | ||||
| 1457.9 | 1458.8 | 1458.0 | alkyl group (-CH2- or –CH3) | bending vibration of C-H | |
| 1418.8 | 1425.2 | carboxyl group (-COOH) | stretching vibration of C-O | ||
| 1378.2 | 1364.2 | 1363.6 | carboxyl group (-COOH) | symmetrical stretching vibration of C=O | |
| 1237.7 | 1275.1, 1209.1 | 1270.0 | carboxyl group (-COOH) | bending vibration of O-H | |
| 1159.1 | 1157.9 | ether (-C-O-C-) | stretching vibration of C-O | ||
| 1053.2 | 1043.9 | 1013.1 | hydroxyl group (-OH) | bending vibration of O-H | |
| 918.0 | 920.8 | D-glucopyranose ring | antisymmetrical ring vibration | ||
| 865.0 | furanose | ||||
| 847.9 | α-type glycosidic linkage | bending vibration of C-H | |||
| 819.6 | α-D-galactopyranose | ||||
| 764.2 | 776.2 | D-glucopyranose ring | symmetrical ring vibration | ||
2.2. Immunomodulatory activities of LP1-3
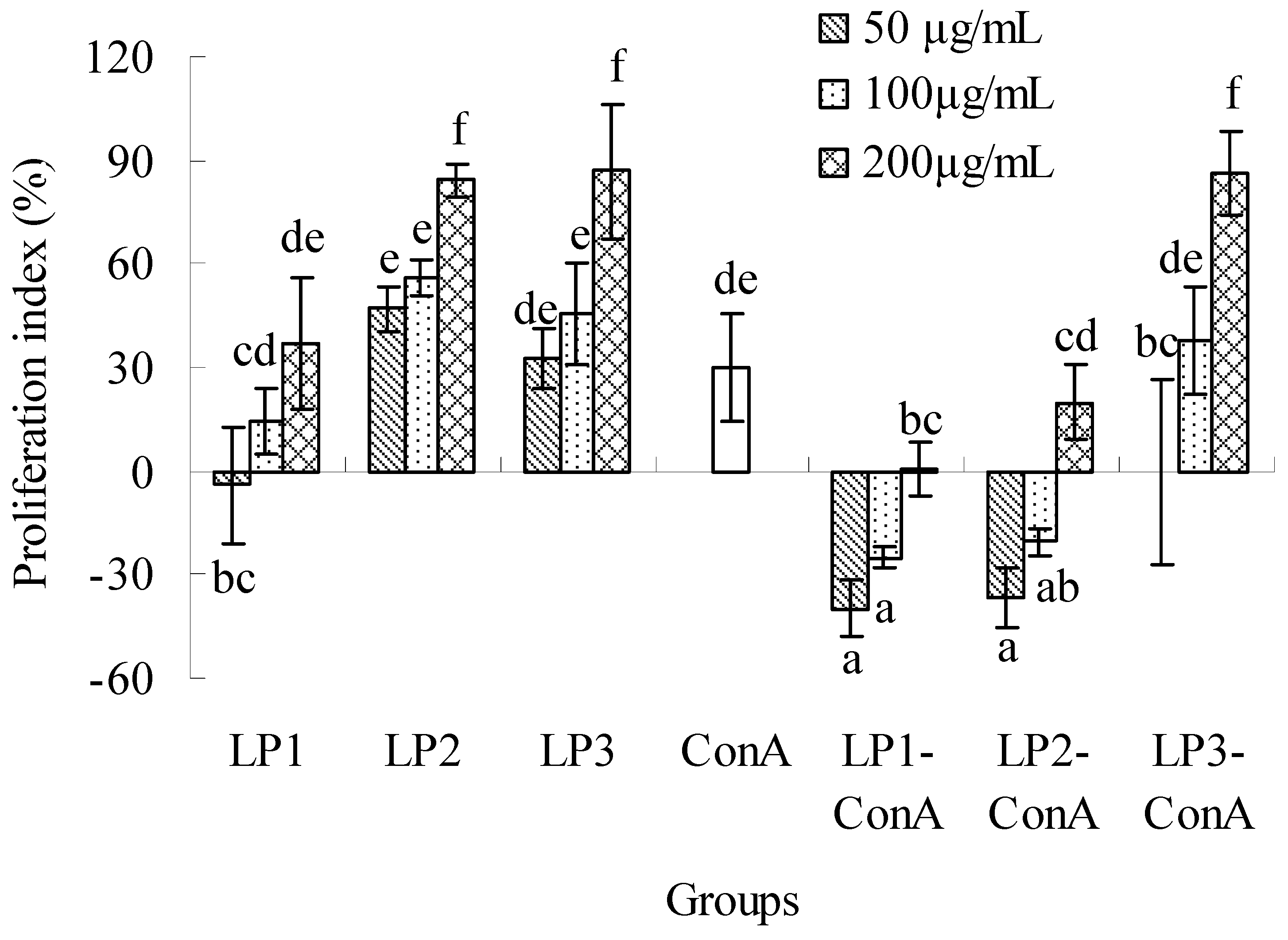
2.2.1. Effects on splenic lymphocyte proliferation

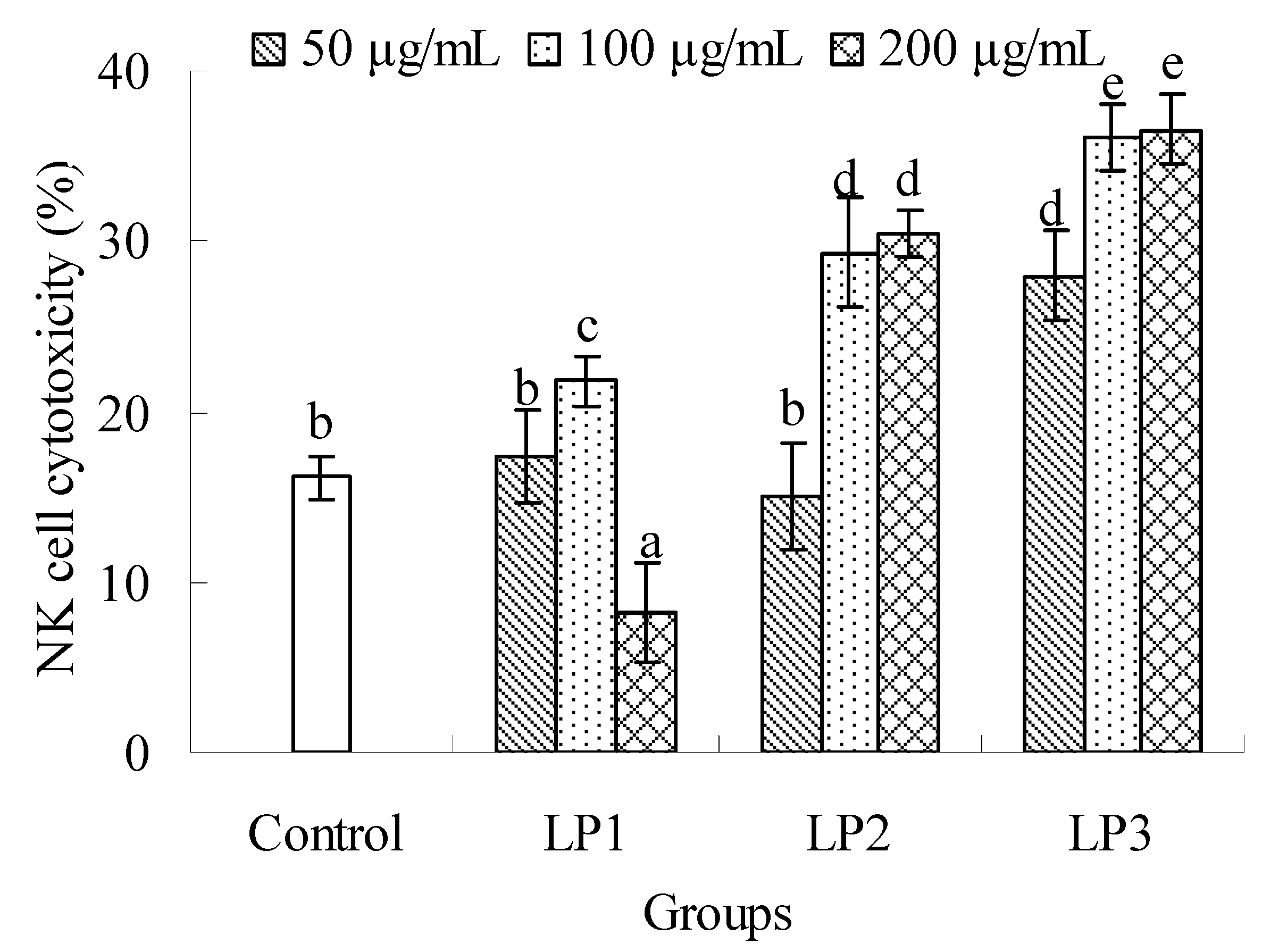
2.2.2. Effects on NK cell cytotoxicity

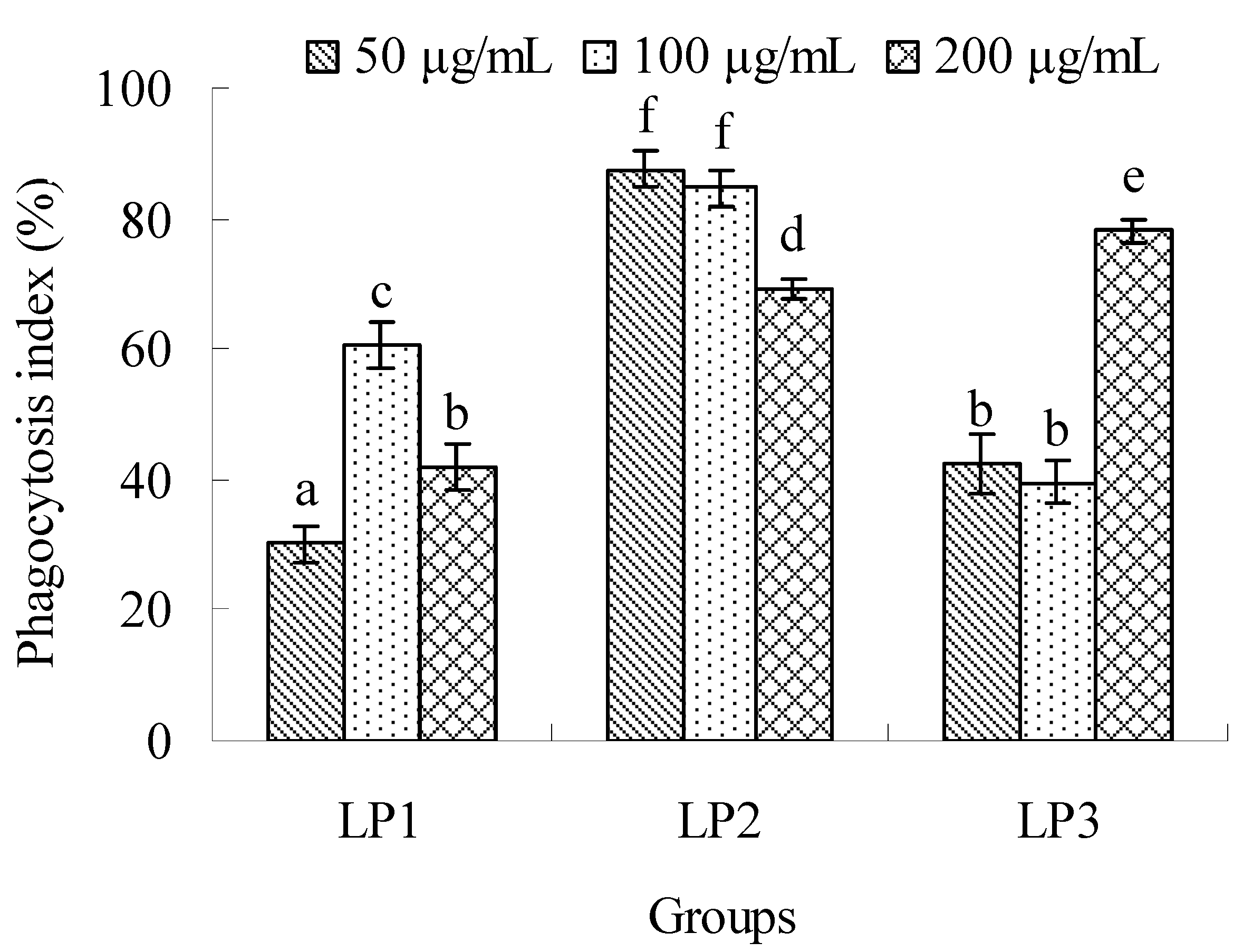
2.2.3. Effects on macrophage phagocytosis

3. Experimental
3.1. Plant materials
3.2. Preparation of LP
3.3. Characterization analysis
3.3.1. Characteristics of polysaccharide-protein complex
3.3.2. Chemical composition analysis
3.3.3. Moisture absorption
3.3.4. Differential scanning calorimetry (DSC)
3.3.5. Fourier transform infrared spectrum (FTIR)
3.4. Immunomodulatory activity analysis
3.4.1. Animals and cells
3.4.2. Splenic lymphocyte proliferation
3.4.3. Natural killer cell cytotoxicity
3.4.4. Macrophage phagocytosis
3.5. Statistical analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Gan, L.; Zhang, S.H.; Liu, Q.; Xu, H.B. A polysaccharide-protein complex from Lycium barbarum upregulates cytokine expression in human peripheral blood mononuclear cells. Eur. J. Pharmacol. 2003, 471, 217–222. [Google Scholar] [CrossRef]
- Liu, J.; Yang, F.; Ye, L.B.; Yang, X.J.; Timani, K.A.; Zheng, Y.; Wang, Y.H. Possible mode of action of antiherpetic activities of a proteoglycan isolated from the mycelia of Ganoderma lucidum in vitro. J. Ethnopharmacol. 2004, 95, 265–272. [Google Scholar] [CrossRef]
- Ye, L.B.; Zheng, X.L.; Zhang, J.S.; Tang, Q.Q.; Yang, Y.; Wang, X.Y.; Li, J.R.; Liu, Y.F.; Pan, Y.J. Biochemical characterization of a proteoglycan complex from an edible mushroom Ganoderma lucidum fruiting bodies and its immunoregulatory activity. Food Res. Int. 2011, 44, 367–372. [Google Scholar] [CrossRef]
- Zhang, J.S.; Tang, Q.J.; Zimmerman-Kordmann, M.; Reutter, W.; Fan, H. Activation of B lymphocytes by GLIS, a bioactive proteoglycan from Ganoderma lucidum. Life Sci. 2002, 71, 623–638. [Google Scholar] [CrossRef]
- Chen, Z.S.; Tan, B.K.H.; Chan, S.H. Activation of T lymphocytes by polysaccharide-protein complex from Lycium barbarum L . Int. Immunopharmacol. 2008, 8, 1663–1671. [Google Scholar] [CrossRef]
- Gan, L.; Zhang, S.H.; Yang, X.L.; Xu, H.B. Immunomodulation and antitumor activity by a polysaccharide-protein complex from Lycium barbarum. Int. Immunopharmacol. 2004, 4, 563–569. [Google Scholar] [CrossRef]
- Yang, J.; Wu, M.C.; Zhang, S.H.; Liang, G.Y. Study on the antifatigue effects of protein-bond polysaccharide from Lentinus edodes. Acta Nutrimenta Sin. 2001, 23, 350–353. [Google Scholar]
- Kim, G.Y.; Lee, M.Y.; Lee, H.J.; Moon, D.O.; Lee, C.M.; Jin, C.Y.; Choi, Y.H.; Jeong, Y.K.; Chung, K.T.; Lee, J.Y.; Choi, I.H.; Park, Y.M. Effect of water-soluble proteoglycan isolated from Agaricus blazei on the maturation of murine bone marrow-derived dendritic cells. Int. Immunopharmacol. 2005, 5, 1523–1532. [Google Scholar] [CrossRef]
- Park, S.J.; Park, D.H.; Kim, D.H.; Lee, S.; Yoon, B.H.; Jung, W.Y.; Lee, K.T.; Cheong, J.H.; Ryu, J.H. The memory-enhancing effects of Euphoria longan fruit extract in mice. J. Ethnopharmacol. 2010, 128, 160–165. [Google Scholar] [CrossRef]
- Yang, C.; He, N.; Ling, X.; Ye, M.; Zhang, C.; Shao, W.; Yao, C.; Wang, Z.; Li, Q. The isolation and characterization of polysaccharides from longan pulp. Sep. Purif. Technol. 2008, 63, 226–230. [Google Scholar] [CrossRef]
- Okuyama, E.; Ebihara, H.; Takeuchi, H.; Yamazaki, M. Adenosine, the anxiolytic-like principle of the Arillus of Euphoria longana. Planta Med. 1999, 65, 115–119. [Google Scholar]
- Su, D.X.; Zhang, M.W.; Liao, S.T.; Hou, F.L.; Zhang, R.L.; Tang, X.J.; Wei, Z.C.; Zhang, Y.; Chi, J.W.; Deng, Y.Y. Effects of water soluble extracts from longan on immune regulation in normal mice. Sci. Agric. Sin. 2010, 43, 1919–1925. [Google Scholar]
- Prasad, K.N.; Hao, J.; Shi, J.; Liu, T.; Li, J.; Wei, X.; Qiu, S.; Xue, S.; Jiang, Y. Antioxidant and anticancer activities of high pressure-assisted extract of longan (Dimocarpus longan Lour.) fruit pericarp. Innov. Food Sci. Emerg. 2009, 10, 413–419. [Google Scholar] [CrossRef]
- Yang, B.; Zhao, M.; Shi, J.; Yang, N.; Jiang, Y. Effect of ultrasonic treatment on the recovery and DPPH radical scavenging activity of polysaccharides from longan fruit pericarp. Food Chem. 2008, 106, 685–690. [Google Scholar] [CrossRef]
- Yang, B.; Zhao, M.; Jiang, Y. Optimization of tyrosinase inhibition activity of ultrasonic-extracted polysaccharides from longan fruit pericarp. Food Chem. 2008, 110, 294–300. [Google Scholar] [CrossRef]
- Yang, B.; Zhao, M.; Jiang, Y. Anti-glycated activity of polysaccharides of longan (Dimocarpus longan Lour.) fruit pericarp treated by ultrasonic wave. Food Chem. 2009, 114, 629–633. [Google Scholar] [CrossRef]
- Zhong, K.; Wang, Q.; He, Y.; He, X. Evaluation of radicals scavenging, immunity-modulatory and antitumor activities of longan polysaccharides with ultrasonic extraction on in S180 tumor mice models. Int. J. Biol. Macromol. 2010, 47, 356–360. [Google Scholar] [CrossRef]
- Zhu, C.P. Study on Structure and Bioactivities of Lycium Barbarum Polysaccharide, Huazhong Agricultural University, Wuhan, China, 2006.
- Yang, B.; Jiang, Y.; Zhao, M.; Chen, F.; Wang, R.; Chen, Y.; Zhang, D. Structural characterisation of polysaccharides purified from longan (Dimocarpus longan Lour.) fruit pericarp. Food Chem. 2009, 115, 609–614. [Google Scholar] [CrossRef]
- She, Z.G.; Hu, G.P.; Guo, Z.Y.; Lin, Y.C.; Lin, M.L. Study on the thermal decomposition of the sulphated polysaccharide Hal-A from Haliotis diverisicolor Reeve. Chin. J. Org. Chem. 2003, 23, 1149–1151. [Google Scholar]
- Zhang, W. Biochemical Technology of Carbohydrate Complexes; Zhejiang University Press: Hangzhou, China, 1994. [Google Scholar]
- Li, S.G.; Wang, D.G.; Tian, W.; Wang, X.X.; Zhao, J.X.; Liu, Z.; Chen, R. Characterization and anti-tumor activity of a polysaccharide from Hedysarum polybotrys Hand.-Mazz. Carbohydr. Polym. 2008, 73, 344–350. [Google Scholar] [CrossRef]
- Ruan, Z.; Su, J.; Dai, H.C.; Wu, M.C. Characterization and immunomodulating activities of polysaccharide from Lentinus edodes. Int. Immunopharmacol. 2005, 5, 811–820. [Google Scholar] [CrossRef]
- Shi, Y.; Sheng, J.; Yang, F.; Hu, Q. Purification and identification of polysaccharide derived from Chlorella pyrenoidosa. Food Chem. 2007, 103, 101–105. [Google Scholar] [CrossRef]
- Guo, L.; Xie, J.; Ruan, Y.; Zhou, L.; Zhu, H.; Yun, X.; Jiang, Y.; Lu, L.; Chen, K.; Min, Z.; Wen, Y.; Gu, J. Characterization and immunostimulatory activity of a polysaccharide from the spores of Ganoderma lucidum. Int. Immunopharmacol. 2009, 9, 1175–1182. [Google Scholar] [CrossRef]
- Capek, P.; Matulova, M.; Combourieu, B. The extracellular proteoglycan produced by Rhodella grisea. Int. J. Biol. Macromol. 2008, 43, 390–393. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, L. Characterization of polysaccharide-protein complexes by size-exclusion chromatography combined with three detectors. Carbohydr. Res. 2008, 343, 2251–2257. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, Y.; Zhang, L. Chemical modification and antitumor activities of two polysaccharide-protein complexes from Pleurotus tuber-regium. Int. J. Biol. Macromol. 2009, 45, 109–115. [Google Scholar] [CrossRef]
- Chen, C.F.; Yang, X.T.; Li, X.Q.; Mi, K.; Yang, Q.Y. A comparison study on the physical-chemical properties and immunomodulatory activities of mycelial polysaccharide extracts from five Ganoderma species. Acta Microbiol. Sin. 2007, 47, 628–633. [Google Scholar]
- Chen, H.S.; Tsai, Y.F.; Lin, S.; Lin, C.C.; Khoo, K.H.; Lin, C.H.; Wong, C.H. Studies on the immuno-modulating and anti-tumor activities of Ganoderma lucidum (Reishi) polysaccharides. Bioorg. Med. Chem. 2004, 12, 5595–5601. [Google Scholar] [CrossRef]
- Lin, K.I.; Kao, Y.Y.; Kuo, H.K.; Yang, W.B.; Chou, A.; Lin, H.H.; Yu, A.L.; Wong, C.H. Reishi polysaccharides induce immunoglobulin production through the TLR4/TLR2-mediated induction of transcription factor Blimp-1. J. Biol. Chem. 2006, 281, 24111–24123. [Google Scholar]
- Zhang, Y.; Kiyohara, H.; Matsumoto, T.; Yamada, H. Fractionation and chemical properties of immunomodulating polysaccharides from roots of Dipsacus asperoides. Planta Med. 1997, 63, 393–399. [Google Scholar] [CrossRef]
- Kralovec, J.A.; Metera, K.L.; Kumar, J.R.; Watson, L.V.; Girouard, G.S.; Guan, Y.; Carr, R.I.; Barrow, C.J.; Ewart, H.S. Immunostimulatory principles from Chlorella pyrenoidosa--part 1: isolation and biological assessment in vitro. Phytomedicine 2007, 14, 57–64. [Google Scholar] [CrossRef]
- Lo, T.C.T.; Jiang, Y.H.; Chao, A.L.J.; Chang, C.A. Use of statistical methods to find the polysaccharide structural characteristics and the relationships between monosaccharide composition ratio and macrophage stimulatory activity of regionally different strains of Lentinula edodes. Anal. Chim. Acta 2007, 584, 50–56. [Google Scholar]
- Leung, M.Y.K.; Liu, C.; Zhu, L.F.; Hui, Y.Z.; Yu, B.; Fung, K.P. Chemical and biological characterization of a polysaccharide biological response modifier from Aloe vera L. var. chinensis (Haw.) Berg. Glycobiology 2004, 14, 501–510. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, M.W.; Liao, S.T.; Tang, X.J.; Zhang, R.F.; Wei, Z.C. Optimization of ultrasonic-enzyme-assisted extraction technology of polysaccharide from longan pulp. Trans. Chin. Soc. Agric. Mach. 2010, 41, 131–136. [Google Scholar]
- Yi, Y.; Zhang, M.W.; Liao, S.T.; Tang, X.J.; Zhang, R.F.; Wei, Z.C. Optimization of de-coloration technology for polysaccharides from longan pulp. Trans. Chin. Soc. Agric. Mach. 2010, 41, 146–150, 155. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Pebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Li, X.; Jiao, L.L.; Zhang, X.; Tian, W.M.; Chen, S.; Zhang, L.P. Anti-tumor and immunomodulating activities of proteoglycans from mycelium of Phellinus nigricans and culture medium. Int. Immunopharmacol. 2008, 8, 909–915. [Google Scholar] [CrossRef]
- Wang, P.Y.; Wang, S.Z.; Lin, S.Q.; Lin, Z.B. Comparison of the immunomodulatory effects of spore polysaccharides and broken spore polysaccharides isolated from Ganoderma lucidum on murine splenic lymphocytes and peritoneal macrophages in vitro. J. Peking Univ. (Health Sci.) 2005, 36, 569–574. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yi, Y.; Liao, S.T.; Zhang, M.W.; Shi, J.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C. Physicochemical Characteristics and Immunomodulatory Activities of Three Polysaccharide-Protein Complexes of Longan Pulp. Molecules 2011, 16, 6148-6164. https://doi.org/10.3390/molecules16076148
Yi Y, Liao ST, Zhang MW, Shi J, Zhang RF, Deng YY, Wei ZC. Physicochemical Characteristics and Immunomodulatory Activities of Three Polysaccharide-Protein Complexes of Longan Pulp. Molecules. 2011; 16(7):6148-6164. https://doi.org/10.3390/molecules16076148
Chicago/Turabian StyleYi, Yang, Sen Tai Liao, Ming Wei Zhang, John Shi, Rui Fen Zhang, Yuan Yuan Deng, and Zhen Cheng Wei. 2011. "Physicochemical Characteristics and Immunomodulatory Activities of Three Polysaccharide-Protein Complexes of Longan Pulp" Molecules 16, no. 7: 6148-6164. https://doi.org/10.3390/molecules16076148
APA StyleYi, Y., Liao, S. T., Zhang, M. W., Shi, J., Zhang, R. F., Deng, Y. Y., & Wei, Z. C. (2011). Physicochemical Characteristics and Immunomodulatory Activities of Three Polysaccharide-Protein Complexes of Longan Pulp. Molecules, 16(7), 6148-6164. https://doi.org/10.3390/molecules16076148