Antiviral Properties of Lactoferrin—A Natural Immunity Molecule


 and
and 
Abstract
:1. Introduction

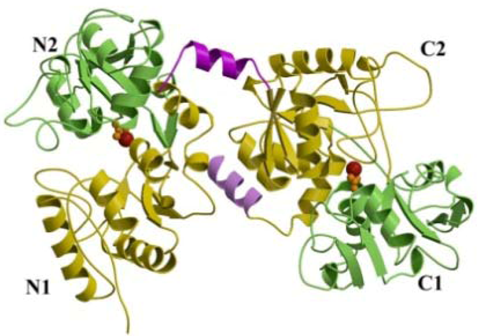
2. Structure
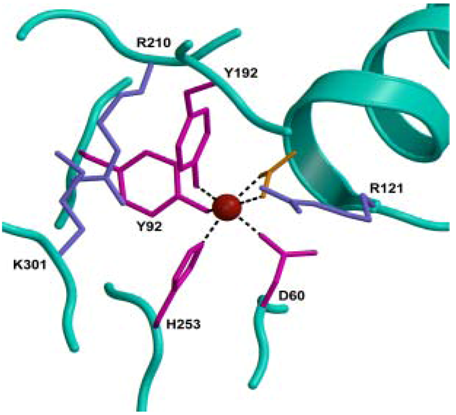
2.1. Iron-Binding Sites

2.2. Conformational Changes
2.3. Binding of other Metals
2.4. Glycosylation
3. Human and Bovine Lactoferrin Gene Structure and Regulation
4. Concentrations in Human Body

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological fluids | Concentration (mg/mL) |
|---|---|
| Colostrum | 8 |
| Milk | 1.5–4 |
| Tears | 2 |
| Saliva | 0.008 |
| Joint fluid | 0.001 |
| Vaginal secretion | 0.008 |
| Seminal fluid | 0.112 |
| Cerebrospinal fluid | Undetectable |
| Plasma | 0.0004 |
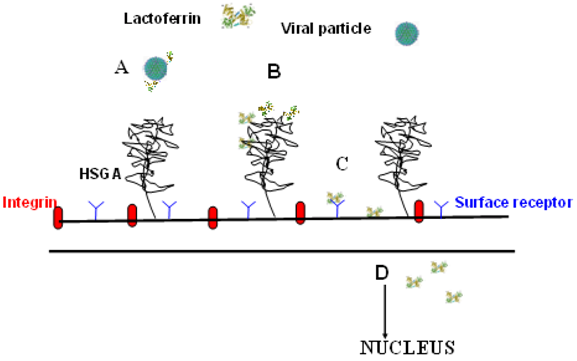
5. Lactoferrin as Human Innate Defence against Infections

7. Inhibition of Viral Infections by Lactoferrin
7.1. Herpesvirus
7.2. Human Immunodeficiency Virus (HIV)
7.3. Friend Virus Complex (FVC)
7.4. Human Hepatitis C Virus (HCV)
7.5. Human Hepatitis B Virus (HBV)
7.6. Respiratory Syncytial Virus (RSV) and Parainfluenza Virus (PIV)
7.7. Alphavirus
7.8. Hantavirus
7.9. Human Papillomavirus (HPV)
7.10. Rotavirus
7.11. Feline Calicivirus (FCV)
7.12. Adenovirus
7.14. Rhinovirus
7.15. Influenza A Virus
7.16. Japanese Encephalitis Virus
7.17. Tomato Yellow Leaf Curl Virus
8. Conclusions

Acknowledgments
References and Notes
- Soerensen, M.; Soerensen, S.P.L. The proteins in whey. C. R. Trav. Lab. Carlsberg. 1939, 23, 55–99. [Google Scholar]
- Johansson, B. Isolation of an iron-containing red protein from human milk. Acta Chem. Scand. 1960, 14, 510–512. [Google Scholar] [CrossRef]
- Montreuil, J.; Tonnelat, J.; Mullet, S. Preparation and properties of lactosiderophilin (lactotransferrin) of human milk. Biochim. Biophys. Acta 1960, 45, 413–421. [Google Scholar] [CrossRef]
- Groves, M.L. The isolation of a red protein from milk. J. Am. Chem. Soc. 1960, 82, 3345–3350. [Google Scholar] [CrossRef]
- Mazurier, J.; Spik, G. Comparative study of the iron-binding properties of human transferrins. I. Complete and sequential iron saturation and desaturation of the lactotransferrin. Biochim. Biophys. Acta 1980, 629, 399–408. [Google Scholar] [CrossRef]
- Baker, E.N. Structure and reactivity of transferrins. Adv. Inorg. Chem. 1994, 41, 389–463. [Google Scholar] [CrossRef]
- Anderson, B.F.; Baker, H.M.; Dodson, E.J.; Norris, G.E.; Rumball, S.V.; Waters, J.M.; Baker, E.N. Structure of human lactoferrin at 3.2-A resolution. Proc. Natl. Acad. Sci. USA 1987, 84, 1769–1773. [Google Scholar]
- Anderson, B.F.; Baker, H.M.; Norris, G.E.; Rice, D.W.; Baker, E.N. Structure of human lactoferrin: Crystallographic structure analysis and refinement at 2.8 A resolution. J. Mol. Biol. 1989, 209, 711–734. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M. Molecular structure, binding properties and dynamics of lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2531–2539. [Google Scholar] [CrossRef]
- Metz-Boutigue, M.-H.; Jolles, J.; Mazurier, J.; Schoentgen, F.; Legran, D.D.; Spik, G.; Montreuil, J.; Jolles, P. Human lactotransferrin: Amino acid sequence and structural comparisons with othertransferrins. Eur. J. Biochem. 1984, 145, 659–676. [Google Scholar] [CrossRef]
- Baker, E.N.; Rumball, S.V.; Anderson, B.F. Transferrins: Insights into structure and function from studies on lactoferrin. Trends Biochem. Sci. 1987, 12, 350–353. [Google Scholar] [CrossRef]
- Bruns, C.M.; Nowalk, A.J.; Arvai, A.S.; McTigue, M.A.; Vaughan, K.G.; Mietzner, T.A.; McRee, D.E. Structure of Haemophilus influenzae Fe(+3)-binding protein reveals convergent evolution within a superfamily. Nat. Struct. Biol. 1997, 4, 919–924. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M.; Kidd, R.D. Lactoferrin and transferrin: Functional variations on a common structural framework. Biochem. Cell Biol. 2002, 80, 27–34. [Google Scholar] [CrossRef]
- MacGillivray, R.T.A.; Moore, S.A.; Chen, J.; Anderson, B.F.; Baker, H.; Luo, Y.; Bewley, M.; Smith, C.A.; Murphy, M.E.P.; Wang, Y.; Mason, A.B.; Woodworth, R.C.; et al. Two high-resolution crystal structures of the recombinant N-lobe of human transferrin reveal a structural change implicated in iron release. Biochemistry 1998, 37, 7919–7928. [Google Scholar]
- Grossmann, J.G.; Neu, M.; Pantos, E.; Schwab, F.J.; Evans, R.W.; Towns-Andrews, E.; Lindley, P.F.; Appel, H.; Thies, W.G.; Hasnain, S.S. X-ray solution scattering reveals conformational changes upon iron uptake in lactoferrin, serum and ovo-transferrins. J. Mol. Biol. 1992, 225, 811–819. [Google Scholar] [CrossRef]
- Octave, J.N.; Schneider, Y.J.; Trouet, A.; Crichton, R.R. Iron uptake and utilization by mammalian cells. I: Cellular uptake of transferrin and iron. Trends Biochem. Sci. 1983, 8, 217–220. [Google Scholar] [CrossRef]
- Day, C.L.; Stowell, K.M.; Baker, E.N.; Tweedie, J.W. Studies of the N-terminal half of human lactoferrin produced from the cloned cDNA demonstrate that interlobe interactions modulate iron release. J. Biol. Chem. 1992, 267, 13857–13862. [Google Scholar]
- Ward, P.P.; Zhou, X.; Conneely, O.M. Cooperative interactions between the amino- and carboxyl38 terminal lobes contribute to the unique iron-binding stability of lactoferrin. J. Biol. Chem. 1996, 271, 12790–12794. [Google Scholar]
- Jameson, G.B.; Anderson, B.F.; Norris, G.E.; Thomas, D.H.; Baker, E.N. Structure of human apolactoferrin at 2.0 A resolution. Refinement and analysis of ligand-induced conformational change. Acta Crystallogr. D. Biol. Crystallogr. 1998, 54, 1319–1335. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence againstmicrobial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef]
- Kim, S.J.; Yu, D.Y.; Pak, K.W.; Jeong, S.; Kim, S.W.; Lee, K.K. Structure of the human lactoferrin gene and its chromosomal localization. Mol. Cells 1998, 8, 663–668. [Google Scholar]
- Schwerin, M.; Solinas Toldo, S.; Eggen, A.; Brunner, R.; Seyfert, H.M.; Fries, R. The bovine lactoferrin gene (LTF) maps to chromosome 22 and syntenic group U12. Mamm. Genome 1994, 5, 486–489. [Google Scholar] [CrossRef]
- Johnston, J.J.; Rintels, P.; Chung, J.; Sather, J.; Benz, E.J., Jr.; Berliner, N. Lactoferrin gene promoter: Structural integrity and nonexpression in HL60 cells. Blood 1992, 79, 2998–3006. [Google Scholar]
- Panella, T.J.; Liu, Y.H.; Huang, A.T.; Teng, C.T. Polymorphism and altered methylation of the lactoferrin gene in normal leukocytes, leukemic cells, and breast cancer. Cancer Res. 1991, 51, 3037–3043. [Google Scholar]
- Rado, T.A.; Wei, X.P.; Benz, E.J. Isolation of lactoferrin cDNA from a human myeloid library and expression of mRNA during normal and leukemic myelopoiesis. Blood 1987, 70, 989–993. [Google Scholar]
- Seyfert, H.M.; Kuhn, C. Characterization of a first bovine lactoferrin gene variant, based on an EcoRI polymorphism. Anim. Genet. 1994, 25, 54. [Google Scholar]
- Teng, C.T. Regulation of lactoferrin gene expression by estrogen and epidermal growth factor: Molecular mechanism. Cell Biochem. Biophys. 1999, 31, 49–64. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F. Lactoferrin in milk from different species. Comp. Biochem. Physiol. 1971, B39, 119–129. [Google Scholar]
- Masson, P.L.; Heremans, J.F.; Dive, C. An iron-binding protein common to many external secretions. Clin. Chim. Acta 1966, 14, 735–739. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F.; Schonne, E. Lactoferrin, an iron-binding protein in neutrophilic leukocytes. J. Exp. Med. 1969, 130, 643–658. [Google Scholar] [CrossRef]
- Dalmastri, C.; Valenti, P.; Visca, P.; Vittorioso, P.; Orsi, N. Enhanced antimicrobial activity of lactoferrin by binding to the bacterial surface. Microbiologica 1988, 11, 225–230. [Google Scholar]
- Naidu, A.S.; Anderson, M.; Miedzobrodzki, J.; Forsgren, A. Bovine lactoferrin receptors in Staphylococcus aureus isolated from bovine mastitis. J. Dairy Sci. 1991, 74, 1218–1226. [Google Scholar] [CrossRef]
- Marchetti, M.; Superti, F. Antiviral activity of lactoferrin. In Recent Developments in Antiviral Research; Pandalai S.G., Ed.; Transworld Research Network: Trivandrum, India, 2001; pp. 193–203. [Google Scholar]
- He, J.; Furmanski, P. Sequence specificity and transcriptional activation in the binding of lactoferrin to DNA. Nature 1995, 373, 721–724. [Google Scholar] [CrossRef]
- van Berkel, P.H.C.; Geerts, M.E.J.; van Veen, H.A.; Mericskay, M.; de Boer, H.A.; Nuijens, J.H. The N-terminal stretch Arg2 Arg3, Arg4, and Arg5 of human lactoferrin is essential for binding to heparin, bacterial lipopolysaccharide, human lysozyme and DNA. Biochem. J. 1997, 328, 145–151. [Google Scholar]
- Mann, D.M.; Romm, E.; Migliorini, M. Delineation of the glycosaminoglycan-binding site in the human inflammatory response protein lactoferrin. J. Biol. Chem. 1994, 269, 23661–23667. [Google Scholar]
- Schaible, U.E.; Kaufmann, H.E. Iron and microbial infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef]
- Orsi, N. The antimicrobial activity of lactoferrin: Current status and perspectives. Biometals 2004, 17, 189–196. [Google Scholar] [CrossRef]
- Bullen, J.J.; Rogers, H.J.; Leigh, L. Iron-binding proteins in milk and resistance to Escherichia coli infections in infants. Br. Med. J. 1972, 1, 69–75. [Google Scholar]
- Weinberg, E.D. The development of awareness of iron-withholding defense. Perspect. Biol. Med. 1993, 36, 215–221. [Google Scholar]
- Ratledge, C.; Dover, L.G. Iron metabolism in pathogenic bacteria. Annu. Rev. Microbiol. 2000, 54, 881–941. [Google Scholar] [CrossRef]
- Braun, V. Bacterial iron transport related to virulence. Contrib. Microbiol. 2005, 12, 210–233. [Google Scholar] [CrossRef]
- Ekins, A.; Khan, A.G.; Shouldice, S.R.; Schryvers, A.B. Lactoferrin receptors in Gram-negative bacteria: Insights into the iron acquisition process. Biometals 2004, 17, 235–243. [Google Scholar] [CrossRef]
- Arnold, R.R.; Cole, M.F.; Mcghee, J.R. A bactericidal effect for human lactoferrin. Science 1977, 197, 263–265. [Google Scholar]
- Ellison, R.T., III; Giehl, T.J.; La Force, F.M. Damage of the outer membrane of enteric gram negative bacteria by lactoferrin and transferrin. Infect. Immun. 1988, 56, 2774–2781. [Google Scholar]
- Ellison, R.T., III; La Force, M.; Giehl, T.J.; Boose, D.; Dunn, B.E. Lactoferrin and transferring damage of the Gramnegative outer membrane is modulated by Ca++ and Mg++. J. Gen. Microbiol. 1990, 136, 1437–1446. [Google Scholar] [CrossRef]
- Visca, P.; Dalmastri, C.; Verzili, D.; Antonini, G.; Chiancone, E.; Valenti, P. Interaction of lactoferrin with Escherichia coli cells and correlation with antibacterial activity. Med. Microbiol.Immunol. 1990, 179, 323–333. [Google Scholar]
- Appelmelk, B.J.; An, Y.Q.; Geerts, M.; Thijs, B.G.; de Boer, H.A.; MacLaren, D.M.; de Graaff, J.; Nuijens, J.H. Lactoferrin is a lipid A-binding protein. Infect. Immun. 1994, 62, 2628–2632. [Google Scholar]
- Brandenburg, K.; Jurgens, G.; Muller, M.; Fukuoka, S.; Koch, M.H.J. Biophysical characterization of lipopolysaccharide and lipid A inactivation by lactoferrin. Biol. Chem. 2001, 382, 1215–1225. [Google Scholar]
- Tomita, M.; Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, K.; Kavase, K. Potent antibacterial peptides generated by pepsin digestion of bovine lactoferrin. J. Dairy Sci. 1991, 74, 4137–4142. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta 1992, 1121, 130–136. [Google Scholar]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kavase, K.; Tomita, M. Antibacterial spectrum of lactoferricin B, a potent bactericide peptide derived from the N-terminal region of bovine lactoferrin. J. Appl. Bacteriol. 1992, 73, 472–479. [Google Scholar] [CrossRef]
- Rossi, P.; Giansanti, F.; Boffi, A.; Ajello, M.; Valenti, P.; Chiancone, E.; Antonini, G. Ca2+ binding to bovine lactoferrin enhances protein stability and infl uences the release of bacterial lipopolysaccharide. Biochem. Cell Biol. 2002, 80, 41–48. [Google Scholar] [CrossRef]
- Lu, L.; Hangoc, G.; Oliff, A.; Chen, L.T.; Shen, R.N.; Broxmeyer, H.E. Protective influence of lactoferrin on mice infected with the polycythemia-inducing strain of Friend virus complex. Cancer Res. 1987, 47, 4184–4188. [Google Scholar]
- Hasegawa, K.; Motsuchi, W.; Tanaka, S.; Dosako, S. Inhibition with lactoferrin of in vitro infection with human herpes virus. Jpn. J. Med. Sci. Biol. 1994, 47, 73–85. [Google Scholar]
- Superti, F.; Berlutti, F.; Paesano, R.; Valenti, P. Structure and activity of lactoferrin—A multi functional protective agent for human health. In Iron Metabolism and Disease; Fuchs, H., Ed.; Research Signpost: Kerala, India, 2008; pp. 1–32. [Google Scholar]
- Andersen, J.H.; Osbakk, S.A.; Vorland, L.H.; Traavik, T.; Gutteberg, T.J. Lactoferrin and cyclic lactoferricin inhibit the entry of human cytomegalovirus into human fibroblasts. Antiviral Res. 2001, 51, 141–149. [Google Scholar] [CrossRef]
- Harmsen, M.C.; Swart, P.J.; de Bethune, M.P.; Pauwels, R.; De Clercq, E.; The, T.H.; Meijer, D.K. Antiviral effects of plasma and milk proteins: Lactoferrin shows a potent activity against both human immunodeficiency virus and human cytomegalovirus replication in vitro. J. Infect. Dis. 1995, 172, 380–388. [Google Scholar] [CrossRef]
- Beljaars, L.; van der Strate, B.W.; Bakker, H.I.; Reker-Smit, C.; van Loenen-Weemaes, A.M.; Wiegmans, F.C.; Harmsen, M.C.; Molema, G.; Meijer, D.K. Inhibition of cytomegalovirus infection by lactoferrin in vitro and in vivo. Antiviral Res. 2004, 63, 197–208. [Google Scholar] [CrossRef]
- Marchetti, M.; Longhi, C.; Conte, M.P.; Pisani, S.; Valenti, P.; Seganti, L. Lactoferrin inhibits herpes simplex virus type 1 adsorption to Vero cells. Antiviral Res. 1996, 29, 221–231. [Google Scholar] [CrossRef]
- Marchetti, M.; Pisani, S.; Antonini, G.; Valenti, P.; Seganti, L.; Orsi, N. Metal complexes of bovine lactoferrin inhibit in vitro replication of herpes simplex virus type 1 and 2. Biometals 1998, 11, 89–94. [Google Scholar] [CrossRef]
- Siciliano, R.; Rega, B.; Marchetti, M.; Seganti, L.; Antonini, G.; Valenti, P. Bovine lactoferrin peptidic fragments involved in inhibition of herpes simplex virus type-1 infection. Biochem. Biophys. Res. Commun. 1999, 264, 19–23. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G.; Siciliano, R.; Rega, B.; Superti, F.; Marchetti, M.; Ammendolia, M.G.; Seganti, L. Antiviral activity of lactoferrin-derived peptides. In Lactoferrin: Structure, Function, and Applications; Shimazaki, K., Tsuda, H., Tomita, M., Kuwata, T., Perraudin, J.P., Eds.; Elsevier Science Ltd: Tokyo, Japan, 2000; pp. 181–186. [Google Scholar]
- Seganti, L.; Di Biase, A.M.; Rega, B.; De Giulio, B.; Nicoletti, M.; Antonini, G.; Valenti, P. Involvement of bovine lactoferrin moieties in the inhibition of herpes simplex virus type 1 infection. Int. J. Immunopathol. Pharmacol. 2001, 14, 71–79. [Google Scholar]
- Shukla, D.; Spear, P.G. Herpesviruses and heparan sulfate: An intimate relationship in aid of viral entry. J. Clin. Invest. 2001, 108, 503–510. [Google Scholar]
- Marchetti, M.; Trybala, E.; Superti, F.; Johansson, M.; Bergstrom, T. Inhibition of herpes simplex virus infection by lactoferrin is dependent on interference with the virus binding to glycosaminoglycans. Virology 2004, 318, 405–413. [Google Scholar] [CrossRef]
- Marchetti, M.; Ammendolia, M.G.; Superti, F. Glycosaminoglycans are not indispensabile for the anti-herpes simplex virus type 2 activity of lactoferrin. Biochemistry 2009, 91, 155–159. [Google Scholar] [CrossRef]
- Välimaa, H.; Tenovuo, J.; Waris, M.; Hukkanen, V. Human lactoferrin but not lysozyme neutralizes HSV-1 and inhibits HSV-1 replication and cell-to-cell spread. J. Virol. 2009, 6, 53. [Google Scholar] [CrossRef]
- Andersen, J.H.; Jenssen, H.; Gutteberg, T.J. Lactoferrin and lactoferricin inhibit Herpes simplex 1 and 2 infection and exhibit synergy when combined with acyclovir. Antiviral Res. 2003, 58, 209–215. [Google Scholar] [CrossRef]
- Anderssen, J.H.; Jenssen, H.; Sandvik, K.; Gutteberg, T.J. Anti-HSV activity of lactoferrin and lactoferricin is dependent on the presence of heparan sulphate at the cell surface. J. Med. Virol. 2004, 74, 262–271. [Google Scholar] [CrossRef]
- Jenssen, H.; Anderssen, J.H.; Uhlin-Hanssen, L.; Gutteberg, T.J.; Rekdal, O. Anti-HSV activity of lactoferricin analogues is only partly related to their affinity for heparan sulfate. Antiviral Res. 2004, 61, 101–109. [Google Scholar] [CrossRef]
- Jenssen, H.; Sandvik, K.; Andersen, J.H.; Hancock, R.E.W.; Gutteberg, T.J. Inhibition of HSV cell-to-cell spread by lactoferrin and lactoferricin. Antiviral Res. 2008, 79, 192–198. [Google Scholar] [CrossRef]
- Ammendolia, M.G.; Pietrantoni, A.; Tinari, A.; Valenti, P.; Superti, F. Bovine lactoferrin inhibits echovirus endocytic pathway by interacting with viral structural polypeptides. Antiviral Res. 2007, 73, 151–160. [Google Scholar] [CrossRef]
- Marr, A.K.; Jenssen, H.; Moniri, M.R.; Hancock, R.E.W.; Panté, N. Bovine lactoferrin and lactoferricin interfere with intracellular trafficking of Herpes simplex virus-1. Biochemistry 2009, 91, 160–164. [Google Scholar] [CrossRef]
- Shimizu, K.; Matsuzawa, H.; Okada, K.; Tazume, S.; Dosako, S.; Kawasaki, Y.; Hashimoto, K.; Koga, Y. Lactoferrin-mediated protection of the host from murine cytomegalovirus infection by a T-cell dependent augmentation of natural killer cell activity. Arch. Virol. 1996, 141, 1875–1889. [Google Scholar] [CrossRef]
- Fujihara, T.; Hayashi, K. Lactoferrin inhibits herpes simplex virus type-1 (HSV-1) infection in mouse cornea. Arch. Virol. 1995, 140, 1469–1472. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Kurokawa, M.; Shin, K.; Teraguchi, S.; Tamura, Y.; Shiraki, K. Oral lactoferrin prevents body weight loss and increases cytokine responses during herpes simplex virus type 1 infection of mice. Biosci. Biotechnol. Biochem. 2004, 68, 537–544. [Google Scholar] [CrossRef]
- Shust, G.F.; Cho, S.; Kim, M.; Madan, R.P.; Guzman, E.M.; Pollack, M.; Epstein, J.; Cohen, H.W.; Keller, M.J.; Herold, B.C. Female genital tract secretions inhibit herpes simplex virus infection: Correlation with soluble mucosal immune mediators and impact of hormonal contraception. Am. J. Reprod. Immunol. 2010, 63, 110–119. [Google Scholar]
- Beaumont, S.L.; Maggs, D.J.; Clarke, H.E. Effects of bovine lactoferrin on in vitro replication of feline herpesvirus. Vet. Ophthalmol. 2003, 6, 245–250. [Google Scholar] [CrossRef]
- Tanaka, T.; Nakatani, S.; Xuan, X.; Kumura, H.; Igarashi, I.; Shimazaki, K. Antiviral activity of lactoferrin against canine herpesvirus. Antiviral Res. 2003, 60, 193–199. [Google Scholar] [CrossRef]
- Swart, P.J.; Harmsen, M.C.; Kuipers, M.E.; Van Dijk, A.A.; Van Der Strate, B.W.; Van Berkel, P.H.; Nuijens, J.H.; Smit, C.; Witvrouw, M.; De Clercq, E.; et al. Charge modification of plasma and milk proteins results in antiviral active compounds. J. Pept. Sci. 1999, 5, 563–576. [Google Scholar] [CrossRef]
- Swart, P.J.; Kuipers, M.E.; Smith, C.; Pawels, R.; De Bethune, M.P.; De Clerck, E.; Meijer, D.K.; Huisman, J.G. Antiviral effects of milk proteins: Acylation results in polyanionic compounds with potent activity against human immunodeficiency virus types 1 and 2 in vitro. AIDS Res. Hum. Retroviruses 1996, 12, 769–775. [Google Scholar] [CrossRef]
- Puddu, P.; Borghi, P.; Gessani, S.; Valenti, P.; Belardelli, F.; Seganti, L. Antiviral effect of bovine lactoferrin saturated with metal ions on early steps of human immunodeficiency virus type 1 infection. Int. J. Biochem. Cell Biol. 1998, 30, 1055–1062. [Google Scholar] [CrossRef]
- Ng, T.B.; Lam, T.L.; Au, T.K.; Ye, X.Y.; Wan, C.C. Inhibition of human immunodeficiency virus type 1 reverse transcriptase, protease and integrase by bovine milk proteins. Life Sci. 2001, 69, 2217–2223. [Google Scholar] [CrossRef]
- Berkhout, B.; van Wamel, J.L.; Beljaars, L.; Meijer, D.K.; Visser, S.; Floris, R. Characterization of the anti-HIV effects of native lactoferrin and other milk proteins and protein-derived peptides. Antiviral Res. 2002, 55, 341–355. [Google Scholar] [CrossRef]
- Groot, F.; Geijtenbeek, T.B.; Sanders, R.W.; Baldwin, C.E.; Sanchez-Hernandez, M.; Floris, R.; van Kooyk, Y.; de JonG, E.C.; Berkhout, B. Lactoferrin prevents dendritic cell-mediated human immunodeficiency virus type 1 transmission by blocking the DC-SIGN—gp120 interaction. J. Virol. 2005, 79, 3009–3015. [Google Scholar]
- Viani, R.M.; Gutteberg, T.J.; Lathey, J.L.; Spector, S.A. Lactoferrin inhibits HIV-1 replication in vitro and exhibits synergy when combined with zidovudine. AIDS 1999, 13, 1273–1274. [Google Scholar] [CrossRef]
- Kobayashi, S.; Sato, R.; Aoki, T.; Omoe, K.; Inanami, O.; Hankanga, C.; Yamada, Y.; Tomizawa, N.; Yasuda, J.; Sasaki, J. Effect of bovine lactoferrin on functions of activated feline peripheral blood mononuclear cells during chronic feline immunodeficiency virus infection. J. Vet. Med. Sci. 2008, 70, 429–435. [Google Scholar] [CrossRef]
- Chen, L.T.; Lu, L.; Broxmeyer, H.E. Effects of purified iron-saturated human lactoferrin on spleen morphology in mice infected with Friend virus complex. Am. J. Pathol. 1987, 126, 285–292. [Google Scholar]
- Lu, L.; Shen, R.N.; Zhou, S.Z.; Srivastava, C.; Harrington, M.; Miyazawa, K.; Wu, B.; Lin, Z.H.; Ruscetti, S.; Broxmeyer, H.E. Synergistic effect of human lactoferrin and recombinant murine interferon-gamma on disease progression in mice infected with the polycythemia-inducing strain of the Friend virus complex. Int. J. Hematol. 1991, 54, 117–124. [Google Scholar]
- Yi, M.; Kaneko, S.; Yu, D.Y.; Murakami, S. Hepatitis C virus envelope proteins bind lactoferrin. J. Virol. 1997, 71, 5997–6002. [Google Scholar]
- Ikeda, M.; Sugiyama, K.; Tanaka, T.; Tanaka, K.; Sekihara, H.; Shimotohno, K.; Kato, N. Lactoferrin markedly inhibits hepatitis C virus infection in cultured human hepatocytes. Biochem. Biophys. Res. Commun. 1998, 245, 549–553. [Google Scholar] [CrossRef]
- Ikeda, M.; Nozaki, A.; Sugiyama, K., Tanaka; Naganuma, A.; Tanaka, K.; Sekihara, H.; Shimotohno, K.; Saito, M.; Kato, N. Characterization of antiviral activity of lactoferrin against hepatitis C virus infection in human cultured cells. Virus Res. 2000, 66, 51–63. [Google Scholar] [CrossRef]
- Nozaki, A.; Ikeda, M.; Naganuma, A.; Nakamura, T.; Inudoh, M.; Tanaka, K.; Kato, N. Identification of a lactoferrin-derived peptide possessing binding activity to hepatitis C virus E2 envelope protein. J. Biol. Chem. 2003, 278, 10162–10173. [Google Scholar]
- Abe, K.; Nozaki, A.; Tamura, K.; Ikeda, M.; Naka, K.; Dansako, H.; Hoshino, H.O.; Tanaka, K.; Kato, N. Tandem repeats of lactoferrin-derived anti-hepatitis C virus peptide enhance antiviral activity in cultured human hepatocytes. Microbiol. Immunol. 2007, 51, 117–125. [Google Scholar]
- Beleid, R.; Douglas, D.; Kneteman, N.; Kaur, K. Helical Peptides Derived from Lactoferrin Bind Hepatitis C Virus Envelope Protein E2. Chem. Biol. Drug Des. 2008, 72, 436–443. [Google Scholar] [CrossRef]
- Tanaka, K.; Ikeda, M.; Nozaki, A.; Kato, N.; Tsuda, H.; Saito, S.; Sekihara, H. Lactoferrin inhibits hepatitis C virus viremia in patients with chronic hepatitis C: A pilot study. Jpn J. CancerRes. 1999, 90, 367–371. [Google Scholar] [CrossRef]
- Okada, S.; Tanaka, K.; Sato, T.; Ueno, H.; Saito, S.; Okusaka, T.; Sato, K.; Yamamoto, S.; Kakizoe, T. Dose-response trial of lactoferrin in patients with chronic hepatitis C. Jpn J. Cancer Res. 2002, 93, 1063–1069. [Google Scholar] [CrossRef]
- Ishii, K.; Takamura, N.; Shinohara, M.; Wakui, N.; Shin, H.; Sumino, Y.; Ohmoto, Y.; Teraguchi, S.; Yamauchi, K. Long-term follow-up of chronic hepatitis C patients treated with oral lactoferrin for 12 months. Hepatol. Res. 2003, 25, 226–233. [Google Scholar] [CrossRef]
- Amati, L.; Cozzolongo, R.; Manghisi, O.G.; Cuppone, R.; Pellegrino, N.M.; Caccavo, D.; Birillo, E. The immune responsiveness in hepatitis C virus infected patients: Effects of interferon alfa/ribavirin combined treatment on the lymphocyte response with special reference to B cells. Curr. Pharm. Des. 2004, 36, 2093–2100. [Google Scholar]
- Caradonna, L.; Mastronardi, M.L.; Magrone, T.; Cozzolongo, R.; Cuppone, R.; Manghis, O.G.; Caccavo, D.; Pellegrino, N.M.; Amoroso, A.; Jirillo, E.; Amati, L. Biological and clinical significance of endotoxemia in the course of hepatitis C virus infection. Curr. Pharm. Des. 2002, 8, 995–1005. [Google Scholar] [CrossRef]
- Ueno, H.; Sato, T.; Yamamoto, S.; Tanaka, K.; Ohkawa, S.; Takagi, H.; Yokosuka, O.; Furuse, J.; Saito, H.; Sawaki, A.; et al. Randomized, double-blind, placebo-controlled trial of bovine lactoferrin in patients with chronic hepatitis C. Cancer Sci. 2006, 97, 1105–1110. [Google Scholar] [CrossRef]
- Kaito, M.; Iwasa, M.; Fujita, N.; Kobayashi, Y.; Kojima, Y.; Ikoma, J.; Imoto, I.; Adachi, Y.; Hamano, H.; Yamauchi, K. Effect of lactoferrin in patients with chronic hepatitis C: Combination therapy with interferon and ribavirin. J. Gastroenterol. Hepatol. 2007, 22, 1894–1897. [Google Scholar] [CrossRef]
- el-RM Tabll, A. Camel lactoferrin markedly inhibits hepatitis C virus genotype 4 infection of human peripheral blood leukocytes. J. Immunoassay Immunochem. 2007, 28, 267–277. [Google Scholar] [CrossRef]
- Hara, K.; Ikeda, M.; Saito, S.; Matsumoto, S.; Numata, K.; Kato, N.; Tanaka, K.; Sekihara, H. Lactoferrin inhibits hepatitis B virus infection in cultured human hepatocytes. Res. Hepatol. 2002, 24, 228–235. [Google Scholar] [CrossRef]
- Li, S.; Zhou, H.; Huang, G.; Liu, N. Inhibition of HBV infection by bovine lactoferrin and iron zinc-saturated lactoferrin. Med. Microbiol. Immunol. 2009, 198, 19–25. [Google Scholar] [CrossRef]
- Petrova, M.; Kamburov, V. Breastfeeding and chronic HBV infection: Clinical and social implications. World J. Gastroenterol. 2010, 16, 5042–5046. [Google Scholar] [CrossRef]
- Portelli, J.; Gordon, A.; May, J.T. Effect of compounds with antibacterial activities in human milk on respiratory syncytial virus and citomegalovirus in vitro. J. Med. Microbiol. 1998, 47, 1015–1018. [Google Scholar] [CrossRef]
- Sano, H.; Nagai, K.; Tsutsumi, H.; Kuroki, Y. Lactoferrin and surfactant protein A exhibit distinct binding specificity to F protein and differently modulate respiratory syncytial virus infection. Eur.J. Immunol. 2003, 33, 2894–2902. [Google Scholar]
- Yamamoto, H.; Ura, Y., Tanemura; Koyama, A.; Takano, S.; Uematsu, J.; Kawano, M.; Tsurudome, M.; O’Brien, M.; Komada, H. Inhibitory effect of bovine lactoferrin on human parainfluenza virus type 2 infection. J. Health Sci. 2010, 54, 613–617. [Google Scholar]
- Waarts, B.L.; Aneke, O.J.; Smit, J.M.; Kimata, K.; Bittman, R.; Meijer, D.K.; Wilschut, J. Antiviral activity of human lactoferrin: Inhibition of alphavirus interaction with heparan sulfate. Virology 2005, 333, 284–292. [Google Scholar] [CrossRef]
- Murphy, M.E.; Kariwa, H.; Mizutani, T.; Yoshimatsu, K.; Arikawa, J.; Takashima, I. In vitro antiviral activity of lactoferrin and ribavirin upon hantavirus. Arch. Virol. 2000, 145, 1571–1582. [Google Scholar]
- Hardestam, J.; Petterson, L.; Ahlm, C.; Evander, M.; Lundkvist, A.; Klingström, J. Effect of Human Saliva Against Hantavirus. J. Med. Virol. 2008, 80, 2122–2126. [Google Scholar] [CrossRef]
- Murphy, M.E.; Kariwa, H.; Mizutani, T.; Tanabe, H.; Yoshimatsu, K.; Arikawa, J.; Takashima, I. Characterization of in vitro and in vivo antiviral activity of lactoferrin and ribavirin upon hantavirus. J. Vet. Med. Sci. 2001, 63, 637–645. [Google Scholar] [CrossRef]
- Drobni, P.; Naslund, J.; Evander, M. Lactoferrin inhibits human papilloma virus binding and uptake in vitro. Antiviral Res. 2004, 64, 63–68. [Google Scholar]
- Mistry, N.; Drobni, P.; Naslund, J.; Sunkari, V.G.; Jenssen, H.; Evander, M. The anti papillomavirus activity of human and bovine lactoferricin. Antiviral Res. 2007, 75, 258–265. [Google Scholar] [CrossRef]
- Degener, A.M.; Longhi, G.; Mischitelli, M.; Longhi, C.; Conte, M.P.; Valenti, P.; Seganti, L.; Marchetti, M.; Superti, F. Lactoferrin inhibits human BK polyomavirus adsorption to Vero cells. Biochem. Cell Biol. 2006, 84, 389–389. [Google Scholar]
- Superti, F.; Ammendolia, M.G.; Valenti, P.; Seganti, L. Antirotaviral activity of milk proteins: Lactoferrin prevents rotavirus infection in the enterocyte-like cell line HT-29. Med. Microbiol. Immunol. 1997, 186, 83–91. [Google Scholar] [CrossRef]
- Superti, F.; Siciliano, R.; Rega, B.; Giansanti, F.; Valenti, P.; Antonini, G. Involvement of bovine lactoferrin metal saturation, sialic acid and protein fragments in the inhibition of rotavirus infection. Biochim. Biophys. Acta 2001, 1528, 107–115. [Google Scholar]
- Superti, F.; Donelli, G. Characterization of SA-11 rotavirus receptorial structures on human colon carcinoma cell line HT-29. J. Med. Virol. 1995, 47, 421–428. [Google Scholar] [CrossRef]
- Perez-Cano, F.J.; Marın-Gallen, S.; Castell, M.; Rodrıguez-Palmero, M.; Rivero, M.; Castellote, C.; Franch, A. Supplementing suckling rats with whey protein concentrate modulates the immune response and ameliorates rat rotavirus-induced diarrhea. J. Nutr. 2008, 138, 2392–2398. [Google Scholar] [CrossRef]
- McCann, K.B.; Lee, A.; Wan, J.; Roginski, H.; Coventry, M.J. The effect of bovine lactoferrin and lactoferricin B on the ability of feline calicivirus (a norovirus surrogate) and poliovirus to infect cell cultures. J. Appl. Microbiol. 2003, 95, 1026–1033. [Google Scholar] [CrossRef]
- Pan, Y.; Rowney, M.; Guo, P.; Hobman, P. Biological properties of lactoferrin: An overview. Aust. J. Dairy Technol. 2007, 62, 31–42. [Google Scholar]
- Arnold, D.; Di Biase, A.M.; Marchetti, M.; Pietrantoni, A.; Valenti, P.; Seganti, L.; Superti, F. Antiadenovirus activity of milk proteins: Lactoferrin prevents viral infection. Antiviral Res. 2002, 53, 153–158. [Google Scholar] [CrossRef]
- Marchetti, M.; Superti, F.; Ammendolia, M.G.; Rossi, P.; Valenti, P.; Seganti, L. Inhibition of poliovirus type 1 infection by iron-, manganese- and zinc-saturated lactoferrin. Med. Microbiol.Immunol. 1999, 187, 199–204. [Google Scholar] [CrossRef]
- Di Biase, AM.; Pietrantoni, A.; Tinari, A.; Siciliano, R.; Valenti, P.; Antonimi, G.; Seganti, L.; Superti, F. Heparin-interacting sites of bovine lactoferrin are involved in anti-adenovirus activity. J. Med. Virol. 2003, 69, 495–502. [Google Scholar] [CrossRef]
- Dechecchi, M.C.; Tamanini, A.; Bonizzato, A.; Cabrini, G. Heparan sulfate glycosaminoglycans are involved in adenovirus type 5 and 2-host cell interactions. Virology 2000, 268, 382–390. [Google Scholar] [CrossRef]
- Pietrantoni, A.; Di Biase, A.M.; Tinari, A.; Marchetti, M.; Valenti, P.; Seganti, L.; Superti, F. Bovine lactoferrin inhibits adenovirus infection by interacting with viral structural polypeptides. Antimicrob. Agents Chemother. 2003, 47, 2688–2691. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Roelvink, P.W.; Lizonova, A.; Lee, J.G.; Li, Y.; Bergelson, J.M.; Finberg, R.W.; Brough, D.E.; Kovesdi, I.; Wickham, T.J. The coxsackievirus-adenovirus receptor protein can function as a cellular attachment protein for adenovirus serotypes from subgroups A, C, D, E, and F. J. Virol. 1998, 72, 7909–7915. [Google Scholar]
- Tomko, R.P.; Xu, R.; Philipson, L. HCAR and MCAR: The human and mouse cellular receptors for subgroup C adenoviruses and group B coxsackieviruses. Proc. Natl. Acad. Sci. USA 1997, 94, 3352–3356. [Google Scholar] [CrossRef]
- Roelvink, P.W.; Kovesdi, I.; Wickham, T.J. Comparative analysis of adenovirus fiber-cell interaction: Adenovirus type 2 (Ad2) and Ad9 utilize the same cellular fiber receptor but use different binding strategies for attachment. J. Virol. 1996, 70, 7614–7621. [Google Scholar]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins avb3 and avb5 promote adenovirus internalization but not virus attachment. Cell 1993, 73, 309–319. [Google Scholar] [CrossRef]
- Günther, P.S.; Mikeler, E.; Hamprecht, K.; Schneider-Schaulies, J.; Jahn, Gerhard; Dennehy, K.M. CD209/DC-SIGN mediates efficient infection of monocyte derived dendritic cells by clinical adenovirus 2C isolates in the presence of bovine lactoferrin. J. Gen. Virol. 2011. in Press.. [Google Scholar]
- Ford, E.; Nelson, K.E.; Warren, D. Epidemiology of epidemic keratoconjunctivitis. Epidemiol. Rev. 1987, 9, 244–261. [Google Scholar]
- Kijlstra, A.; Jeurissen, S.H.; Koning, K.M. Lactoferrin levels in normal human tears. Br. J. Ophthalmol. 1983, 67, 199–202. [Google Scholar] [CrossRef]
- Flanagan, J.L. Role of lactoferrin in the tear film. Biochimie 2009, 91, 35–43. [Google Scholar] [CrossRef]
- Johansson, C.; Jonsson, M.; Marttila, M.; Persson, D.; Fan, X.L.; Skog, J.; Frängsmyr, L.; Wadell, G.; Arnberg, N. Adenoviruses use lactoferrin as a bridge for CAR-independent binding to and infection of epithelial cells. J. Virol. 2007, 81, 954–963. [Google Scholar] [CrossRef]
- Adams, W.C.; Bond, E.; Havenga, M.J.; Holterman, L.; Goudsmit, J.; Karlsson Hedestam, G.B.; Koup, R.A.; Loré, K. Adenovirus serotype 5 infects human dendritic cells via a coxsackievirus-adenovirus receptor-independent receptor pathway mediated by lactoferrin and DC-SIGN. J. Gen. Virol. 2009, 90, 1600–1610. [Google Scholar] [CrossRef]
- Sanchez, L.; Ismail, M.; Liew, F.Y.; Brock, J.H. Iron transport across Caco-2 cell monolayers. Effect of transferrin, lactoferrin and nitric oxide. Biochim. Biophys. Acta 1996, 1289, 291–297. [Google Scholar] [CrossRef]
- Rangel, L.M.; Fernandez-Tomas, C.; Dahmus, M.E.; Gariglio, P. Poliovirus-induced modification of host cell RNA polymerase IIO is prevented by cycloheximide and zinc. J. Biol. Chem. 1988, 263, 19267–19269. [Google Scholar]
- Ratka, M.; Lackmann, M.; Ueckermann, C.; Karlins, U.; Koch, G. Poliovirus-associated protein kinase: Destabilization of the virus capsid and stimulation of the phosphorylation reaction by Zn2+. J. Virol. 1989, 63, 3954–3960. [Google Scholar]
- Lin, T.Y.; Chu, C.; Chiu, C.H. Lactoferrin inhibits enterovirus 71 infection of human embryonal rhabdomyosarcoma cells in vitro. J. Infect. Dis. 2002, 186, 1161–1164. [Google Scholar] [CrossRef]
- Chen, H.L.; Wang, L.C.; Chang, C.H.; Yen, C.C.; Cheng, W.T.K.; Wu, S.C.; Hung, C.M.; Kuo, M.F.; Chen, C.M. Recombinant porcine lactoferrin expressed in the milk of transgenic mice protects neonatal mice from a lethal challenge with enterovirus type 71. Vaccine 2008, 26, 891–898. [Google Scholar] [CrossRef]
- Tinari, A.; Pietrantoni, A.; Ammendolia, M.G.; Valenti, P.; Superti, F. Inhibitory activity of bovine lactoferrin against echovirus induced programmed cell death in vitro. Int. J. Antimicrob.Agents 2005, 25, 433–438. [Google Scholar] [CrossRef]
- Ammendolia, M.G.; Pietrantoni, A.; Tinari, A.; Valenti, P.; Superti, F. Bovine lactoferrin inhibits echovirus endocytic pathway by interacting with viral structural polypeptides. Antiviral Res. 2007, 73, 151–160. [Google Scholar] [CrossRef]
- Clarke, N.M.; May, J.T. Effect of antimicrobial factors in human milk on rhinoviruses and milk borne cytomegalovirus in vitro. J. Med. Microbiol. 2000, 49, 719–723. [Google Scholar]
- Pietrantoni, A.; Dofrelli, E.; Tinari, A.; Ammendolia, M.G.; Puzelli, S.; Fabiani, C.; Donatelli, I.; Superti, F. Bovine lactoferrin inhibits Influenza A virus induced programmed cell death in vitro. Biometals 2010, 23, 465–475. [Google Scholar] [CrossRef]
- Taha, S.H.; Mehrez, M.A.; Sitohy, M.Z.; Abou Dawood, A.G.; Abd-El Hamid, M.M.; Kilany, W.H. Effectiveness of esterified whey proteins fractions against Egyptian Lethal Avian Influenza A (H5N1). J. Virol. 2010, 7, 330–333. [Google Scholar] [CrossRef]
- Chien, Y.J.; Chen, W.J.; Hsu, W.L.; Chiou, S.S. Bovine lactoferrin inhibits Japanese encephalitis virus by binding to heparan sulfate and receptor for low density lipoprotein. Virology 2008, 379, 143–151. [Google Scholar] [CrossRef]
- Abdelbacki, A.M.; Taha, S.H.; Sitohy, M.Z.; Abou Dawood, A.I.; Abd-El Hamid, M.M.; Rezk, A.A. Inhibition of tomato yellow leaf curl virus (TYLCV) using whey proteins. Virol. J. 2010, 3, 7–26. [Google Scholar]
- Osterlund, P.J.; Pirhonen, N.; Ikonen, E.; Ronkko, M.; Strengell, S.M.; Makela, M.; Broman, O.J.; Hamming, R.; Hartmann, T.; Ziegler, T.; Julkunen, I. Pandemic H1N1 2009 influenza A virus induces weak cytokine responses in human macrophages and dendritic cells and is highly sensitive to the antiviral actions of interferons. J. Virol. 2010, 84, 1414–1422. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Berlutti, F.; Pantanella, F.; Natalizi, T.; Frioni, A.; Paesano, R.; Polimeni, A.; Valenti, P. Antiviral Properties of Lactoferrin—A Natural Immunity Molecule. Molecules 2011, 16, 6992-7018. https://doi.org/10.3390/molecules16086992
Berlutti F, Pantanella F, Natalizi T, Frioni A, Paesano R, Polimeni A, Valenti P. Antiviral Properties of Lactoferrin—A Natural Immunity Molecule. Molecules. 2011; 16(8):6992-7018. https://doi.org/10.3390/molecules16086992
Chicago/Turabian StyleBerlutti, Francesca, Fabrizio Pantanella, Tiziana Natalizi, Alessandra Frioni, Rosalba Paesano, Antonella Polimeni, and Piera Valenti. 2011. "Antiviral Properties of Lactoferrin—A Natural Immunity Molecule" Molecules 16, no. 8: 6992-7018. https://doi.org/10.3390/molecules16086992
APA StyleBerlutti, F., Pantanella, F., Natalizi, T., Frioni, A., Paesano, R., Polimeni, A., & Valenti, P. (2011). Antiviral Properties of Lactoferrin—A Natural Immunity Molecule. Molecules, 16(8), 6992-7018. https://doi.org/10.3390/molecules16086992